天目山针阔混交林林木空间特性
2015-01-08范佩佩韦新良郭如意汤孟平
范佩佩,韦新良,郭如意,汤孟平
(1.浙江农林大学环境与资源学院,浙江 临安311300;2.浙江农林大学 浙江省森林生态系统碳循环与固碳减排重点实验室,浙江 临安311300)
天目山针阔混交林林木空间特性
范佩佩1,2,韦新良1,2,郭如意1,2,汤孟平1,2
(1.浙江农林大学环境与资源学院,浙江 临安311300;2.浙江农林大学 浙江省森林生态系统碳循环与固碳减排重点实验室,浙江 临安311300)
为了解天目山针阔混交林林木空间的特性,以期对该地区森林可持续经营模式的实践提供理论基础,设置了15块30 m×30 m的固定标准地,采用实测调查法,调查样地内所有胸径≥5 cm的林木特征值。以林木冠幅作为空间分割尺度来生成Voronoi图,确定林木空间结构单元,统计分析林木空间不同尺度上的差异性。结果表明:单株树种的空间面积差异性较大,群组中每株树木周围平均有4~5株树。优势种中苦槠Castanopsis sclerophylla,锥栗Castanea henryi和麻栎Quercus acutissima等树种的空间面积大小较稳定,而马尾松Pinus massoniana,杉木Cunninghamia lanceolata,白栎Quercus fabric和山矾Symplocos caudata等树种空间面积大小差异性较大。林木胸径的大小对其所占空间面积有一定影响,而对其周围邻近木的株数影响不大。目标树的最近邻木株数为3~13株,多数为5~7株,平均为6株。天目山针阔混交林为强度混交林,林分群落结构相对稳定。图8表2参25
森林生态学;针阔混交林;Voronoi图;空间结构;混交度;天目山
森林功能与森林结构密切相关。森林空间结构体现了树木在林地上的分布格局及其属性在空间上的排列方式,在很大程度上决定了林分的稳定性、发展的可能性和经营空间的大小[1]。林分结构是森林经营和分析中的一个重要因子,是对林分发展过程的综合反映[2]。以林分空间结构为目标结合非空间结构要求,调控空间结构,能为林分空间结构优化调控提供新途径[3]。目前,空间结构分析已成为国际上天然林经营模拟技术的主要研究内容[4-5]。林分的树种结构、大小组成、空间格局等是反映林分结构的重要方面[6]。通过调整林分结构以达到发挥森林多功能的理念越来越受到重视,并且在表达、分析、模拟、重建林分结构方面已有大量研究[7-8]。对林木的空间及其特性进行研究,了解和掌握森林中林木对空间的利用程度,对优化森林结构,合理配置森林树种及其空间位置以加强森林营造与恢复等森林经营管理具有重要的理论指导意义和实践应用价值。
1 研究地区和材料
1.1 研究地区
天目山国家级自然保护区地处浙江省临安市西北部(30°18′30″~30°24′55″N,119°24′11″~119°28′21″E),海拔为300~1 556 m,具有典型的中亚热带的森林生态系统和森林景观,森林植被十分茂盛。该区年平均气温为8.8~14.8℃,雨水充沛,年降水量为1 390~1 870 mm,年相对湿度为76%~81%,积雪期较长,形成浙江西北部的多雨中心。天目山土壤随着海拔升高由亚热带红壤向湿润的温带型棕黄壤过渡。这些独特的环境条件构成了天目山植物区系的古老性、复杂性和种类丰富性,共计有苔类植物70种、藓类植物240种、蕨类植物110种、种子植物1 570种,其中国家重点保护植物25种,以天目山命名的植物24种[12]。
天目山针阔混交林中乔木层共有49种树种,物种丰富度较高[13]。每个森林群落结构因子只与森林景观的个别因子之间有一定的相关性[14]。树高、郁闭度、枝下高、草本层盖度、色调、树种组成、分布状况和采伐剩余物等8个因子与森林景观效果相关显著[15]。针阔混交林中枫香Liquidambar formosana的种内种间竞争随着对象木胸径的变大而逐渐变小,在种群内呈聚集状[16-17]。
1.2 研究材料
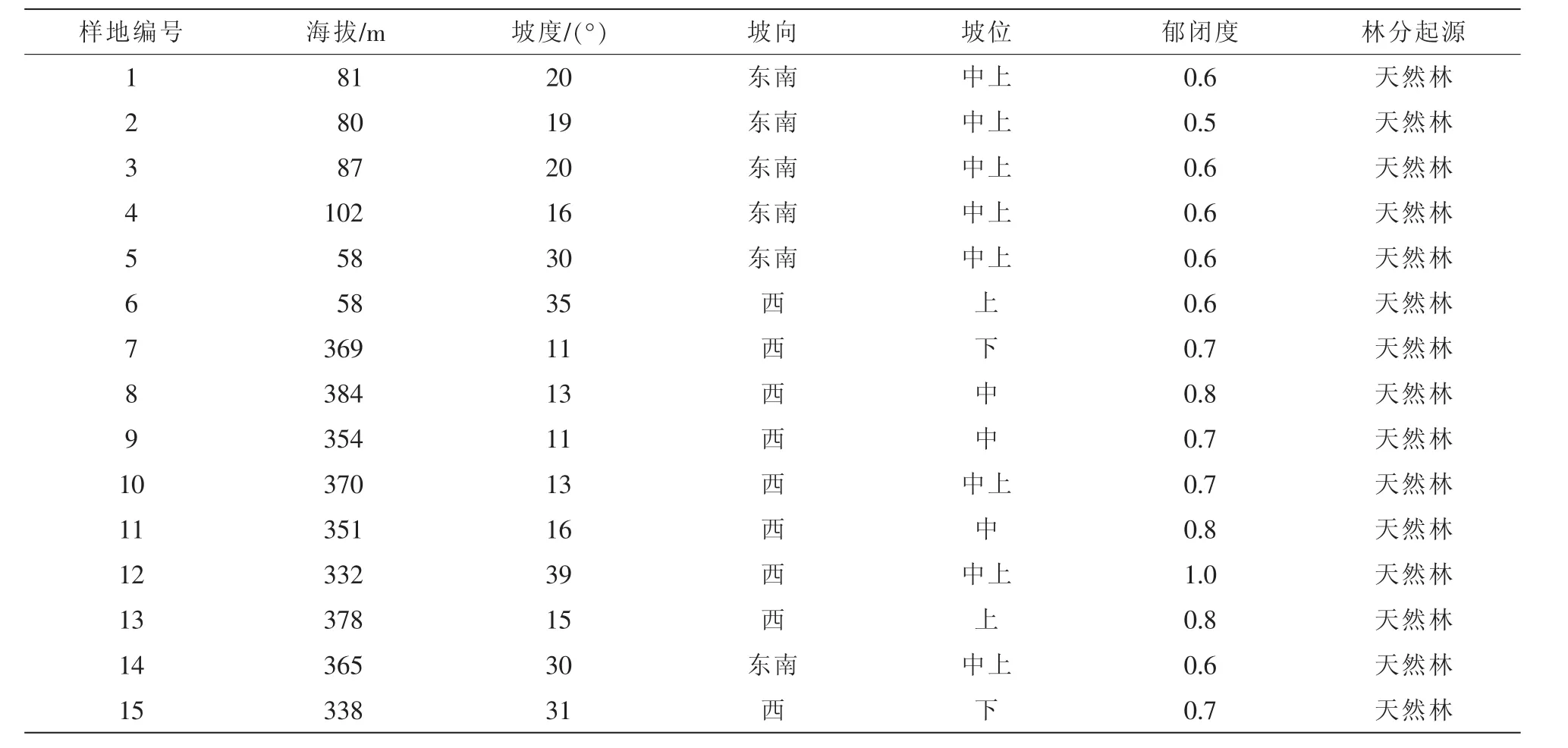
表1 样地概况Table 1 Situation of sample plots
研究所选样地系浙江省天目山区针阔混交林研究样地,建立于2011年。在样地设置时,踏查了全林分,了解林分界线和特点,选择没有人为干扰且具有典型性和代表性,树种类型多样,同时易于进行外业调查的天然林地,设置了15个30 m×30 m样地。采用全面调查法(表1),调查样地内所有内胸径≥5 cm的林木特征值,包括树种、胸径、树高、枝下高、冠幅等,同时用全站仪测定并记录每棵林木的具体坐标。
经调查统计,天目山针阔混交林包含的树种多是浙江省乡土树种,具有代表性及地域性。针叶树多为马尾松Pinus massoniana,杉木Cunninghamia lanceolata,偶有金钱松Pseudolarix amabilis,柳杉Cryptomeria fortunei;阔叶树以枫香,短柄枹Quercus glandulifera var.brevipetiolata,白栎Quercus fabri,苦槠Castanopsis sclerophylla,山矾Symplocos caudata,锥栗Castanea henryi,麻栎Quercus acutissima,青冈Cyclobalanopsis glauca,黄连木Pistacia chinensis,木荷Schima superba,化香Platycarya strobilacea,山合欢Albizia kalkora,石栎Lithocarpus glabra,黄檀Dalbergia hupeana,小叶栎Quercus chenii等为主,偶有无患子Sapindus mukorossi,野桐Mallotus lour,天目槭Acer sinopurpurascens,檫树Sassafras tzumu,华东楠Machilus leptophylla,石楠Photinia serrulata,榉树Zelkova schneideriana等其他树种;灌木层主要是檵木Loropetalum chinense,山胡椒Lindera glauca和山樱Prunus serrulata等。
2 研究过程与方法
2.1 林木空间单元界定
林木的生长主要通过树叶的光合作用积累生物量所形成,树叶的空间及其潜在伸展空间对林木的生长至关重要。由树叶和枝条构成的树冠是树木的营养体。树冠的长度、宽度和高度体现了树木进行光合作用的有效载体和光的截获能力,是决定生产效率的重要指标。因此,林木的空间即是指林分中林木树冠的体积量及其所占据的生态位置与潜在发展态势。水平方向上,树冠间的相互关系主要有相离、相切、相交等3种形态,均可用冠幅来表示。林分中的林木空间主要取决于林木间树冠的相互关系,因此,为了更加准确地体现林木之间的空间关系,本研究在基于冠幅权重的基础上,进行编程分析生成新的Voronoi多边形来划分空间,得出的每个林木的多边形可以反映出林木个体大小差异以及林木之间相互影响的空间关系,即可代表林木空间。
2.2 林木空间单元测度
2.2.1 Voronoi图的构建 Voronoi图以诸多地理空间实体作为生长目标将整个连续空间剖分为若干个Voronoi多边形。Voronoi图中的空间实体与Voronoi多边形一一对应,常用Voronoi多边形确定空间实体的影响范围[18]。利用VS开发平台,调用C#开源类库进行编程分析,再运用Arcgis进行数据展示来生成Voronoi图。①把样地内每1株树都看做1个点,平面上n个点便成为n个Voronoi多边形的中心。②选定某一点作为参考点,以该点为起点做与其他n-1个点的连线,根据式(1)和式(2)确定连线上的分割点,再过该点作这n-1根连线的垂线,这些垂直相交构成一些多边形,但是只有那些离参考点最近的垂线围成的多边形才是所需要的,这个多边形内只有1个参考点。重复以上过程,依次用其他n-1个点做参考点形成n-1个多边形,从而构成Voronoi网络模型。编程要点是,林木所在的Voronoi区域能够体现该林木实际所占据的空间范围以及与其相邻的周边林木之间的空间关系。
2.2.2 林木空间分割点的确定 为简便和精确地反映出林木的空间特征,以样地内每株林木为生长目标,以林木冠幅(林木的垂直投影面积)作为空间分割尺度。依据式(1)和式(2)通过计算机运算确定空间分割点的位置:

式(1)和式(2)中: Wa和Wb分别为树木a和树木b的冠幅,La和Lb分别为树木a和树木b所代表的点到2点连线上分割点的距离,Lab为树木a和树木b所代表2点间的距离。
2.2.3 林木空间单元的划分及面积计算 林木空间单元按2个尺度进行测度:①单株林木空间尺度:以单株林木的空间为一个尺度,既每株树所对应的Voronoi图中的多边形。②群组林木空间尺度:以单株林木空间和与其相邻的周边的林木空间的组合为一个尺度,即单株树所对应的多边形和与其多边形相邻的林木所在多边形的空间组合。空间面积计算:基于ArcGis,根据面数据的geometry属性可计算得出空间面积。
2.3 边缘校正
处于样地边缘的林木空间结构单元是不完整的,会给分析带来误差,称之为边缘效应[19]。为了消除边缘效应,本研究采用八邻域样地法进行边缘校正,即分别在原样地的上、左、下、右、左上、右上、左下、右下等8个方向复制原样地,形成由9个相同样地组成的一个新的大样地。这样每个对象木都有一个相对完整的结构单元,减少理论值与实际值之间的偏差,消除边缘影响[20]。
2.4 混交度
混交度用来反映林分中树种空间隔离程度,为中心木的n株邻近木中与中心木不属同种个体所占的比例, 计算公式为[21-22]:

式(3)中:Mi为第i株中心木的点混交度;n为邻近木株数;vij为离散性变量,当对象木i与第j株最近相邻木非同种时vij=1,反之,vij=0。
在此将Mi的取值划分为5个区间:0.00,(0.00,0.25),(0.25,0.50),(0.50,0.75),(0.75,1.00),分别对应于混交度的定性描述:零度混交、弱度混交、中度混交、强度混交和极强度混交。
3 结果分析
3.1 林木空间尺度特征
3.1.1 林木空间尺度总体特征 基于Voronoi图分别计算出单株以及群组所占的空间面积,单株林木空间面积中,最大为120.81 m2,最小为0.03 m2,平均为7.60 m2;群组林木空间面积中,最大为342.79 m2,最小为0.48 m2,平均为34.25 m2。群组林木空间面积的均值是单株的4.5倍,说明群组中每株树木周围平均有4~5株树。由图1和图2可以看出:无论单株还是群组的面积最大值均远远大于平均值,由表2可以看出:单株林木空间比群组林木空间面积的变异系数大,说明单株的空间面积差异性更大,这是由于群组中目标树受最近邻木的影响使得总体面积分布较平均。
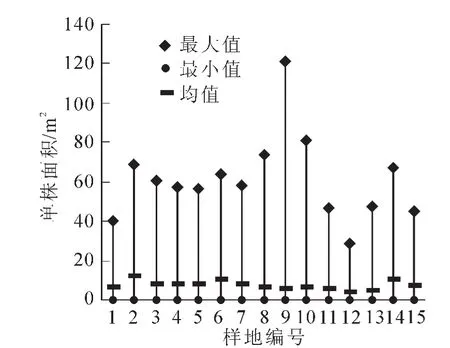
图1 单株面积大小统计特征值Figure 1 General statistics of tree area size

图2 群组面积大小统计特征值Figure 2 General statistics of group area size

表2 不同空间尺度变异系数Table 2 Coefficient of variation of different space scales
3.1.2 林木优势种空间尺度特征 采用重要值法计算,按照重要值大小排列得出13种优势种分别为:马尾松、杉木、枫香、短柄枹、白栎、苦槠、山矾、锥栗、麻栎、黄连木、木荷、化香、山合欢[13],并计算出单株和群组优势种所占的平均空间面积。由图3和图4可以看出:单株优势种中,平均面积最大的是白栎,最小的是麻栎;群组优势种中,平均面积最大的是黄连木,最小的是苦槠。优势种中苦槠、锥栗和麻栎等3种树种的空间面积大小较稳定,最大值与最小值相对接近平均值;马尾松、杉木、白栎和山矾等4种优势树种中均存在一些个体占据远大于平均值的空间面积,存在这种现象的原因可能是由于该优势种具有较大的冠幅以及较强的竞争力。

图3 单株优势种面积大小统计特征值Figure 3 General statistics of tree area size of dominant species

图4 群组优势种面积大小统计特征值Figure 4 General statistics of group area size of dominant species
3.1.3 优势种空间与胸径关系分析 由图5可以看出:马尾松和黄连木的平均胸径较大,白栎和黄连木的平均空间面积较大;从整体来看,各优势种的平均胸径变化趋势与平均空间面积变化趋势是相似的。说明林木的胸径大小与其所占空间面积有一定的相关性。存在一些树种有较大平均胸径而其所占空间面积相对较小,或者平均胸径较小而其所占空间面积相对较大;例如马尾松虽然胸径较大但其冠幅相较于同等平均胸径的阔叶类树种(如黄连木)要小一些,所以其所占空间面积也相对较小,反之亦然,如白栎。除了冠幅,还与树种的年龄、树高、混交度等因素有关。
3.2 林木空间邻近木特征
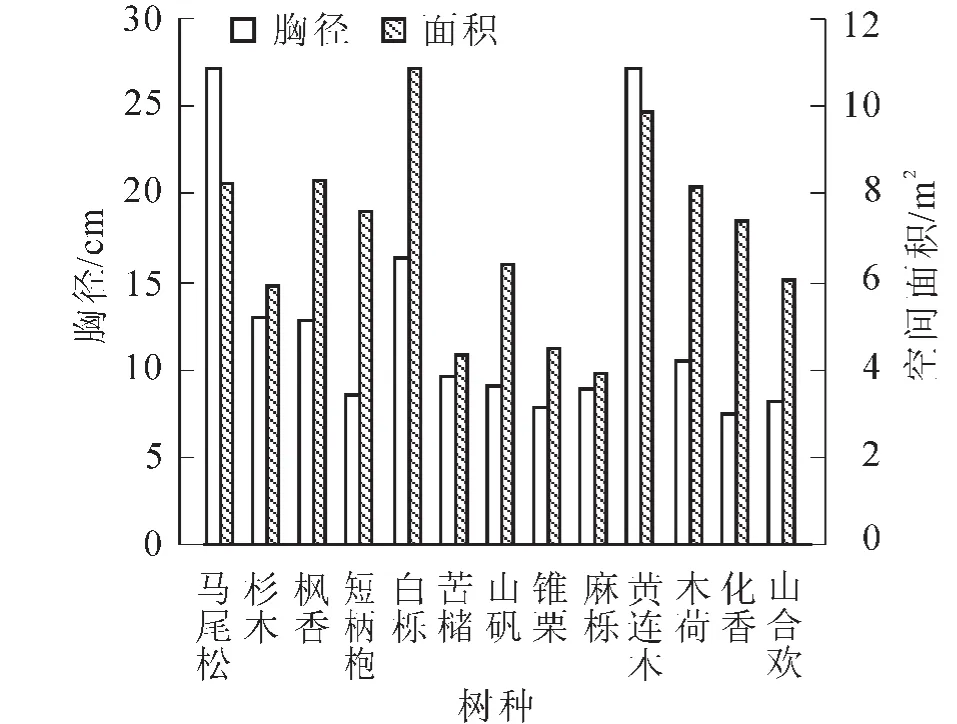
图5 各优势种平均面积与平均胸径Figure 5 Mean area and mean DBH of dominant species
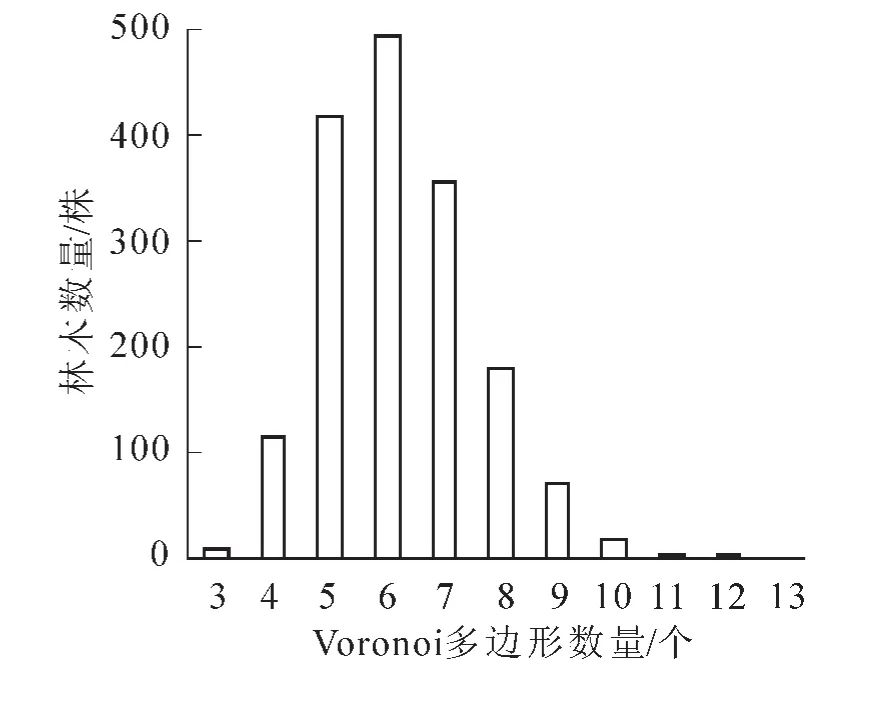
图6 Voronoi多边形边数统计值Figure 6 General statistics of Voronoi polygon edge number
根据Voronoi图的特点,每个Voronoi多边形内仅包含1株林木。目标树的最近邻木株数与其所在Voronoi多边形的边数相等。在目标树周围的林木中,最近邻木对目标树的生长有最直接的影响。基于Voronoi图可以确定每株目标树的最近邻木的株数。图6表明:目标树的最近邻木株数为3~13个,有11种可能的取值,多数为5,6和7株,平均为6株。这个结果与天目山近自然毛竹Phyllostachys edulis林空间结构与生物量的关系结果基本一致[23],说明不同类型的森林存在相似的空间结构特征。计算得出各个样地Voronoi多边形边数的变异系数均在0.20上下浮动,说明各样地分布情况相同,总体分布在6株上下波动,较均匀。由图7可以看出:平均邻近木株数并不随平均胸径变化而变化,平均每株树木周围有6株邻近木,可见胸径的大小对于不同树种的邻近木株数影响不大。
3.3 林木空间树种混交特征
天然林树种丰富、混交复杂。用混交度能够充分反映各样地林分的树种隔离程度,计算得出所有样地林分的平均混交度为0.711,因此,所调查的针阔混交林为强度混交林。图8是各样地林分的平均混交度,可以看出1,2和6号样地混交度较低,属于中度混交,其余均属于强度混交。这说明天目山针阔混交林林分中,同树种聚集在一起的情况不多,多数树种不与本树种为伴,大多数林木均为混交生长,林分群落结构相对稳定。总体来看,邻近木组成的结构越多样,树种孤立的情况越少,林分的稳定性也会相应增强。

图7 各优势种平均邻近木株数与平均胸径Figure 7 Mean nearest tree number and mean DBH of dominant species
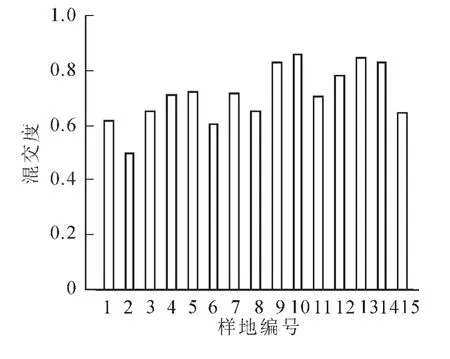
图8 各样地平均混交度Figure 8 Mean mingling in each plots
4 结论与讨论
单株林木空间面积中,最大为120.81 m2,最小为0.03 m2,平均为7.60 m2;群组林木空间面积中,最大为342.79 m2,最小为0.48 m2,平均为34.25 m2。群组中每株树木周围平均有4~5株树。由于群组中目标树受最近邻木的影响使得总体面积分布较平均,单株的空间面积差异性更大。单株优势种中,平均面积最大的是白栎,最小的是麻栎;群组优势种中,平均面积最大的是黄连木,最小的是苦槠。优势种中苦槠、锥栗和麻栎等3种树种的空间面积大小较稳定,而马尾松、杉木、白栎和山矾等4种树种空间面积大小差异性较大。从整体来看,林木的平均胸径变化趋势与平均空间面积变化趋势是一致的,具有相关性。由于树种的冠幅、年龄、树高、混交度等因素的影响,存在一些胸径与空间面积变化不一致的树种,如马尾松和白栎。
天目山针阔混交林为强度混交林,林分中同树种聚集在一起的情况不多,多数树种不与本树种为伴,大多数林木均为混交生长,林分群落结构相对稳定。总体来看,邻近木组成的结构越多样,树种孤立的情况越少,林分的稳定性也会相应增强。目标树的最近邻木株数为3~13株,多数为5,6和7株,平均为6株。各样地分布情况相同,总体分布在6株上下波动,较均匀。林木胸径的大小对于其周围邻近木株数影响不大。
林分空间结构决定林木之间的生长竞争态势及其空间生态位,对森林未来的发展具有决定性作用[24]。这些结果对天目山针阔混交林的结构和功能,以及实现区域森林类型与结构的优化配置,促进森林资源的可持续性和多效利用有一定参考意义。林木空间结构存在的差异性与很多因素有关,如林分密度、林分年龄、树种及植被群落结构等,还需要开展进一步的研究。地理信息系统(GIS)有强大的空间分析功能,通常被用于大尺度的森林景观结构分析[25]。本研究表明,地理信息系统(GIS)同样是林分小尺度上分析和提取林木空间结构信息的有效工具。
[1] 岳永杰,余新晓,李钢铁,等.北京松山自然保护区蒙古栎林的空间结构特征[J].应用生态学报,2009,20(8):1811-1816.
YUE Yongjie,YU Xinxiao,LI Gangtie,et al.Spatial structure of Quercus mongolica forest in Beijing Songshan Mountain Nature Reserve[J].Chin J Appl Ecol,2009,20(8):1811-1816.
[2] 雷相东,唐守正.林分结构多样性指标研究综述[J].林业科学,2002,38(3):140-146.
LEI Xiangdong,TANG Shouzheng.Indicators on structural diversity within stand:a review[J].Sci Silv Sin,2002,38(3):140-146.
[3] 汤孟平,徐文兵,陈永刚,等.毛竹林空间结构优化调控模型[J].林业科学,2013,49(1):120-126.
TANG Mengping,XU Wenbing,CHEN Yonggang,et al.Spatial structure optimizing adjustment and control model of Phyllostachys edulis stand[J].Sci Silv Sin,2013,49(1):120-126.
[4] 惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究[J].林业科学研究,2001,14(1):23-27.
HUI Gangying,HU Yanbo.Measuring species spatial isolation in mixed forests[J].For Res,2001,14(1):23-27.
[5] 黄运峰,丁易,臧润国,等.海南岛霸王岭热带低地雨林树木的空间格局[J].植物生态学报,2012,36(4):269-280.
HUANG Yunfeng,DING Yi,ZANG Runguo,et al.Spatial pattern of trees in tropical lowland rain forest in Bawangling of Hainan Island[J].Chin J Plant Ecol,2012,36(4):269-280.
[6] 方国景,汤孟平.天目山常绿阔叶林优势种群胸径的空间连续性分析[J].浙江农林大学学报,2014,31(5):663-667.
FANG Guojing,TANG Mengping.Spatial continuity for DBH in dominant populations of an evergreen broadleaved forest in national nature reserve of Mount Tianmu[J].J Zhejiang A&F Univ,2014,31(5):663-667.
[7] 王宏翔,惠刚盈,张弓乔,等.基于空间结构调查的林分密度估计[J].应用生态学报,2014,25(7):1912-1918.
WANG Hongxiang,HUI Gangying,ZHANG Gongqiao,et al.Stand density estimation based on the measurement of spatial structure[J].Chin J Appl Ecol,2014,25(7):1912-1918.
[8] 刘小恺,刘茂松,黄峥,等.宁夏沙湖4种干旱区群落中主要植物种间关系的格局分析[J].植物生态学报,2009,33(2):320-330.
LIU Xiaokai,LIU Maosong,HUANG Zheng,et al.Pattern analysis of inter-specific relationships in four arid communities in sand lake,Ningxia Hui Autonomous Region[J].Chin J Plant Ecol,2009,33(2):320-330.
[9] 惠刚盈.基于相邻木关系的林分空间结构参数应用研究[J].北京林业大学学报,2013,35(4):1-8.
HUI Gangying.Studies on the application of stand spatial structure parameters based on the relationship of neighborhood trees[J].J Beijing For Univ,2013,35(4):1-8.
[10] 惠刚盈,GADOW K,ALBERT M.一个新的林分空间结构参数—大小比数[J].林业科学研究,1999,12(1):1-6.
HUI Gangying,GADOW K,ALBERT M.A new parameter for stand spatial structure neighborhood comparison[J]. For Res,1999,12(1):1-6.
[11] 惠刚盈,李丽,赵中华,等.林木空间分布格局分析方法[J].生态学报,2007,27(11):4717-4729.
HUI Gangying,LI Li,ZHAO Zhonghua,et al.The comparison of methods in analysis of the tree spatial distribution pattern[J].Acta Ecol Sin,2007,27(11):4717-4729.
[12] 汤孟平,周国模,施拥军,等.天目山常绿阔叶林优势种群及其空间分布格局[J].植物生态学报,2006,30(5):743-752.
TANG Mengping,ZHOU Guomo,SHI Yongjun,et al.Study of dominant plant populations and their spatial patterns in evergreen broadleaved forest in Tianmu Mountain[J].Chin J Plant Ecol,2006,30(5):743-752.
[13] 王敬,韦新良,徐建,等.天目山区针阔混交林树种测树因子特征研究[J].华东森林经理,2014,28(1):35-40.
WANG Jing,WEI Xinliang,XU Jian,et al.Survey of species measurement factor in Tianmu Mountain on coniferousbroadleaved mixed forest[J].East Chin For Manage,2014,28(1):35-40.
[14] 韦新良,何莹.森林景观效果生成因子的相关性[J].浙江农林大学学报,2011,28(5):701-705.
WEI Xinliang,HE Ying.A correlation study of generation factors for forest landscape effects[J].J Zhejiang A&F Univ,2011,28(5):701-705.
[15] 韦新良,何莹.生态景观林景观效果构成特性定量分析[J].西北林学院学报,2011,26(6):181-185.
WEI Xinliang,HE Ying.Quantive analysis on scenery effect factor landscape forest[J].J Northwest For Univ,2011, 26(6):181-185.
[16] 骆文建,韦新良,汤孟平,等.天目山枫香种内与种间竞争的数量研究[J].浙江农林大学学报,2012,29(5):641-646.
LUO Wenjian,WEI Xinliang,TANG Mengping,et al.Intraspecific and interspecific competition in Liquidambar formosana on Mount Tianmu[J].J Zhejiang A&F Univ,2012,29(5):641-646.
[17] 张志华,韦新良,汤孟平,等.天目山针阔混交林中枫香的结构特征[J].浙江农林大学学报,2012,29(6):867-874.
ZHANG Zhihua,WEI Xinliang,TANG Mengping,et al.Structural characteristics of Liquidambar formosana for a mixed coniferous-broadleaf forest in Mount Tianmu[J].J Zhejiang A&F Univ,2012,29(6):867-874.
[18] 陈军,赵仁亮,乔朝飞.基于Voronoi图的GIS空间分析研究[J].武汉大学学报:信息科学版,2003,28(增刊):32-37.
CHEN Jun,ZHAO Renliang,QIAO Chaofei.Voronoi diagram based GIS spatial analysis[J].Geom Info Sci Wuhan Univ,2003,28(supp):32-37.
[19] 陈新美,张会儒,姜慧泉.东北过伐林区蒙古栎林空间结构分析与评价[J].西南林学院学报,2010,30(6):20-24.
CHEN Xinmei,ZHANG Huiru,JIANG Huiquan.Analysis and evaluation on spatial structure of Quercus mongolica forests in over-logged reg ion in northeast China[J].J Southwest For Univ,2010,30(6):20-24.
[20] 郝月兰,张会儒,唐守正.Voronoi图方法确定云冷杉林最近邻木的适用性研究[J].林业资源管理,2011(6):59-65.
HAO Yuelan,ZHANG Huiru,TANG Shouzheng.Study on the adaptability of voronoi diagram on determining the nearest neighboring trees of natural mixed spruce forests[J].For Res Manage,2011(6):59-65.
[21] 安慧君,张韬.聚集指数边界效应的校正方法与应用[J].南京林业大学学报:自然科学版,2005,29(3):57-60.
AN Huijun,ZHANG Tao.Studies on adjustment approach and its application of bounder effect on aggregation index[J].J Nanjing For Univ Nat Sci Ed,2005,29(3):57-60.
[22] 王鹏,陈丽华,卞西陈,等.北沟林场天然次生林群落结构与种群分布格局[J].应用生态学报,2011,22(7):1668-1674.
WANG Peng,CHEN Lihua,BIAN Xichen,et al.Community structure and distribution pattern of a natural secondary forest in Beigou Forest Farm[J].Chin J Appl Ecol,2011,22(7):1668-1674.
[23] 汤孟平,徐文兵,陈永刚,等.天目山近自然毛竹林空间结构与生物量的关系[J].林业科学,2011,47(8):1-6.
TANG Mengping,XU Wenbing,CHEN Yonggang,et al.Relationship between spatial structure and biomass of a close-to-nature Phyllostachys edulis stand in Tianmu Mountain[J].Sci Silv Sin,2011,47(8):1-6.
[24] 郑丽凤,周新年,江希钿,等.松阔混交林林分空间结构分析[J].热带亚热带植物学报,2006,14(4):275-280.
ZHENG Lifeng,ZHOU Xinnian,JIANG Xidian,et al.Analysis of the stand spatial structure of Pinus massonianabroadleaved mixed forest[J].J Trop Subtrop Bot,2006,14(4):275-280.
[25] 陆元昌,洪玲霞,雷相东.基于森林资源二类调查数据的森林景观分类研究[J].林业科学,2005,41(2):21-29.
LU Yuanchang,HONG Lingxia,LEI Xiangdong.Study on classification of forest landscape components based on forest resource inventory data for management[J].Sci Silv Sin,2005,41(2):21-29.
Forest tree spatial characteristics of a coniferous,broad-leaf mixed forest on Mount Tianmu
FAN Peipei1,2,WEI Xinliang1,2,GUO Ruyi1,2,TANG Mengping1,2
(1.School of Environmental and Resource Sciences,Zhejiang A&F University,Lin’an 311300,Zhejiang,China;2. Zhejiang Provincial Key Laboratory of Carbon Cycling in Forest Ecosystems and Carbon Sequestration,Zhejiang A&F University,Lin’an 311300,Zhejiang,China)
To understand characteristics of forest tree spacing in a coniferous,broad-leaf mixed forest on Mount Tianmu and to provide a theoretical basis for sustainable forest management,fifteen 30 m×30 m fixed plots were established to measure characteristics of all trees over 5 cm DBH.The forest crown was used for a spatially segmented scale to generate a Voronoi Diagram,to determine the spatial unit between trees,and for statistical analysis of differences with various scales of tree spacing.Results showed that single tree spacing had large differences,with an average of four to five trees in a group.The spatial area was stable for dominant species of Castanopsis sclerophylla,Castanea henryi,and Quercus acutissima,but large spatial difference were found with Pinus massoniana,Cunninghamia lanceolata,Quercus fabric,and Symplocos caudata.DBH influenced spatial area,but not the nearest-tree number.The range of nearest-tree number for target trees was 3 to 13,with the mode being 5,6,and 7,and the mean was 6.Overall,Mount Tianmu was a well-established, mixed forest,with a relatively stable community structure.[Ch,8 fig.2 tab.25 ref.]
forest ecology;coniferous,broad-leaf mixed forest;Voronoi diagram;spatial structure;mingling; Mount Tianmu
S718.45
A
2095-0756(2015)05-0675-08
10.11833/j.issn.2095-0756.2015.05.004
2014-12-11;
2015-01-16
国家自然科学基金资助项目(31170595);浙江省公益技术研究项目(2013C33017)
范佩佩,从事森林及其环境评价与规划设计研究。 E-mail:980733599@qq.com。通信作者:韦新良,教授,博士,从事森林及其环境评价与规划设计研究。 E-mail:weixl@zafu.edu.cn
