孤独症:大脑极端男性化的表现形态?*
2015-01-23曹漱芹曹颜颜
曹漱芹 曹颜颜
(浙江师范大学杭州幼儿师范学院, 杭州 310012)
孤独症是一种广泛性发育障碍, 主要表现为社会交往和沟通障碍、兴趣狭隘和重复、刻板行为等心理功能缺陷, 同时又呈现出令人费解的孤岛能力和超常的细节加工能力。孤独症的发病率表现出显著的性别差异, 并有逐年增高的趋势。自上世纪80年代以来, 心理学家已提出多种理论来描述和解释孤独症的临床特征, 尤以心理理论假设、弱的中央统合假设和执行功能障碍假设为典型代表。然而, 迄今为止, 学界对孤独症的发生机制还缺乏足够的了解。2002年, 英国剑桥大学的Baron-Cohen带领的研究团队在多年研究的基础上提出了广受关注的“极端男性脑理论”, 力求对患者的症状作出最综合、全面的解释。该理论的提出和有力的解释效力为人类认识孤独症现象提供了一种全新的视角。
1 孤独症极端男性脑理论的内涵及发展历程
1.1 普通人群的共情-系统化理论(Empathizing-Systemizing Theory of General Population)
Baron-Cohen在1997年首先区分了男女心理上的性别差异。他认为, 女性擅长语言、社会判断、共情与合作以及童年假装游戏等; 而男性擅长数学推理、嵌入式数据任务、心理旋转和空间技能等(Baron-Cohen & Hammer, 1997)。即, 女性大脑更倾向于共情(E), 男性大脑则更倾向于系统化(S)。在此, 共情是识别他人情感和想法(认知成分)并对其作出恰当情绪反应(情感成分)的驱力,个体可以通过共情预测他人的行为、共享他人的感受; 系统化则是一个归纳的过程, 是一种基于“如果-就(if-then)”的规则理解和建构系统的驱力(Baron-Cohen, 2002), 6种不同的系统分别是:(1)技术系统, 如交通工具、机械等; (2)自然系统, 如生态学、生物学等; (3)抽象系统, 如逻辑、数学等;(4)社会系统, 如法律、经济等; (5)可组织的系统,如分类学; (6)运动系统, 如玩乐器或扔飞镖等物理运动(Baron-Cohen, 2010)。个体可以通过观察系统中物体或行为的规律, 分析“输入-操作-输出”关系来推断控制这些系统的规则, 以达到理解该系统的目的(Baron-Cohen & Belmonte, 2005)。
在进行上述定义之后, Baron-Cohen接着又分析了共情和系统化技能在男性和女性中的分布。他认为:(1)共情和系统化技能在一般男性和女性人群中都呈正态分布; (2)男性的系统化水平普遍高于女性; (3)女性的共情水平普遍高于男性; (4)共情和系统化技能的性别分布大部分是重叠的。最后一条意味着, 虽然男性普遍更擅长系统化,女性更擅长共情, 但也有许多男性拥有较好的共情能力, 同时也有不少女性的系统化水平较高(Baron-Cohen, 2004)。
1.2 孤独症极端男性脑理论(Extreme Male Brain Theory of Autism)
极端男性脑(EMB)理论是Baron-Cohen等基于大量有关孤独症的心理理论和社会脑的研究成果而提出的, 也是对共情-系统化理论的扩展。Baron-Cohen指出, 男性的共情水平低于女性, 而孤独症个体的共情水平低于男性; 男性系统化水平高于女性, 而孤独症个体的系统化水平高于男性, 所以孤独症个体具有一种极端男性化的认知风格(Baron-Cohen, 2004)。具体来说, 孤独症患者的共情障碍主要体现在社会交往、语言沟通和对他人心理状态的理解三方面; 系统化超常也包括三方面, 分别是孤岛能力、对系统的痴迷以及重复行为(Baron-Cohen, 2004)。
为了更加清晰地描述这种特征, Baron-Cohen(2010)用共情和系统化两个维度划分出了5种“脑类型” (E代表共情, S代表系统化), 具体理论模型见图1。大多数孤独症个体(约65%)表现为极端S类型, 这类个体系统化强于一般水平, 但共情时就会遇到困难(Goldenfeld, Baron-Cohen, & Wheelwright, 2005)。Baron-Cohen (2004)还认为大脑类型可以遗传, 从而解释了孤独症高的遗传性。研究发现, 孤独症患者的家庭中经常包含物理学家、工程师、数学家等, 所有这些人似乎都拥有高度系统化的技能。

图1 EMB理论模型
1.3 孤独症高度系统化理论(Hyper-Systemizing Theory of Autism)
在提出EMB理论后的几年中, Baron-Cohen搜集了大量关于孤独症个体共情障碍和系统化无损甚至超常的证据并对其进行了扩展, 于2006年提出了高度系统化理论。该理论认为孤独症患者具有高度的系统化水平, 细节加工是系统化的前提, 而感觉超敏则是细节加工的基础(Baron-Cohen,Ashwin, Ashwin, Tavassoli, & Chakrabarti, 2009)。高度系统化理论将系统化水平分为1~8级, 个体的系统化兴趣或驱力随等级的升高而逐渐增强。健康人群的系统化水平分布在1~4级, 孤独症患者为5~8级, 因此孤独症患者拥有异常强烈的系统化驱力。该理论指出每一个体都有一个类似于音量调节器的系统化机制(SM), 任何个体的SM水平都取决于该个体的生物学基础, 即不同个体中SM设置的水平不同。男性的SM设置在一个比女性稍高的水平上, 孤独症患者的SM设置在一个比正常男性更高的水平上, 而孤独症儿童父母的SM则处于阿斯伯格综合症与正常男性之间的水平(Baron-Cohen, 2006)。如图2所示。
高度系统化理论认为, 孤独症个体需要借助“输入-操作-输出”的关系来理解系统。这种理解往往通过改变系统内一个变量、控制其他变量,并对这种操作下系统中的所有细节特征的变化进行反复观察而达到目标。因此, 孤独症患者的重复行为可视为个体为理解物体运动背后的规律(即理解系统)所做的努力。在Baron-Cohen看来, 孤独症患者通过重复行为产生相同的结果, 可从可预测的世界中得到一些情感上的快乐。他们的刻板、重复行为、对枯燥系统的着迷、阻抗变化、同一性需求以及孤岛能力都是高度系统化的表现(Baron-Cohen, 2008)。
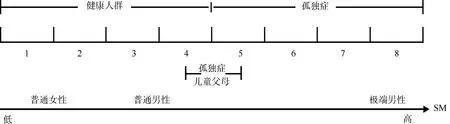
图2 系统化水平等级及不同人群的分布
总之, EMB理论不再把普通人群与孤独症患者视为两个异质的群体, 而是将其纳入普通人群的范畴来解释其障碍特质; 指出孤独症的出现是大脑雄性化的体现, 从而融合了病理学和心理学的解释理论(徐光兴, 2007)。EMB理论提出后,Baron-Cohen等团队展开了一系列的研究, 不断对其进行完善。众多学者也加入其中, 并纷纷对其展开验证, 由此在多个领域迅速积累了大量研究成果。
2 心理学研究:共情障碍和系统化超常
Baron-Cohen及其团队研制了共情商测验(Empathy Quotient, EQ)和系统化商测验(Systemizing Quotient, SQ)用于测量个体的共情和系统化能力。Auyeung等人(2009)采用父母报告法对256名4~11岁的孤独症儿童和1256名控制组进行测查,结果显示, 健康男孩的共情分低于女孩, 而孤独症儿童的共情分比健康男孩还要低, 系统化的得分情况则相反; 同时, 98.2%的孤独症儿童为S或极端S的脑类型, 表现出高度男性化的倾向。2014年英国剑桥大学孤独症研究中心的研究小组对811名成年孤独症患者及3906名控制组进行在线测试, 所有被试都完成三项问卷:EQ、SQ修订版(SQ-R)及自闭特质问卷(AQ)。结果显示, 被试在EQ上的得分:健康女性>健康男性>孤独症患者;SQ-R和AQ的得分情况则恰好相反。研究人员还对被试的“脑类型”进行了分析:健康男女最常见的脑类型分别为S型和E型, 而孤独症患者则为S型或极端S型。研究结果说明, 男女孤独症患者都表现出了极端典型的男性特质(Baron-Cohen, Cassidy,Auyeung, Allison, Achoukhi, Robertson, … La, 2014)。这再次为EMB理论提供了强有力的心理学证据。
除了系统化商和共情商的测查, 孤独症群体“低共情-高系统化”的倾向在具体的心理测验任务上也得以验证。表1中, 时间跨度长达7年之久的6项研究共涉及158名阿斯伯格综合症或高功能孤独症患者, 智商均处于正常范围内, 其中4~14岁的儿童38名, 14~63.9岁的成人120名。这些研究表明:在目光接触、言语发展、友谊问卷等一系列与共情相关的任务中, 正常男性的得分低于女性, 而孤独症谱系患者的平均得分比正常男性还要低; 在镶嵌图形、视觉搜索、直觉物理等一系列与系统化相关的任务中, 正常男性的得分显著高于女性, 而孤独症谱系患者的平均得分比正常男性还要高。这些研究为孤独症群体的共情障碍和系统化超常提供了直接的心理学证据,但这些研究并未直接对孤独症群体内部的性别差异作出具体探讨。

表1 支持孤独症EMB理论的实验心理学研究
3 神经科学研究:大脑结构和功能研究
已有研究表明, 人类大脑结构和功能存在性别差异, 而孤独症患者的某些大脑结构和功能可能也呈现出极端男性化的状态, 这构成其心理和行为表现的神经基础。
3.1 脑体积研究
Giedd等人(1996)运用核磁共振成像技术对99名4~18岁的正常儿童和青少年大脑的定量研究表明:男性大脑普遍比女性大, 在整个大脑体积上男性比女性大约9%, 且这种差异主要是脑白质的作用。在孤独症研究中, 研究者同样发现,孤独症儿童的脑体积更大。早在1943年Kanner观察11名病例时就发现, 其中5名患者的头围尺寸过大。近年来, 人们运用脑成像技术和尸体解剖等方法针对孤独症个体的脑结构展开了大量研究, 最为一致的发现是孤独症个体生命早期大脑的过度生长现象(Courchesne, Campbell, & Solso,2011)。与健康对照组相比, 在出生后的第一年到第二年之间, 孤独症儿童的大脑体积出现过度生长的状况(大约25%~30%), 而后增长速度下降或趋于稳定(Parellada et al., 2014), 脑体积逐渐接近正常水平。而进一步的检测发现, 孤独症儿童脑体积过大的原因主要来自于脑白质的过度生长。但这种过度生长具有区域的选择性, 一般表现为局部脑区内部的脑白质的过度生长, 而在长距离的脑区之间的脑白质连接则反而显得体积不足(曹漱芹, 方俊明, 2008)。
研究表明, 正常男性某些脑区的体积大于正常女性, 主要包括杏仁核、颞上回和小脑等, 而孤独症群体这些区域的体积大于正常人群; 而在前额皮层、扣带回、胼胝体、侧额-顶叶皮质和外侧裂言语区等区域, 正常男性的体积小于正常女性,而孤独症群体在这些区域的体积也小于正常群体,具体的研究参见表2。根据EMB理论的预测, 孤独症群体某些大脑区域或结构可能同样表现出“孤独症谱系>正常男性>正常女性”的倾向, 而某些脑区或结构的情况则相反(Baron-Cohen et al.,2011)。但是, 这一假设目前并未见有研究进行直接验证。
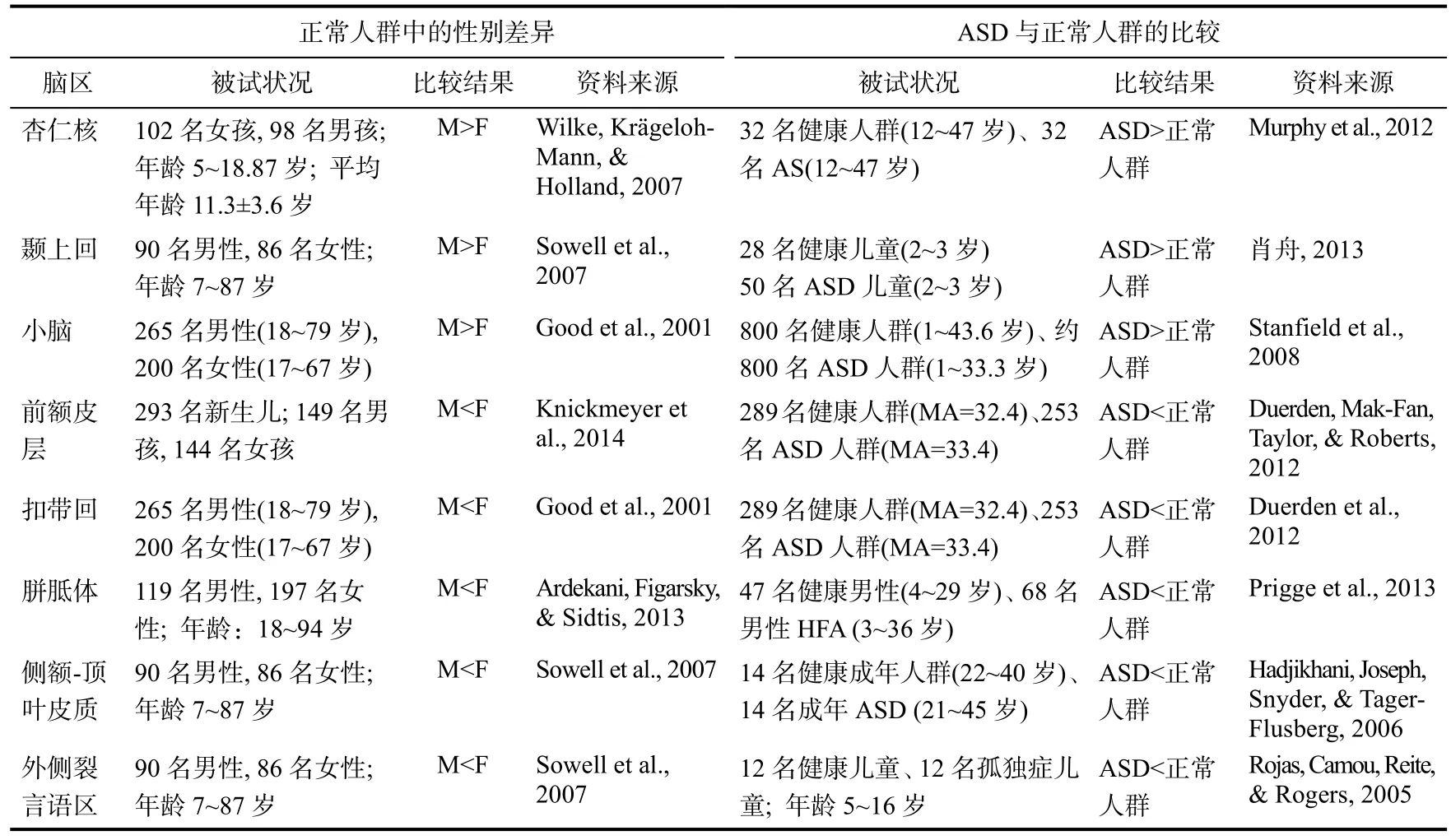
表2 普通人群脑体积的性别差异及孤独症谱系群体与正常人群的比较
3.2 脑功能研究
有关孤独症极端男性脑神经功能的研究于近几年才开展, 主要研究范式是在被试完成共情或系统化任务的同时对其脑区活动展开脑成像研究。
Baron-Cohen认为共情和系统化的神经基础分别是“社会脑”和“分析脑” (Baron-Cohen &Belmonte, 2005)。“社会脑”模型最初由Brothers(1990)提出, 该模型包括三个区域:杏仁核、眶额皮层和内侧额叶以及部分颞叶(颞上沟和梭状回),而大量研究显示孤独症患者在这些区域均存在异常。几项功能性磁共振成像(fMRI)研究显示, 在共情任务, 包括RMET、识别心理状态词语、心理理论、语音加工及面孔加工中, 成年HFA和AS患者在相应脑区, 包括杏仁核(Amygdala)、眶额皮层(Orbitofrontal cortex)、内侧额叶左侧(Left medial frontal cortex)、颞上沟(Superior Temporal Sulcus)及梭状回面孔区(Fusiform Face Area)的激活均显著低于对照组(Baron-Cohen & Belmonte,2005; Tager-Flusberg, 2008)。
Hall等人(2012)对孤独症及控制组的社会脑功能进行的一项fMRI研究发现:在根据面孔进行社会判断尤其是有关情感性质的判断时, 男女的社会脑功能表现出显著差异, 差异最大的脑区为额下皮层(Inferior frontal cortex); 这种差异在ASD个体中被扩大, 显示出极端男性类型。默认网络(DMN)的功能连接也出现类似情况。DMN包括前额皮层、楔前叶、后扣带皮层和颞上沟, 涉及自我参照任务(如与自我相关的识别, 包括对自己情绪的了解)和社会识别(包括心理理论和对他人观点的识别) (Takeuchi et al., 2013)。共情与DMN相关, 因此有研究者推测孤独症DMN的功能连接可能存在异常。Assaf等人(2010)采用fMRI技术对16名HFA患者和16名健康控制组进行研究, 发现在静息状态下, 正常人群中男性的DMN相较于女性的功能连接偏低, 而在孤独症患者中,男性的这种功能连接度更低。这两项研究都表明在与共情相关的脑区活动上, 孤独症也有极端男性化的表现。
上述研究揭示了孤独症患者共情障碍的神经基础, 然而迄今为止, 系统化超常的神经基础——“分析脑”的机制尚不明确。目前的研究主要集中于在镶嵌图形测验(EFT)和视觉搜索测验中对孤独症患者与常人的比较。在EFT中, 孤独症患者的腹侧枕叶(Ventral occipital)区域显示出异常高的激活, 而前额叶(Prefrontal)和顶叶(Parietal)区的激活则异常低(Baron-Cohen & Belmonte, 2005)。枕叶主要负责处理视觉信息, 此区域异常高的激活可能与孤独症患者超常的视觉加工能力有关。同时, 孤独症患者非常擅长视觉搜索测验, 他们能快速有效地确定目标刺激是否出现。Keehn, Brenner,Palmer, Lincoln和Müller (2008)对孤独症患者这一优势能力的神经基础进行了探索, 结果发现孤独症患者的额-顶网络(Frontal-parietal Network)的激活增加, 而这一区域与视觉注意相联系。
2006年, Baron-Cohen等人运用fMRI技术对AS儿童的12名父母及控制组在共情任务(RMET)和系统化任务(EFT)中的大脑活动进行了综合探查, 结果发现:在MRET任务中, 左侧颞中回(Left Medial Temporal Gyrus)和左侧背外侧前额皮质(Left dorsolateral Prefrontal Cortex)的激活显示出了“女性>男性>AS的父母”的状况, 已有研究证实这些脑区与心理理论相关; 在EFT任务中, 左侧额下回(Left Inferior Frontal Gyrus)的激活显示出了“AS的父母>男性>女性”的状况。这项研究表明, 孤独症儿童的父母在完成上述两项任务时大脑的活动趋向于高度男性化, 与EMB理论的预测相一致。
上述研究表明, 孤独症群体共情缺陷的神经机制我们已有所了解, 而有关系统化超常的神经机制研究虽然做了一些尝试, 却仍不够明晰, 未来的研究应致力于进一步揭示孤独症“分析脑”的神经基础。
4 生物学研究:产前雄性激素的研究
在心理学和神经科学水平上的一系列发现引出了一个问题:导致上述差异现象出现的生物学基础是什么?研究发现, 产前雄性激素(包括胎儿期睾丸分泌的睾丸素)是形成性别差异的关键生物学机制。胎儿在母体中接触到的睾丸素水平越高, 出生后的雄性化行为就越明显。睾丸素是非常小的亲脂性分子, 能够轻易穿过血脑屏障和细胞膜, 从而作用于大脑使神经结构和功能产生性别差异(Baron-Cohen, Knickmeyer, & Belmonte,2005)。剑桥大学从2002年开始持续追踪, 对儿童产前睾丸激素(fT)水平和出生后行为发展之间的关系进行了一系列探索。所得研究成果如表3(Knickmeyer & Baron-Cohen, 2006; Auyeung,Lombardo, & Baron-Cohen, 2013)。该表表明:(1)除一项研究无差异外, 在目光接触、言语发展、共情等社会性领域中, 女性的表现都优于男性,而在狭窄兴趣、系统化及自闭特质等非社会性领域中, 男性的表现则优于女性; (2)fT水平与个体在社会性领域的表现呈负相关, 与非社会性领域的表现呈正相关。这些都说明fT对性别差异的形成有重要影响。
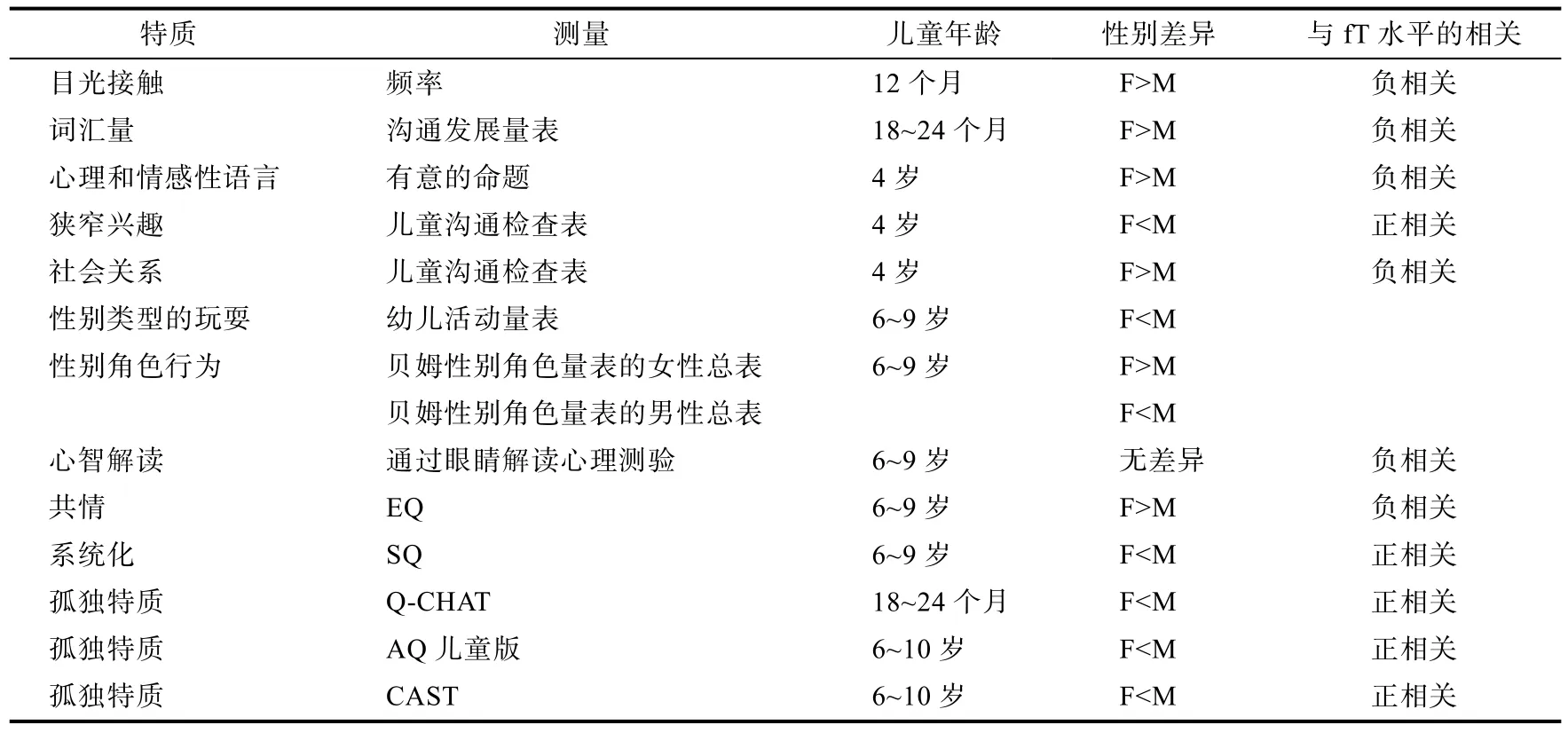
表3 剑桥大学产前睾丸素(fT)项目
在孤独症儿童的研究上, Baron-Cohen认为,孤独症的一个主要诱发因素可能正是胎儿期暴露于高水平的宫内睾丸素之中(Baron-Cohen, 2002)。为了证明这一假设, Auyeung等(2009)对235名孕妇子宫内的雄性激素水平进行取样分析, 并计算产前雄性激素的含量与儿童8岁时孤独症谱系商数(AQ)之间的相关。结果表明雄性激素水平越高则儿童的自闭指数更高, 表现在社交、想象力、共同注意、共情的发展状况比较差, 但对细节的记忆力强。Auyeung, Taylor, Hackett和Baron-Cohen (2010)又选取了129名(66名男孩, 63名女孩)18到24个月大的儿童作为被试, 分析其羊水中的雄性激素水平, 并对儿童的孤独症特质进行测定, 同样发现孤独症特质与雄性激素水平呈正相关。2014年, Baron-Cohen等人第一次对128名出生于1993~1999年的孤独症患者羊水样本中性甾体(包括睾丸素)的活性进行分析, 发现与对照组相比患者的睾丸素活性升高。这些结果表明, 胎儿期高的雄性激素, 会降低儿童的共情能力, 导致行为和能力类型的雄性化, 使得儿童心理朝向系统化的方向发展, 严重时会呈现出“极端男性脑”特质(Chapman et al., 2006)。这几个重要的追踪研究, 为EMB理论提供了最为重要的生物学证据。
5 其他研究
5.1 性别比研究
众所周知, 孤独症群体中男性更容易患上孤独症, 这也间接支持了EMB理论。1943年,Kanner报告的11个案例中有8个是男孩; 1944年Asperger报告的4个案例全是男孩。Asperger曾写道:“值得注意的是, 我们所看到的孤独症儿童几乎全是男孩” (引自Rivet & Matson, 2011)。研究发现, 随智商水平的逐渐上升, 孤独症儿童发病率的男女性别比例呈逐步增大的趋势。在低功能孤独症(IQ<55)中, 男女儿童发病率的性别比例为2.5:1~3:1左右; 而在AS中, 此种障碍发病率的男女性别比例在6:1~16:1左右(关文军, 邓猛,2013)。美国疾病防治中心2012年的数据显示, 每88名儿童中就有1名是孤独症患者, 而每50名男孩中就有1名男孩是孤独症(CDC, 2012)。这些数据都间接地揭示了男性特质与孤独症之间的潜在关联。
5.2 指长比研究
指长比是又一有趣的间接证据。指长比(digit ratios)是指各指(趾)长度(digit, D)的比值, 常用食指指长(2D)和环指指长(4D)的比值(2D:4D)来表示。2D:4D是公认的产前雄性激素活动的生物学标志, 母体子宫内较高的雄性激素水平将导致低2D:4D个体的产生(Valla & Halazonetis, 2014)。指长比具有显著的性别差异, 通常男性的环指长于食指(4D>2D), 女性的食指基本与环指等长或稍长(2D≥4D)。女性2D:4D的平均值为1.00, 高于男性的平均值0.96 (引自曹漱芹, 曹颜颜, 2014)。Hauth, Bruijn, Staal, Buitelaar和Rommelse (2014)针对216名孤独症障碍儿童及202名控制组儿童的研究发现, 孤独症组2D:4D显著高于控制组。然而, 也有质疑性的证据。例如, Falter, Plaisted和Davis (2008)的研究发现孤独症儿童的2D:4D与阿斯伯格综合症儿童及正常儿童并无显著差异。Bejerot等人(2012)针对50名20至47岁的孤独症成人的研究发现男性孤独症患者2D:4D更高而女性患者的2D:4D更低。以上差异的存在, 可能是由于被试选择、测量方法的差异等导致。但总体而言, 现有的大部分研究支持自闭症群体指长比更低的结论。Hönekopp (2012)在一项元分析中指出, 孤独症患者的2D:4D总体上显著低于健康控制组, 表明孤独症群体的指长比具有雄性化倾向, 具有极端男性的特质。但这一结论还需要进一步的研究验证。
5.3 利手研究
利手一般反映的是个体的大脑单侧优势。Geschwind的性激素理论指出利手一侧化的原因决定于胎内睾丸素的水平, 宫内过高的睾丸素会抑制左脑的发育, 使大脑的发育能量移向右半球(引自Treffert, 2011)。其中左半球主要负责语言功能, 称为标准优势, 正常人群中, 70%的人都是右利; 而左利手和混利手人群的语言功能可能侧化于左或右半球的任何一侧, 称为变侧优势(静进,2006)。研究证实, 在孤独症群体中存在更多左利手和混利手的情况。我国暨南大学对20 名孤独症幼儿利手状况的一项调查显示, 孤独症混合利发生率为65%, 而正常儿童混合利率仅为20%,两组差异十分显著(陈卓铭, 李冰肖, 尹义臣, 杜志宏, 2005)。最近, Forrester, Pegler, Thomas和Mareschal (2014)对4~5岁的孤独症儿童及控制组的超过1200种单手操作活动进行研究, 也发现孤独症儿童的混合利高于正常儿童。Lindell和Hudry (2013)对二十余项利手情况的调查研究进行了回顾, 这些研究均证实孤独症群体左利和混合利的发生率高于控制组。然而, 也同样存在不同的研究结论。Preslar, Kushner, Marino和Pearce(2014)进行的一项元分析表明, 孤独症群体的利手情况与正常人群并无显著差异。可见, 孤独症群体的利手状况还需要更大的研究样本和更加细致的研究范式来加以确认。如若利手的形成与孤独症的发病有着相似的生物学机制, 那么我们可以预测利手与EMB也存在着某种相关, 这需要未来研究的进一步验证。
5.4 玩耍行为研究
男孩与女孩在玩耍中显示出的性别差异也为共情和系统化的早期差异提供了证据。在玩具选择上, 1岁大的男孩就倾向于独立选择与系统化相关的玩具, 如:交通工具和建构积木等; 而女孩则更喜欢洋娃娃。在画画时, 男孩也喜欢画工具和武器(Baron-Cohen, 2002)。新生儿的研究显示, 一天大的男婴注视机械运动的时间比注视人面孔时间更长, 而女婴则刚好相反(Baron-Cohen, 2008)。在孤独症儿童的玩耍行为方面, 研究结论比较一致。Meyer (2008)对孤独症女孩的玩具选择进行了研究, 结果发现, 孤独症女孩倾向于选择如小汽车或可操作性的玩具, 并且在选择这类玩具后参与玩耍的时间也比其他玩具要长。Knickmeyer,Wheelwright和Baron-Cohen (2008)也对66名孤独症儿童的玩耍偏好进行了研究, 同样发现孤独症男孩表现出与他们性别相一致的偏好, 孤独症女孩则表现出男性类型的玩耍偏好。这些结果与EMB理论的预测一致。
5.5 父母职业研究
研究一致发现, 孤独症儿童的父亲和祖父从事工程类职业的可能性比一般男性要高(Roelfsema et al., 2012)。而孤独症谱系障碍在物理科学和工程科学家庭中的发病率比在文学专业的家庭高出六倍, 研究者认为这些职业都是在高度专业化的技术领域, 不需要高水平的社会交往能力(Wen &Wen, 2014), 从事这类职业的个体大脑类型倾向于S型。EMB理论认为大脑的类型是可以遗传的,因此孤独症在物理学家、工程师和数学家的家庭中发生率较高(Kanazawa & Vandermassen, 2005)。对此, 高度系统化理论做了进一步说明:孤独症儿童是其父母双方两个高度系统化者的选型交配的结果。患者父母的SM都设置在平均水平之上,结果导致他们孩子的SM设置在一个极端水平,即极端男性脑的出现(Baron-Cohen, 2006)。
5.6 孤岛能力研究
孤岛能力, 是指个体在一般能力普遍落后的情况下某一个或几个领域表现出来的与其整体能力极不匹配的超常能力(曹漱芹, 2013)。孤岛能力虽然少见, 但在医学与心理学领域已有超过一个世纪的研究历史。长时间的观察发现, 相较于其他障碍, 孤岛能力在孤独症群体中更为普遍。Rimland采用父母报告法对5400名孤独症儿童进行调查, 发现其中的531 (9.8%)名儿童具有孤岛能力, 而其在智力障碍中的发病率仅为0.06% (引自Wallace, 2008)。1995年, Young走访了许多国家和地区的51名“天才”, 并进行了标准心理测试,其中41人被确诊为孤独症患者(引自Treffert,2011)。孤独症患者中男性显著多于女性, 而具有领域孤岛能力的孤独症者也以男性居多, 比例约为6:1 (Corrigana, Richardsa, Treffert, & Dager,2012)。在仔细研究后发现, 孤岛能力大多具有右脑倾向, 且仅表现在狭窄的领域中, 主要涉及日历计算、音乐、艺术、数学和机械技艺等(Durukan& Türkbay, 2010)。高度系统化理论认为具有孤岛能力的孤独症患者都有一个共同点, 就是这些个体都是识别刺激中重复模式(即系统化)的专家,而这些孤岛天才所擅长的领域也都是高度可系统化的领域。探寻孤独症和天才之间的联系, 应该从感觉超敏开始, 正是这种感觉超敏引起了卓越的细节加工能力, 而这恰恰是高度系统化的先决条件(Baron-Cohen et al., 2009)。这为未来天才症候群的研究提供了新的思路。
6 质疑性的观点
尽管EMB理论得到各种层面研究汇聚式的验证和支持, 但仍存在一些尚未解决的问题和不足, 也因此受到质疑。
6.1 普通人群中研究结论不一致
近年来, 大量研究对于普通人群共情-系统化理论进行了验证。按照预测, 普通个体的系统化商(SQ)应与其孤独特质指数(AQ)存在正相关, 而与其共情商(EQ)存在负相关。然而, 这一预测在普通群体中有不同的发现。例如, Valla等人(2010)的研究发现, 65名普通男性(平均年龄20.2岁)的EQ与SQ分数呈负相关, 社会性任务(RMET)的得分与非社会性任务(EFT)的得分也存在负相关,而在79名普通女性(平均年龄20.2岁)身上这种负相关均不存在; Morsanyi, Primi, Handley, Chiesi和Galli (2012)开展了两项实验检验EMB理论的核心主张。结果显示, 225名被试(男性62名, 女性163名; 年龄17~46岁)的AQ、SQ和系统化技能之间不存在相关。这些研究均表明, 共情-系统化的假设在普通人群中的适用性还需进一步更大样本、更全面的验证。
此外, 尽管Baron-Cohen等通过大量的研究总体上证实了早期宫内睾丸素水平与儿童期EQ和SQ等的相关, 但这一发现在不同性别幼儿中也有不一致的结论。例如, Chakrabarti, Belmonte和Baron-Cohen (2006)年的研究显示, 男性胎儿宫内的睾丸素水平(fT)与幼儿时期的EQ呈负相关, 与SQ呈正相关; 女性胎儿宫内的睾丸素水平与其幼儿时的EQ也呈负相关, 但与SQ则不相关。在脑体积的研究方面, 也有质疑性的证据。Inano等人(2013)对861名正常被试(年龄24.0~84.8岁, 平均年龄56.1±9.8岁)的神经解剖体积进行检测, 结果发现大脑白质、小脑、和杏仁核的体积并不存在性别差异, 这一发现与EMB的预测也并不一致。这些研究都对于EMB理论的生理基础提出了质疑。
6.2 孤独症群体研究结论的不一致
尽管EMB理论的假设在孤独症群体中得到了多个层面的汇聚式支持, 但同样也有不一致的发现。近期的好几项研究揭示, 女性孤独症确实具有男性化的特质, 而部分男性孤独症可能反而具有女性化的特质。例如, Bejerot等人(2012)的研究发现, 与普通女性相比, 女性ASD具有更高的睾丸素水平、更少的女性面部特征以及更大的头围, 也就是表现出极端男性化的特质; 然而, 相较于普通男性, 男性ASD并没有表现出极端男性的特质, 而是表现出更多女性化的特质, 具有更少的男性身体特征以及更高的2D:4D等。与此相一致, Lai等(2013)利用基于像素的形态学测量法分析了孤独症男女在大脑水平上的神经解剖差异,结果也显示女性患者具有“男性化”的大脑, 而男性患者则没有。因此, 研究者推测:虽然男女患者是由同一诊断标准定义, 但是他们可能涉及不同的生物基础, 分属于不同的亚群。这些研究表明, 孤独症群体内部在心理和大脑水平上存在性别差异, 可能不能用单一的“极端男性脑”理论对整个群体加以统一解释。EMB理论要最终获得全面认可, 还有待于对孤独症群体的性别差异作出明确的解释。
6.3 基因研究的不足
综观文献, 有关EMB理论的基因研究逐渐成为当前的研究热点, 但总体上仍显数量不足, 目前仅看到两项相关研究。一是Chakrabarti等人(2009)年开展的研究。该研究将68个候选基因分成三组:性甾体基因、社交反应基因和神经发育基因, 并分析这三组基因与孤独症特质、共情和AS的相关。结果显示, 有19个基因与AQ或EQ具有显著相关, 它们分别为性甾体组的ESR2和CYP11B1, 社交反应组的MAOB、AVPR17A1和CYP19A1, 以及神经发育组的HOXA1、NTRK1和NLGN4X。另一项是Warrier, Baron-Cohen和Chakrabarti (2013)开展的研究。他们对GABRB3中45个基因型的单核基因多态性(SNPs)与AS及其相关特质的相关进行检测。结果显示, 两个SNPs(rs9806546和rs11636966)与EQ显著相关; 两个SNP-SNP对(rs12438141-rs1035751和rs12438141-rs7179514)与EFT的分数显著相关; 一个SNP-SNP对(rs7174437-rs1863455)与心理旋转的分数显著相关。这两项研究为揭示EMB的相关基因奠定了基础, 但具体的致病基因还需进一步的精确定位;同时还需要扩大候选基因的范围以确定更多相关基因。
6.4 跨文化研究的不足
社会文化对孤独症群体的影响, 表现在两方面:一是对孤独症群体的症状表达与解释产生影响, 体现在社会文化对性别角色的塑造和基于性别角色期望对不同性别个体行为差异的解释与对待方面; 另一方面是对孤独症儿童的诊断产生影响, 体现在社会文化等因素对孤独症儿童的社会知觉、症状解释及诊断识别产生影响(Kreiser &White, 2014)。一项涉及多个国家的跨文化研究对145名来自英国、美国、以色列及韩国的孤独症儿童进行了对比, 发现这四个国家的孤独症儿童在沟通交流损伤和刻板行为问题上存在极大的跨文化差异(Matson et al., 2011)。因此, 不同的文化也可能会对个体的EQ、SQ得分以及脑类型的分布产生影响。例如, 在大多数国家, 女性的系统化商一般低于男性, 但在以色列等国家, 女性由于常常接受多种技术训练而在数学、空间能力测验上优于男性(Shaffer & Kipp, 2002/2013), 也就是说可能系统化分数更高。而目前EMB理论的相关研究主要在英国展开, 日本、美国等几个国家近几年才开始涉及, 且非常有限。跨文化研究的不足由此限制了EMB理论的解释效力。
7 研究展望
EMB理论很好的解释了孤独症群体中男性显著多于女性的现象, 以及他们社会、语言沟通等与共情相关的缺陷与孤岛能力并存等临床特征,为我们认识孤独症现象及寻找相应的教育干预途径带来了重要的启示。EMB理论的研究已取得相当丰硕的成果, 但还需更详细标准化的数据, 尤其是神经科学水平的研究。2014年, 剑桥大学孤独症研究中心的研究团队检索了1990年至2013年期间的一百多项有关人类大脑结构性别差异的相关研究, 并第一次对这些研究两性全脑和部分脑体积差异的大脑成像文献进行了元分析。研究发现, 大脑确实存在性别差异:(1)男性的整个大脑体积平均比女性的大8%到13%; (2)男女的脑区体积也存在差异, 这些区域包括处理情感的边缘系统和语言系统等。该研究还创建出了一个大脑性别差异图(见图3), a代表未修正的脑区灰质体积性别差异总图; b~f展示的是脑区性别差异的具体细节(见表4) (Ruigrok et al., 2014)。这项研究提供了一个大脑性别差异的标准, 对于孤独症的研究可能具有重要意义。未来研究可以此为基础, 探究孤独症在这些脑区是否表现出极端男性的特质。
此外, 未来可以从以下几方面开展研究:
在理论建构上, 要开展本土化研究, 验证该理论的跨文化一致性, 以扩大理论的解释范围。不同的文化可能会对个体的EQ、SQ得分以及脑类型的分布产生影响。为了检验EMB理论的跨文化稳定性, Wakabayashi等人(2007)在日本对3组被试(孤独症组、一般人群控制组和大学生控制组)进行测查, 每个被试均完成EQ、SQ问卷。结果显示:3组被试的EQ、SQ得分以及脑类型的分布情况与EMB理论的预测相一致。此项研究说明EMB理论适用于日本文化, 但是否同样适用于中国文化, 还需要进一步验证。未来应该进行本土化的理论建构, 探讨EMB理论在中国孤独症患者中的独特价值。

图3 基于体素的脑区灰质体积性别差异图(红色:女性>男性; 蓝色:男性>女性。彩图见电子版)

表4 脑区灰质体积性别差异(图3中b~f)
在神经机制上, 要丰富研究范式探索系统化的神经机制“分析脑”模型。EMB理论在心理学水平上的研究成果已经非常丰富, 而神经科学研究则是近几年才开展, 且较偏重于对共情的研究。共情的神经机制“社会脑”已经得到相关研究的支持, 而系统化的神经机制“分析脑”则尚未明确。Takeuchi (2013)等人发现系统化与外部注意系统(EAS)的功能相关。EAS包括外侧前额皮层、前扣带皮层背侧和顶下小叶等, 其中外侧前额皮层和顶下小叶与空间任务、注意和推理相关。未来对系统化的研究可以此为基础进行探查, 结合采用事件相关电位(ERPs)、脑磁图(MEG)、fMRI、镜红外成像技术等多种技术手段帮助建立“分析脑”模型。
在被试选取上, 要解决研究对象的性别失衡问题。现有研究对女性孤独症还没有充分认识,我们不应盲目认为有关男性孤独症的发现也适用于女性。未来研究应尽量平衡孤独症被试的性别比例, 避免严重失衡导致研究结果异常。另外,Krahn与Fenton (2012)认为EMB理论带有明显的经验主义色彩, 可能会使孤独症儿童的诊断无意中偏向男孩; 即使该理论是正确的, 也会对孤独症女孩的诊断、早期干预和服务产生相当严重的后果。这也要求今后的研究提高对女性孤独症被试的重视。
在教育应用上, 要根据EMB理论开发教育干预模式。Baron-Cohen (2009)认为, 现有的孤独症干预方法只是简单的塑造行为, 过度依赖于外部强化或奖励, 应注重利用孤独症儿童的优势和内在需要实施干预。他依据EMB理论开发了利用孤独症的系统化驱力改善其情绪识别及社交能力的方法。这些方法的共同特点是高度结构化和关注系统, 因此可以吸引孤独症儿童的注意力。可能没有一种单一的教育方法对孤独症起作用, 但很有可能那些能够吸引孤独症儿童的方法是最有效的。这在伦理上也意味着, 我们应该让环境来适应孤独症儿童, 而不是让他们加入我们的世界。
总之, EMB理论由 Baron-Cohen正式提出已有十余年的时间, 该理论的核心观点在于认为孤独症不是一种疾病, 而是一种认知风格。近年来,该理论在心理学、神经科学、生物学及其他相关领域的研究发展迅速, 研究者对其内涵有了更加深入的了解。但仍有些问题尚待解决, 这些问题需要研究者通过技术手段的完善以及研究范式的创新来进一步深入探讨。
曹漱芹. (2013). 孤独症群体的孤岛能力.心理科学进展,21(8), 1457-1465.
曹漱芹, 曹颜颜. (2014). 指长比与儿童自闭症—兼论自闭症“极端男性脑”理论及启示.中国特殊教育,(8), 67-72.
曹漱芹, 方俊明. (2008). 孤独症生命早期脑过度生长现象及启示.心理科学进展, 16(2), 282-289.
陈卓铭, 李冰肖, 尹义臣, 杜志宏. (2005). 自闭症儿童利手调查分析.广东医学, 26(7), 912-914.
关文军, 邓猛. (2013). 孤独症儿童的性别差异及其解释模型.中国特殊教育,(12), 52-58.
静进. (2006). 儿童左利的生物学意义及其某些神经心理特征.中国儿童保健杂志, 14(2), 113-115.
肖舟. (2013).2-3岁孤独症谱系障碍儿童极端男性化大脑理论的脑影像学研究(硕士学位论文). 南京医科大学.
徐光兴. (2007). 自闭症的性别差异及其与认知神经功能障碍的关系.心理科学, 30(2), 425-428.
Ardekani, B. A., Figarsky, K., & Sidtis, J. J. (2013). Sexual dimorphism in the human corpus callosum: An MRI study using the OASIS brain database.Cerebral Cortex, 23(10),2514-2520.
Assaf, M., Jagannathan, K., Calhoun, V. D., Miller, L.,Stevens, M. C., Sahl, R., … Pearlson, G. D. (2010).Abnormal functional connectivity of default mode subnetworks in autism spectrum disorder patients.NeuroImage,53(1), 247-256.
Auyeung, B., Baron-Cohen, S., Ashwin, E., Knickmeyer, R.,Taylor, K., & Hackett, G. (2009). Fetal testosterone and autistic traits.British Journal of Psychology, 100(1), 1-22.
Auyeung, B., Lombardo, M. V., & Baron-Cohen, S. (2013).Prenatal and postnatal hormone effects on the human brain and cognition.Pflügers Archiv-European Journal of Physiology, 465(5), 557-571.
Auyeung, B., Taylor, K., Hackett, G., & Baron-Cohen, S.(2010). Foetal testosterone and autistic traits in 18 to 24-month-old children.Molecular Autism, 1, 11.
Auyeung, B., Wheelwright, S., Allison, C., Atkinson, M.,Samarawickrema, N., & Baron-Cohen, S. (2009). The children’s empathy quotient and systemizing quotient: Sex differences in typical development and in autism spectrum conditions.Journal of Autism and Development Disorders,39(11), 1509-1521.
Baron-Cohen, S. (2002). The extreme male brain theory of autism.Trends in Cognitive Sciences, 6(6), 248-254.
Baron-Cohen, S. (2004).The essential difference:Male and female brains and the truth about autism. New York: Basic Books.
Baron-Cohen, S. (2006). The hyper-systemizing, assortative mating theory of autism.Progress in Neuro-Psychopharmacology and Biological Psychiatry, 30(5), 865-872.
Baron-Cohen, S. (2008). Autism, hypersystemizing, and truth.Quarterly Journal of Experimental Psychology, 61(1), 64-75.
Baron-Cohen, S. (2009).The empathizing-systemizing theory of autism: Implications for education.Tizard L earning Disability Review, 14(3), 4-13.
Baron-Cohen, S. (2010). Empathizing, systemizing, and the extreme male brain theory of autism.Progress in Brain Research, 186, 167-175.
Baron-Cohen, S., Ashwin, E., Ashwin, C., Tavassoli, T., &Chakrabarti, B. (2009). Talent in autism: Hyper-systemizing,hyper-attention to detail and sensory hypersensitivity.Philosophical T ransactions of the Royal Soc iety B:Biological Sciences, 364(1522), 1377-1383.
Baron-Cohen, S., Auyeung, B., Nørgaard-Pedersen, B.,Hougaard, D. M., Abdallah, M. W., Melgaard, L., …Lombardo, M. V. (2014). Elevated fetal steroidogenic activity in autism.Molecular Psychiatry, 20(3), 369-376.
Baron-Cohen, S., & Belmonte, M. K. (2005). Autism: A window on to the development of the social and the analytic brain.Annual Review of N euroscience, 28,109-126.
Baron-Cohen, S., Cassidy, S., Auyeung, B., Allison, C.,Achoukhi, M., Robertson, S., … La, M.-C. (2014).Attenuation of typical sex differences in 800 adults with autism vs. 3,900 controls.PLoS ONE, 9(7), e102251.
Baron-Cohen, S., & Hammer, J. (1997). Is autism an extreme form of the" male brain"?Advances in Infancy Research,11, 193-218.
Baron-Cohen, S., Knickmeyer, R. C., & Belmonte, M. K.(2005). Sex differences in the brain: Implications for explaining autism.Science, 310(5749), 819-823.
Baron-Cohen, S., Lombardo, M. V., Auyeung, B., Ashwin, E.,Chakrabarti, B., & Knickmeyer, R. (2011). Why are autism spectrum conditions more prevalent in males?PLoS Biology, 9(6), e1001081.
Baron-Cohen, S., Ring, H., Chitnis, X., Wheelwright, S.,Gregory, L., Williams, S., … Bullmore, E. (2006). fMRI of parents of children with Asperger syndrome: A pilot study.Brain and Cognition, 61(1), 122-130.
Baron-Cohen, S., & Wheelwright, S. (2003). The friendship questionnaire (FQ): An investigation of adults with Asperger syndrome or high functioning autism, and normal sex differences.Journal of Autism and Development Disorders,33(5), 509-517.
Baron-Cohen, S., Wheelwright, S., Scahill, V., Lawson, J., &Spong, A. (2001). Are intuitive physics and intuitive psychology independent? A test with children with Asperger syndrome.Journal of Developmental and Learning Disorders,5, 47-78.
Bejerot, S., Eriksson, J. M., Bonde, S., Carlström, K.,Humble, M. B., & Eriksson, E. (2012). The extreme male brain revisited: Gender coherence in adults with autism spectrum disorder.The British Journal of Psychiatry, 201(2),116-123.
Brothers, L. (1990). The social brain: A project for integrating primate behaviour and neurophysiology in a new domain.Concepts in Neuroscience, 1,27-51.
Centers for Disease Control. (2012). Prevalence of autism spectrum disorders-autism and developmental disabilities monitoring network, United States.Morbidity and Mortality Weekly Report Surveillance Summaries, 58, 1-20.
Chakrabarti, B., Belmonte, E., & Baron-Cohen, S. (2006).Empathizing with basic emotions: Common and discrete neural substrates.Social Neuroscience, 1, 364-384.
Chakrabarti, B., Dudbridge, F., Kent, L., Wheelwright, S.,Hill-Cawthorne, G., Allison, C., … Baron-Cohen, S.(2009). Genes related to sex steroids, neural growth, and social-emotional behavior are associated with autistic traits, empathy, and Asperger syndrome.Autism Research,2, 157-177.
Chapman, E., Baron-Cohen, S., Auyeung, B., Knickmeyer,R., Taylor, K., & Hackett, G. (2006). Fetal testosterone and empathy: Evidence from the empathy quotient (EQ)and the “reading the mind in the eyes” test.Social Neuroscience, 1(2), 135-148.
Corrigana, N. M., Richardsa, T. L., Treffert, D. A., & Dager,S. R. (2012). Toward a better understanding of the savant brain.Comprehensive Psychiatry, 53(6), 706-717.
Courchesne, E., Campbell, K., & Solso, S. (2011). Brain growth across the life span in autism: Age-specific changes in anatomical pathology.Brain R esearch, 1380,138-145.
Duerden, E. G., Mak-Fan, K. M., Taylor, M. J., & Roberts, S.W. (2012). Regional differences in grey and white matter in children and adults with autism spectrum disorders: An activation likelihood estimate (ALE) Meta-analysis.Autism Research, 5(1), 49-66.
Durukan, İ., & Türkbay, T. (2010). Savant syndrome: Clinical and neuropsychological features.Current Ap proaches in Psychiatry, 2(2), 237-253.
Falter, C. M., Plaisted, K. C., & Davis, G. (2008).Visuo-spatial processing in autism-testing the predictions of extreme male brain theory.Journal of Developmental and Learning Disorders, 38, 507-515.
Forrester, G. S., Pegler, R., Thomas, M. S. C., & Mareschal, D.(2014). Handedness as a marker of cerebral lateralization in children with and without autism.Behavioural B rain Research, 268, 14-21.
Giedd, J. N., Vaituzis, A. C., Hamburger, S. D., Lange, N.,Rajapakse, J. C., Kaysen, D., … Rapoport, J. L. (1996).Quantitative MRI of the temporal lobe, amygdala, and hippocampus in normal human development: Ages 4-18 years.Journal of Comparative Neurology, 366(2), 223-230.
Goldenfeld, N., Baron-Cohen, S., & Wheelwright, S. (2005).Empathizing and systemizing in males, females and autism.Clinical Neuropsychiatry, 2(6), 338-345.
Good, C. D., Johnsrude, I., Ashburner, J., Henson, R. N. A.,Friston, K. J., & Frackowiak, R. S. J. (2001). Cerebral asymmetry and the effects of sex and handedness on brain structure: A voxel-based morphometric analysis of 465 normal adult human brains.NeuroImage, 14(3), 685-700.
Hadjikhani, N., Joseph, R. M., Snyder, J., & Tager-Flusberg,H. (2006). Anatomical differences in the mirror neuron system and social cognition network in autism.Cerebral Cortex, 16(9), 1276-1282.
Hall, J., Philip, R. C. M., Marwick, K., Whalley, H. C.,Romaniuk, L., McIntosh, A. M., … Lawrie, S. M. (2012).Social cognition, the male brain and the autism spectrum.PLoS One, 7(12), e49033.
Hauth, I., Bruijn, Y. G., Staal, W., Buitelaar, J. K., &Rommelse, N. N. (2014). Testing the extreme male brain theory of autism spectrum disorder in a familial design.Autism Research, 7(4), 491-500.
Hönekopp, J. (2012). Digit ratio 2D:4D in relation to autism spectrum disorders, empathizing, and systemizing: A quantitative review.Autism Research, 5(4), 221-230.
Inano, S., Takao, H., Hayashi, N., Yoshioka, N., Mori, H.,Kunimatsu, A.,... Ohtomo, K. (2013). Effects of age and gender on neuroanatomical volumes.Journal of Magnetic Resonance Imaging, 37(5), 1072-1076.
Jolliffe, T., & Baron-Cohen, S. (1997). Are people with autism and Asperger syndrome faster than normal on the embedded figures task?Journal of Child Psychology and Psychiatry, 38(5), 527-534.
Kanazawa, S., & Vandermassen, G. (2005). Engineers have more sons, nurses have more daughters: An evolutionary psychological extension of Baron-Cohen's extreme male brain theory of autism.Journal of Theor etical B iology,233(4), 589-599.
Kanner, L. (1943). Autistic disturbances of affective contact.Nervous Child, 2(3), 217-250.
Keehn, B., Brenner, L., Palmer, E., Lincoln, A. J., & Müller,R. A. (2008). Functional brain organization for visual search in ASD.Journal of the International Neuropsychological Society, 14(6), 990-1003.
Knickmeyer, R. C., & Baron-Cohen, S. (2006). Fetal testosterone and sex differences.Early Human Development,82(12), 755-760.
Knickmeyer, R. C., Wang, J. P., Zhu, H. T., Geng, X. J.,Woolson, S., Hamer, R. M.,... Gilmore, J. H. (2014).Impact of sex and gonadal steroids on neonatal brain structure.Cerebral Cortex, 24(10), 2721-2731.
Knickmeyer, R. C., Wheelwright, S., & Baron-Cohen, S.(2008). Sex-typical play: Masculinization/Defeminization in girls with an autism spectrum condition.Journal of Autism and Development Disorders, 38(6), 1028-1035.
Krahn, T. M., & Fenton, A. (2012). The extreme male brain theory of autism and the potential adverse effects for boys and girls with autism.Journal of Bioethical Inquiry, 9(1),93-103.
Kreiser, N. L., & White, S. W. (2014). ASD in females: Are we overstating the gender difference in diagnosis?Clinical Child and Family Psychology Review, 17(1), 67-84.
Lai, M.-C., Lombardo, M. V., Suckling, J., Ruigrok, A. N.,Chakrabarti, B., Ecker, C., … Bullmore, E. T. (2013).Biological sex affects the neurobiology of autism.Brain,136(9), 2799-2815.
Lawson, J., Baron-Cohen, S., & Wheelwright, S. (2004).Empathising and systemizing in adults with and without Asperger syndrome.Journal of Autism and Development Disorders, 34(3), 301-310.
Lindell, A. K., & Hudry, K. (2013). Atypicalities in cortical structure, handedness, and functional lateralization for language in autism spectrum disorders.Neuropsychology Review, 23(3), 257-270.
Matson, J. L., Worley, J. A., Fodstad, J. C., Chung, K. M.,Suh, D., Jhin, H. K., … Furniss, F. (2011). A multinational study examining the cross cultural differences in reported symptoms of autism spectrum disorders: Israel, South Korea, the United Kingdom, and the United States of America.Research in A utism Spectru m Disor ders, 5(4),1598-1604.
Meyer, L. R. (2008). Toy choice in an autistic female.Ohio Journal of Science, 108(1), 18.
Morsanyi, K., Primi, C., Handley, S. J., Chiesi, F., & Galli, S.(2012). Are systemizing and autistic traits related to talent and interest in mathematics and engineering? Testing some of the central claims of the empathizing-systemizing theory.British Journal of Psychology, 103(4), 472-496.
Murphy, C. M., Deeley, Q., Daly, E. M., Ecker, C., O'Brein,F. M., Hallahan, B., … Murphy, D. G. (2012). Anatomy and aging of the amygdala and hippocampus in autism spectrum disorder: An in vivo magnetic resonance imaging study of Asperger syndrome.Autism Research, 5(1), 3-12.
O'Riordan, M., Plaisted, K., Driver, J., & Baron-Cohen, S.(2001). Superior visual search in autism.Journal o f Experimental Psychology:Human Perception and Performance, 27(3), 719-730.
Parellada, M., Penzol, M. J., Pina, L., Moreno, C.,González-Vioque, E., Zalsman, G., & Arango, C. (2014).The neurobiology of autism spectrum disorders.European Psychiatry, 29(1), 11-19.
Preslar, J., Kushner, H. J., Marino, L., & Pearce, B. (2014).Autism, lateralisation, and handedness: A review of the literature and meta-analysis.Laterality, 19(1), 64-95.
Prigge, M. B. D., Lange, N., Bigler, E. D., Merkley, T. L.,Neeley, E. S., Abildskov, T. J.,... Lainhart, J. E. (2013).Corpus callosum area in children and adults with autism.Research in Autism Spectrum Disorders, 7(2), 221-234.
Rivet, T. T., & Matson, J. L. (2011). Review of gender differences in core symptomatology in autism spectrum disorders.Research in A utism Spectrum Disorders, 5(3),957-976.
Roelfsema, M. T., Hoekstra, R. A., Allison, C., Wheelwright,S., Brayne, C., Matthews, F. E., & Baron-Cohen, S. (2012).Are autism spectrum conditions more prevalent in an information-technology region? A school-based study of three regions in the Netherlands.Journal of Autism and Development Disorders, 42(5), 734-739.
Rojas, D. C., Camou, S. L., Reite, M. L., & Rogers, S. J.(2005). Planum temporale volume in children and adolescents with autism.Journal o f A utism and Developmental Disorders, 35(4), 479-486.
Ruigrok, A. N. V., Salimi-Khorshidi, G., Lai, M.-C.,Baron-Cohen, S., Lombardo, M. V., Tait, R. J., & Suckling,J. (2014). A meta-analysis of sex differences in human brain structure.Neuroscience & Biobehaviora l R eviews,39, 34-50.
Shaffer, D. R., & Kipp, K. (2013).Developmental psychology:Childhood and adolescence.(Zou, et al. Trans.). Beijing:China Light Industry Press. (Original work published 2002)
Sowell, E. R., Peterson, B. S., Kan, E., Woods, R. P., Yoshii,J., Bansal, R., … Toga, A. W. (2007). Sex differences in cortical thickness mapped in 176 healthy individuals between 7 and 87 years of age.Cerebral Cortex, 17(7),1550-1560.
Stanfield, A. C., McIntosh, A. M., Spencer, M. D., Philip, R.,Gaur, S., & Lawrie, S. M. (2008). Towards a neuroanatomy of autism: A systematic review and meta-analysis of structural magnetic resonance imaging studies.European Psychiatry,23(4), 289-299.
Tager-Flusberg, H. (2008). Cognitive neuroscience of autism.Journal of the International Neuropsychological Society,14(6), 917-921.
Takeuchi, H., Taki, Y., Thyreau, B., Sassa, Y., Hashizume, H.,Sekiguchi, A., … Kawashima, R. (2013). White matter structures associated with empathizing and systemizing in young adults.NeuroImage, 77, 222-236.
Treffert, D. A. (2011).Islands of genius:The bountiful mind of the autistic, acquired, and sudden savant.London, UK:Jessica Kingsley Publishers.
Valla, J. M., Ganzel, B. L., Yoder, K. J., Chen, G. M., Lyman,L. T., Sidari, A.P., … Belmonte, M. K. (2010). More than maths and mindreading: Sex differences in empathizing/systemizing covariance.Autism Research, 3, 174-184.
Valla, K., & Halazonetis, D. J. (2014). Correlation of 2D:4D digit ratio and craniofacial shape in prepubertal children.American Journal of Human Biology, 26(3), 337-346.
Wakabayashi, A., Baron-Cohen, S., Uchiyama, T., Yoshida,Y., Kuroda, M., & Wheelwright, S. (2007). Empathizing and systemizing in adults with and without autism spectrum conditions: Cross-cultural stability.Journal of Autism and Development Disorders, 37(10), 1823-1832.
Wallace, G. L. (2008). Neuropsychological studies of savant skills: Can they inform the neuroscience of giftedness?Roeper Review, 30(4), 229-246.
Warrier, V., Baron-Cohen, S., & Chakrabarti, B. (2013).Genetic variation in GABRB3 is associated with Asperger syndrome and multiple endophenotypes relevant to autism.Molecular Autism, 4(1), 48.
Wen, W., & Wen, S. W. (2014). Expanding upon the ‘extreme male brain’ theory of autism as a common link between other major risk factors: A hypothesis.Medical Hypotheses, 82(5), 615-618.
Wilke, M., Krägeloh-Mann, I., & Holland, S. K. (2007).Global and local development of gray and white matter volume in normal children and adolescents.Experimental Brain Research, 178(3), 296-307.
