紫翅猪毛菜、钠猪毛菜不同个体大小繁殖分配差异及随海拔的变化
2015-01-19刘尊驰刘华峰孙园园郝晓冉
刘尊驰, 刘华峰, 赵 丹, 罗 宁, 孙园园, 郝晓冉, 刘 彤
石河子大学生命科学学院, 石河子 832003
紫翅猪毛菜、钠猪毛菜不同个体大小繁殖分配差异及随海拔的变化
刘尊驰, 刘华峰, 赵 丹, 罗 宁, 孙园园, 郝晓冉, 刘 彤*
石河子大学生命科学学院, 石河子 832003
以新疆准噶尔盆地藜科猪毛菜属植物紫翅猪毛菜(SalsolaaffinisC. A. Mey)、钠猪毛菜(SalsolanitrariaPall)为研究对象,用繁殖分配比例的方法对比分析了两种猪毛菜不同海拔同一种群内不同个体大小繁殖分配的特点,并用异速生长模型分析了不同海拔繁殖生物量与营养生物量之间分配与个体大小的依赖关系。结果发现:1)不同海拔繁殖生物量(R)与营养生物量(V)呈不同程度的异速生长。紫翅猪毛菜随海拔的升高R-V的异速生长斜率显著升高,截距随海拔的升高没有显著增加;而钠猪毛菜的斜率随海拔升高显著降低,截距则显著升高。2)紫翅猪毛菜在较低海拔个体大小与繁殖分配呈负相关,在较高海拔呈正相关;钠猪毛菜在较低海拔个体大小与繁殖分配呈正相关,在较高海拔呈负相关;两种猪毛菜繁殖分配的适应对策相反。3)将同一种群个体大小分成大、中、小3种类型,多重比较发现紫翅猪毛菜在较低海拔,中小个体的繁殖分配显著高于大个体的繁殖分配;在较高海拔,大个体的繁殖分配显著高于中小个体的繁殖分配。钠猪毛菜在较低海拔,大个体的繁殖分配显著高于中、小个体的繁殖分配;在较高海拔,小个体的繁殖分配显著高于大、中个体的繁殖分配。综合分析认为:两个物种随海拔变化产生不同的繁殖分配策略,除遗传效应外,环境和个体大小对钠猪毛菜繁殖分配的变化均产生重要影响,而紫翅猪毛菜繁殖分配的变化主要由海拔差异导致。由于微生境对同一种群的个体大小产生影响,进而产生不同的繁殖分配模式,所以在干旱区更应重视个体大小对繁殖分配的影响。
准噶尔盆地; 海拔; 猪毛菜; 繁殖分配; 异速生长
从环境中获取资源,并对资源进行配置是植物生长的基本特性。当环境条件发生变化,资源受限制时,植物各种功能器官(生长、繁殖和防御等)对有限资源存在竞争问题,植物必须权衡这些功能器官之间的资源分配[1-3],资源分配模式很大程度上反映了植物生活史对策[4-5]。植物的繁殖对策是通过调整资源分配模式,以最佳的分配策略适应变化的环境[6-7],提高植物的适合度[8]。因而繁殖分配是物种生活史进化的结果,但也能够反映植物对当前环境的适应性特征[9-10]。不同物种繁殖分配模式对比研究有助于揭示植株繁殖对策的适应性进化特点,分析植物对环境变化的响应规律。
目前,繁殖分配的研究大多在种群层次上,然而种群是异质的,存在着个体大小、年龄组成等种群统计学特征。尤其在干旱区,降水稀少,微生境差异(例如不同坡位、低洼处、树冠下等)均会导致水分、光照和温度变化,进而对植株生长和繁殖产生作用,所以在干旱区即使同一海拔下由于微生境变化也会产生个体大小问题,但至今关于干旱区不同个体大小的繁殖分配研究非常少。
植物的繁殖分配是整体特征协调的反应,受自身遗传因素的影响,同时也受环境作用下的表型可塑性影响。植物对环境变化表现出3种程度的可塑性:第一,为表观可塑性,主要由植物大小变化导致,各器官之间的比例保持不变;第二,为真实可塑性,主要是各器官之间的比例通过构件增生和局部生理性适应发生变化所致;第三,为这两种可塑性协调作用导致的综合可塑性响应[11]。因此植物繁殖分配的变化主要来源于两个方面:(1)由个体大小变化导致的繁殖体生物量变化,在种群水平常称“表观可塑性”,在种群以上称个体大小依赖的繁殖分配改变[12-15];(2)受异质性环境的影响,生长和繁殖权衡关系改变导致的繁殖分配变化,在种群水平常称“真实可塑性”,在种群以上水平常称非个体大小依赖的繁殖分配改变[11,16]。因此区分个体大小、环境以及二者协同作用下繁殖分配才可以更好地分析繁殖分配产生的原因。目前繁殖分配研究多采用异速生长模型[16-20],异速生长斜率和截距的变化代表着繁殖分配变化的3种机制:1)截距变化斜率不变,表明同一种群不同个体大小营养生物量与繁殖生物量存在固定的异速生长关系,繁殖分配变化主要是由个体大小变化所致;2)截距不变斜率变化,环境变化导致生长和繁殖的权衡关系发生改变,个体大小的影响不显著;3)斜率和截距都发生改变,个体大小影响显著以及生长和繁殖的权衡关系发生改变。
海拔是影响植物生长的综合因子,伴随着温度、水分等环境因子变化,对植物繁殖分配同样产生影响[21-23]。目前研究不同海拔繁殖分配的差异通常是以繁殖生物量与总生物量的比值进行计算的,然而植物的分配通常是异速的[11,15],受表型可塑性的影响,那么干旱区植物随海拔变化繁殖分配的异速生长特征怎样等问题研究较少。猪毛菜属是藜科大属,是新疆荒漠区植被的重要组成部分[24]。调查发现猪毛菜属中紫翅猪毛菜(SalsolaaffinisC. A. Mey)、钠猪毛菜(SalsolanitrariaPall)两种一年生草本植物在新疆准噶尔盆地海拔200—1100 m之间均有分布,但分布范围差异较大[25-27]。那么以上两个物种为适应海拔变化,同一种群不同个体大小繁殖分配特征?不同海拔下同一种群不同个体大小繁殖分配如何变化?两个物种的以上变化是否一致等问题极有必要深入研究,以剖析两个物种繁殖分配随着海拔变化的特点,揭示这两个物种繁殖分配对干旱区荒漠的适应差异。
1 材料与方法
1.1 研究区概况
研究区位于新疆准噶尔盆地周缘及腹地,海拔200—1100 m。本区属中温带气候,具有典型的大陆性干旱气候特征,平均气温为4—6 ℃,最冷月温度-20 ℃,最热月温度22—26 ℃,年温差42—44 ℃,日温差平均16 ℃以上,平均积雪深度为10—40 cm[28]。沙漠区年降水量约100 mm,蒸发量2000 mm以上。
研究区植被以旱生或超旱生型植物为主。植物种类稀少,群落结构简单,藜科、菊科、十字花科、豆科和禾本科的植物较常见[29]。
1.2 实验材料
本实验研究的两种猪毛菜均为猪毛菜属一年生草本植物,在我国分布于新疆北部。紫翅猪毛菜(S.affinisC. A. Mey):高10—40 cm,花期7—8月,果期8—10月,主要生长于砾质荒漠、平原荒漠,分布范围广泛;钠猪毛菜(S.nitrariaPall):高10—40 cm,花期7—8月,果期9—10月,集中分布于丘间沙地,少量分布于平原荒漠、及轻度盐碱地等[30]。
1.3 取样方法及生物量测定
2012年6月沿准噶尔盆地周边及腹地,选取紫翅猪毛菜、钠猪毛菜主要分布的11个区域为样地。因紫翅猪毛菜、钠猪毛菜成片生长,近而每个样方大小以植株自然种群大小作为取样面积,每个样地标记3—4个样方,每两个样地相距30 km以上(图1)。
2012年10—11月两物种果实成熟期,进行采样。根据每个样地植株的大小,将采集的植株平均分成大、中、小3种类型。调查各样方不同个体大小植株的数量,得出3种类型植株数量比(表1)。每类植株在不同样方随机取样,各取10株即每个样地共取样30株。将植株连根完整挖出,分株装入密封袋中带回实验室进一步处理。
将采取的每个植株分割成营养器官(根、茎和叶)、繁殖器官(果实=种子+苞片)两部分,在80 ℃条件下烘72 h,然后用1/10000电子天平测量各部分干重。

图1 紫翅猪毛菜、钠猪毛菜样地分布图Fig.1 Distribution map of S. affinis and S. nitraria
1.4 统计分析
繁殖生物量与营养生物量的关系用经典异速模型R=aVb,对数转换logR= loga+blogV进行分析[31]。采用标准主轴回归(Standardized Major Axis,SMA)计算不同海拔V-R异速生长方程的截距loga和斜率b,若回归斜率与1有显著差异,表明为异速生长,若与1没有显著差异,表明为等速生长[32];根据Pitman的方法计算斜率的置信区间[33]。检验不同海拔间的斜率异质性,若斜率是异质的,则对不同海拔的斜率进行post-hoc多重比较[34]。以上分析均采用R软件SMATR Version 2.0来计算[35]。所有统计检验的显著水平均采用P= 0.05。
繁殖分配 = 繁殖器官生物量(干重)/植株总生物量(干重)[36]
采用SPSS19.0分析同一海拔不同个体大小繁殖分配的差异,首先用非参数检验 (Nonparametric Test)中1-Sample K-S检验数据是否为正态分布,对于正态分布的数据进行大、中、小个体繁殖分配的方差同质性检验,用单因素方差分析(One-way ANOVA)检验3组数据的差异性(显著水平设为0.05),对不服从正态分布的用K Independent Samples(Kruska -l Wallis H)进行比较,采用Origin8.5软件做图。
2 结果与分析
2.1 紫翅猪毛菜、钠猪毛菜样地特征
表1可见,紫翅猪毛菜、钠猪毛菜不同样地同一种群的个体大小比例不同,有的样地差异很大,个体大小在种群内变化较大。
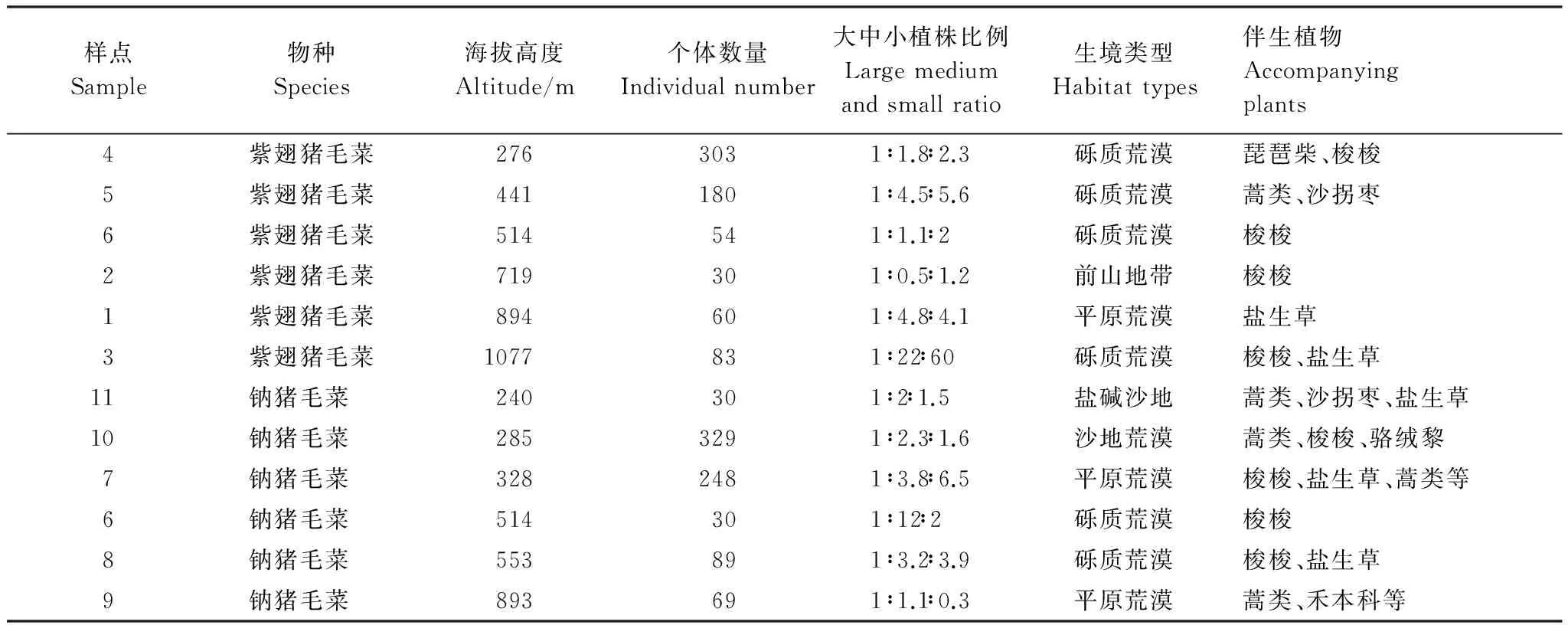
表1 紫翅猪毛菜、钠猪毛菜样地特征
2.2 不同海拔的繁殖分配模式
由表2可见,紫翅猪毛菜、钠猪毛菜不同海拔繁殖生物量与营养生物量的异速生长斜率均呈极显著异质性。多重比较得紫翅猪毛菜较高海拔异速生长的斜率显著大于较低海拔,除海拔276 m和514 m两样地的斜率与1没有显著差异外,即繁殖生物量与营养生物量等速生长,其他样地斜率与1均呈显著和极显著差异。钠猪毛菜较低海拔异速生长的斜率显著大于较高海拔,海拔240 m的斜率极显著大于1,海拔514 m和893 m处的斜率显著和极显著小于1,其他海拔的斜率与1没有显著差异。
2.3 不同海拔logR与logV的关系
图2可见,较低海拔与较高海拔相比,紫翅猪毛菜较高海拔大个体繁殖分配相对较高,较低海拔小个体的繁殖分配相对较高;不同海拔生长的植株繁殖分配进行比较发现,较高海拔植株的繁殖分配相对较高,海拔719 m植株的繁殖分配最高。钠猪毛菜较高海拔小个体繁殖分配较高,较低海拔大个体繁殖分配较高;不同海拔生长的植株繁殖分配进行比较发现,钠猪毛菜较高海拔的繁殖分配比较低海拔高。

表2 紫翅猪毛菜、钠猪毛菜繁殖生物量与营养生物量的分配模式

图2 紫翅猪毛菜、钠猪毛菜不同海拔logR与logV的回归图Fig.2 logR-logV regression of S. affinis and S. nitraria at different altitudesR: 繁殖生物量Repproductive biomass;Y: 营养生物量Vegetative biomass
2.4 海拔与斜率和截距的关系
图3可见,钠猪毛菜R-V的异速生长斜率与海拔呈显著负相关,截距与海拔呈显著正相关,表明钠猪毛菜较高海拔与较低海拔相比,较高海拔小个体的繁殖分配较高;紫翅猪毛菜随海拔的升高斜率显著增加,而截距呈增加趋势,较高海拔与较低海拔相比,较高海拔大个体的繁殖分配较高。

图3 紫翅猪毛菜、钠猪毛菜的斜率和截距与海拔的关系Fig.3 The slope and intercept for logR-logV regression vs altitude of S. affinis and S. nitraria
2.5 不同海拔不同个体大小的繁殖分配特点
图4可见,紫翅猪毛菜和钠猪毛菜不同海拔同一种群的繁殖分配随个体大小变化呈现不同的变化趋势。紫翅猪毛菜在较低海拔,繁殖分配与个体大小呈负相关关系,在较高海拔繁殖分配与个体大小呈正相关关系;钠猪毛菜在较低海拔繁殖分配与个体大小呈正相关关系,在较高海拔繁殖分配与个体大小呈负相关关系。

图4 不同海拔繁殖分配与个体大小的关系Fig.4 The relationship of reproductive allocation and individual size at different altitudes
图5可见,紫翅猪毛菜在较低海拔,中小个体的繁殖分配显著高于大个体的繁殖分配。在较高海拔,大个体的繁殖分配显著高于中小个体的繁殖分配。不同海拔下,同一种群不同个体大小与繁殖分配的变化趋势不同。
图6可见,钠猪毛菜在较高海拔的繁殖分配高于较低海拔。在较低海拔,大个体的繁殖分配显著高于中、小个体的繁殖分配。在较高海拔,小个体的繁殖分配显著高于大、中个体的繁殖分配。同样表明不同海拔同一种群不同个体大小与繁殖分配的变化趋势不同,且这种变化趋势与紫翅猪毛菜相反。
3 讨论
本研究发现钠猪毛菜不同海拔下的斜率和截距均有显著差异,说明钠猪毛菜繁殖分配的变化受个体大小与海拔协同作用的影响;紫翅猪毛菜不同海拔种群间斜率有显著差异,但截距间没有显著差异,表明海拔变化导致紫翅猪毛菜生长和繁殖的权衡关系发生改变,个体大小的影响不显著。植物为适应不同的生境,不同海拔样地内所有植株的logR与logV的回归斜率变化没有固定模式,研究结果有斜率没有显著大于1的[14],也

图5 紫翅猪毛菜不同海拔不同平均个体大小繁殖分配 Fig.5 The reproductive allocation of average individual size of S. affinis at different altitudes

图6 钠翅猪毛菜不同海拔不同平均个体大小繁殖分配 Fig.6 The reproductive allocation of average individual size altitudes of S. nitraria at different altitudes
有显著大于1和显著小于1同时存在的[37],本研究的紫翅猪毛菜、钠猪毛菜斜率的变化与后者研究一致,表明两种猪毛菜为适应不同的时空异质性生境而产生了完全不同的繁殖策略,繁殖分配表现出丰富的可塑性反应。
R与V异速生长的斜率反映了同一种群中不同个体大小植株营养生物量转化为繁殖生物量的成效[37],对于本研究而言,斜率越大说明营养生长分配给繁殖的比例也越大。本研究发现随海拔的升高,紫翅猪毛菜logR与logV的斜率显著增加,表明较低海拔与较高海拔相比,较低海拔小个体繁殖转化率更高,较高海拔大个体的繁殖转化率更高。钠猪毛菜logR与logV的斜率与海拔呈显著负相关,表明较低海拔与较高海拔相比,较低海拔大个体的繁殖成效更高,较高海拔小个体的繁殖成效更高。两物种随海拔的升高,斜率变化不同,表明繁殖分配的适应策略完全不同,可以确定这主要是由于物种间遗传差异所致。
截距反映了logR与logV二维回归变量空间的位置,可以作为二者关系的整体聚集度。如果同一种群所有个体繁殖生物量增加,则截距也会相应地增加,即反映相同个体大小下营养生物量转化成繁殖生物量整体效率情况[37]。本结果表明钠猪毛菜随海拔的升高截距显著增加,说明与较低海拔相比,较高海拔种群内所有个体的繁殖转化率较高。紫翅猪毛菜的截距变化趋势不明显,表明该物种个体的繁殖转化率对海拔变化的响应较低。
本研究发现紫翅猪毛菜和钠猪毛菜不同海拔同一种群内不同个体大小与繁殖分配关系变化不同。紫翅猪毛菜在较低海拔,繁殖分配与个体大小呈负相关关系,在较高海拔繁殖分配与个体大小呈正相关关系;钠猪毛菜在较低海拔繁殖分配与个体大小呈正相关关系,在较高海拔繁殖分配与个体大小呈负相关关系。将同一种群不同大小植株分成大、中、小3种类型,研究发现紫翅猪毛菜在较低海拔,中、小个体的繁殖分配显著高于大个体的繁殖分配;在较高海拔,大个体的繁殖分配显著高于中小个体的繁殖分配。钠猪毛菜较高海拔的繁殖分配高于较低海拔的繁殖分配;大个体在较低海拔的繁殖分配显著高于中小个体的繁殖分配;在较高海拔,小个体的繁殖分配显著高于大、中个体的繁殖分配。不同海拔紫翅猪毛菜、钠猪毛菜不同个体大小与繁殖分配之间的变化无固定模式,这可能是由于各个海拔生境差异较大,而在同一海拔内植株生长的微生境(温度、湿度及土壤肥力等)有较大的差异,因而产生了不同表型可塑性反应,对植株的繁殖产生重要影响。
在准噶尔盆地两个物种分布的200—1100 m海拔下,降水稀少,且降雨时间和量波动较大,两个物种多分布于地势低洼处、灌丛下等,导致了温度、湿度及土壤肥力的生境差异,这些微生境的变化不仅造成不同样地紫翅猪毛菜、钠猪毛菜植株大小的变化,也造成了植株大小比例的不同。研究发现紫翅猪毛菜、钠猪毛菜同一海拔不同个体大小植株的繁殖分配差显著异,且对海拔变化的响应趋势也不同,这可能是导致两个物种分布范围差异的重要原因。
综上,钠猪毛菜繁殖分配的变化主要是个体大小和海拔的共同作用,而紫翅猪毛菜繁殖分配的变化主要由海拔变化导致,个体大小的影响不显著,两个物种在不同海拔产生了不同的繁殖对策。同一海拔同一种群内由于微生境作用,也会产生不同大小植株,进而形成不同的繁殖分配模式,所以在干旱区研究植物的繁殖分配更应该重视个体大小对繁殖分配的影响。
致谢:石河子大学生命科学学院研究生于航、陈正霞和曾勇帮助野外调查采样,特此致谢。
[1] Obeso J R. The costs of reproduction in plants. New Phytologist, 2002, 155(3): 321-348.
[2] Niu K C, Schmid B, Choler P, Du G Z. Relationship between reproductive allocation and relative abundance among 32 species of a Tibetan alpine meadow: effects of fertilization and grazing. PLoS ONE, 2012, 7(4): e35448.
[3] 徐波, 王金牛, 石福孙, 高景, 吴宁. 青藏高原东缘野生暗紫贝母生物量分配格局对高山生态环境的适应. 植物生态学报, 2013, 37(3): 187-196.
[4] Bostock S J, Benton R A. The reproductive strategies of five perennial Compositae. The Journal of Ecology, 1979, 67(1): 91-107.
[5] Brenchley J L, Raven J A, Johnston A M. A comparison of reproductive allocation and reproductive effort between semelparous and iteroparous fucoids (Fucales, Phaeophyta). Hydrobiologia, 1996, 326-327(1): 185-190.
[6] Fabbro T, Körner C. Altitudinal differences in flower traits and reproductive allocation. Flora-Morphology, Distribution, Functional Ecology of Plants, 2004, 199(1): 70-81.
[7] Sawrey O R P. Effect of Altitude and Co-occurring Species on Sequential Reproductive Stages in a Grassland Harebell [D]. Dunedin: University of Otago, 2013.
[8] 何维明, 钟章成. 植物繁殖对策的概念及其研究内容. 生物学杂志, 1997, 14(6): 1-3.
[9] Harper J L. Population Biology of Plants. London: Academic Press, 1977.
[10] 牛克昌, 赵志刚, 罗燕江, 杜国祯. 施肥对高寒草甸植物群落组分种繁殖分配的影响. 植物生态学报, 2006, 30(5): 817-826.
[11] Weiner J. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology, Evolution and Systematics, 2004, 6(4): 207-215.
[12] Niu K C, Choler P, Zhao B B, Du G Z. The allometry of reproductive biomass in response to land use in Tibetan alpine grasslands. Functional Ecology, 2009, 23(2): 274-283.
[13] Bonser S P, Aarssen L W. Interpreting reproductive allometry: individual strategies of allocation explain size-dependent reproduction in plant populations. Perspectives in Plant Ecology, Evolution and Systematics, 2009, 11(1): 31-40.
[14] Weiner J, Campbell L G, Pino J, Echarte L. The allometry of reproduction within plant populations. Journal of Ecology, 2009, 97(6): 1220-1233.
[15] McConnaughay K D M, Coleman J S. Biomass allocation in plants: ontogeny or optimality? A test along three resource gradients. Ecology, 1999, 80(8): 2581-2593.
[16] Qin X L, Weiner J, Qi L, Xiong Y C, Li F M. Allometric analysis of the effects of density on reproductive allocation and Harvest Index in 6 varieties of wheat (Triticum). Field Crops Research, 2013, 144: 162-166.
[17] Cheplick G P, Reekie E G, Bazzaz F A. The allometry of reproductive allocation.// Reproductive Allocation in Plants. Pittsburgh: Academic Press, 2005: 97-128.
[18] asienski M, Bazzaz F A. The fallacy of ratios and the testability of models in biology. Oikos, 1999, 84(2): 321-326.
[19] Karlsson P S, Mendez M. The resource economy of plant reproduction //Reekie E, Bazzaz F A. Reproductive Allocation in Plants. San Diego: Academic Press, 2005: 1-40.
[20] Tian D H, Pan Q M, Simmons M, Chaolu H, Du B H, Bai Y F, Wang H, Han X G. Hierarchical reproductive allocation and allometry within a perennial bunchgrass after 11 years of nutrient addition. PLoS ONE, 2012, 7(9): e42833.
[21] Milla R, Giménez-Benavides L, Escudero A, Reich P B. Intra-and interspecific performance in growth and reproduction increase with altitude: a case study with two Saxifraga species from northern Spain. Functional Ecology, 2009, 23(1): 111-118.
[22] Sakai A, Sasa A, Sakai S. Do sexual dimorphisms in reproductive allocation and new shoot biomass increase with an increase of altitude? A case of the shrub willow Salix reinii (Salicaceae). American Journal of Botany, 2006, 93(7): 988-992.
[23] 李冰, 刘左军, 赵志刚, 胡春, 任红梅, 伍国强. 海拔对钝裂银莲花不同花色居群间繁殖特征及繁殖分配的影响. 草业学报, 2013, 22(1): 10-19.
[24] 毛祖美, 张佃民. 新疆北部早春短命植物区系纲要. 干旱区研究, 1994, 11(3): 1-26.
[25] 李宽亮, 刘彤, 李勇冠, 邵丽萍, 张荣, 宋占丽, 刘华峰. 褐翅猪毛菜种子异型性及其萌发特性研究. 石河子大学学报: 自然科学版, 2012, 30(3): 276-281.
[26] 罗宁, 刘彤, 刘华峰, 刘尊驰, 汤景光. 准噶尔荒漠紫翅猪毛菜和散枝猪毛菜种子萌发特性的差异. 石河子大学学报: 自然科学版, 2014, 32(5): 608-614.
[27] 刘华峰, 刘彤, 罗宁, 陈正霞, 刘尊驰. 紫翅猪毛菜果实异型性及个体大小依赖的空间变化. 石河子大学学报: 自然科学版, 2013, 31(6): 729-735.
[28] 潘晓玲, 张宏达. 准噶尔盆地植被特点与植物区系形成的探讨. 中山大学学报论丛, 1996, (2): 97-101.
[29] 孔令韶, 郭柯, 王其兵. 新疆南准噶尔荒漠优势植物的化学成分含量特点. 生态学报, 2002, 22(8): 1202-1210.
[30] 毛祖美. 新疆植物志. 乌鲁木齐: 乌鲁木齐科技卫生出版社, 1994.
[31] Klinkhamer P G L, Meelis E, De Jong T J, Weiner J. On the analysis of size-dependent reproductive output in plants. Functional Ecology, 1992, 6(3): 308-316.
[32] Warton D I, Wright I J, Falster D S, Westoby M. Bivariate line-fitting methods for allometry. Biological Reviews, 2006, 81(2): 259-291.
[33] Pitman E J G. A note on normal correlation. Biometrika, 1939, 31: 9-12.
[34] Warton D I, Weber N C. Common slope tests for bivariate errors-in-variables models. Biometrical Journal, 2002, 44(2): 161-174.
[35] Falster D S, Warton D I, Wright I J. User′s guide to SMATR: Standardised Major Axis Tests & Routines Version 2. 0. 2006. http://www.bio.mq.edu.au/ecology/SMATR/.
[36] Bazzaz F A, Ackerly D D, Reekie E G. Reproductive allocation in plants // Fenner M. Seeds: The Ecology of Regeneration in plant Communities. 2nd ed. Wallingford: CABI Publishing, 2000: 1-29.
[37] Guo H, Weiner J, Mazer S J, Zhao Z G, Du G Z, Li B. Reproductive allometry in Pedicularis species changes with elevation. Journal of Ecology, 2012, 100(2): 452-458.
Influence of altitude and difference of different-sized individuals on reproductive allocation inSalsolaaffinisC. A. Mey. andSalsolanitrariaPall.
LIU Zunchi, LIU Huafeng, ZHAO Dan, LUO Ning, SUN Yuanyuan, HAO Xiaoran, LIU Tong*
CollegeofLifeScience,ShiheziUniversity,Shihezi832003,China
In this study, we focused on two plant species in the genusSalsola(Chenopodiaceae),SalsolaaffinisC. A. Mey. andSalsolanitrariaPall., which are distributed in the Junggar Basin, Sinkiang, northwest China. We compared the characteristic of within-population reproductive allocation of different-sized plants (large, medium, and small) ofS.affinisandS.nitrariaat different altitudes using the ratios (the percentage of total biomass in reproductive biomass). In addition, we analyzed the relationship between reproductive biomass (R) and vegetative biomass (V) and the size-dependent allocation at different altitudes using a classical allometric model. We obtained the following results. (1) There were different levels of allometry between theRandVat different altitudes. The allometric slope (exponent) of theR-Vrelationship increased significantly with increasing altitude forS.affinis; however, there was a significant negative relationship forS.nitraria. We also found a significant positive relationship between the intercept of the regression of logRon logVand elevation forS.nitraria, although the relationship was not significant among populations ofS.affinis. (2) There was a negative correlation at lower altitude and a positive correlation at higher altitude for the relationship between individual size and reproductive allocation inS.affinis. Conversely, there was a positive correlation at lower altitude and a negative correlation at higher altitude for the relationship between individual size and reproductive allocation inS.nitraria. This indicates that theseSalsolaspecies exhibit contrasting adaptation strategies. (3) Plants within the same population ofS.affinisandS.nitrariawere divided into three size classes, large, medium, and small. Multiple comparisons showed that, at lower altitudes, the reproductive allocation of small and medium-sized individuals ofS.affiniswas significantly higher than that of large individuals, but that at higher altitudes, the reproductive allocation of large individuals was significantly higher. In contrast, the reproductive allocation of large individuals ofS.nitrariawas significantly higher than that of medium-sized and small individuals at lower altitudes, whereas the reproductive allocation of small individuals was significantly higher than that of medium-sized and large individuals at higher altitudes. Comprehensive analysis suggests that twoSalsolaspecies adopt different reproductive allocation strategies concomitant with a change in elevation. In addition to the genetic effect, the environment and individual size had significant influences on the variation in reproductive allocation ofS.nitraria, whereas individual size inS.affinishad no significant influence on the variation in reproductive allocation with altitude. The microenvironment of the arid Junggar Basin region also had an impact on individual plant size in the same population, and produced different reproductive allocation patterns. We therefore suggest that more attention should be paid to the impact of individual size on reproductive allocation in arid areas.
Junggar Basin; altitude;Salsola; reproductive allocation; allometry
国家自然科学基金项目(41061004); 国家自然科学基金委-新疆联合基金重点项目(U1130304/L10)
2014-01-15;
日期:2014-11-19
10.5846/stxb201401150116
*通讯作者Corresponding author.E-mail: betula@126.com
刘尊驰, 刘华峰, 赵丹, 罗宁, 孙园园, 郝晓冉, 刘彤.紫翅猪毛菜、钠猪毛菜不同个体大小繁殖分配差异及随海拔的变化.生态学报,2015,35(18):5957-5965.
Liu Z C, Liu H F, Zhao D, Luo N, Sun Y Y, Hao X R, Liu T.Influence of altitude and difference of different-sized individuals on reproductive allocation inSalsolaaffinisC. A. Mey. andSalsolanitrariaPall..Acta Ecologica Sinica,2015,35(18):5957-5965.
