拟南芥的种子及胚胎发育
2014-12-24苏国锋李从从
苏国锋 李从从
(1.正定县南楼乡人民政府,河北 正定050800;2.华北制药金坦生物技术股份有限公司,河北 石家庄050000)
拟南芥属于被子植物门,双子叶植物纲。 由于其植株小、结实多、生命周期短、基因组简单、遗传操作简便,成为遗传研究的主要模式生物。全世界有超过六千家实验室正在对拟南芥的生长发育及其对环境应答的过程开展深入研究。它在粮食增产、农作物耐逆、环境保护等领域做出了重要贡献。
地球上80%的绿色植物是被子植物,种子在被子植物生活史中占有重要地位,而胚胎是种子的主要组成部分。 因此对种子及胚胎的发育及其调控机制的研究意义重大。本文主要论述了模式植物拟南芥的种子及胚胎的发育。
1 拟南芥的种子发育过程
双子叶模式植物拟南芥的种子发育包括细胞分裂、器官起始及种子成熟三个主要过程(图1)(Agarwal et al., 2011)。
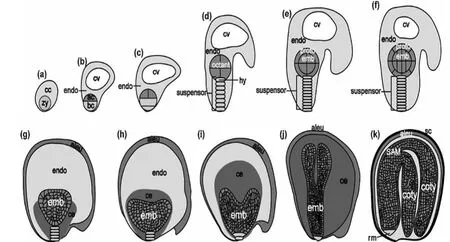
图1 拟南芥种子发育不同时期的示意图(Agarwal et al., 2011)Figure 1 Seed development stages in Arabidopsis(Agarwal et al., 2011)
在种子的发育过程中,合子的形成意味着种子发育的起始,拟南芥种子发育的细胞分裂阶段只有2 天,在此期间,完成了双受精及受精后合子到原胚的转化(图1a-d)。 所谓双受精是指在胚囊中一个精核和卵细胞融合形成二倍体的合子,将来发育成胚胎,第二个精核和中央细胞的两个极核融合将来发育成三倍体的胚乳。合子经第一次不对称分裂形成一个双核的胚和一个多核的胚乳。 接下来顶细胞(apical cell) 进行垂直方向的分裂,基部细胞(basal cell) 进行水平方向的分裂,胚乳核继续分裂形成一个多核体。到原胚后期,顶细胞分裂形成八细胞的胚,叫做八分体(octant)(Agarwal et al., 2011)。
受精后3-4 天,胚胎从球形胚发育到心形胚时期,器官的起始由此开始,同时胚乳开始细胞化(图1e-g)。 内层细胞分裂导致胚胎轴向的形成和局部分化。 胚胎从球形胚后期向心形胚转变时,整个胚胎大概有200 多个细胞, 胚胎周围的胚乳快速的细胞化, 形成糊粉层(aleurone)。 子叶原基及根分生组织(root meristem) 也是在这个时期形成的(Agarwal et al., 2011)。
受精后的5-24 天是器官的生长以及成熟时期(图1h-k),胚胎从心形胚发育到线形的子叶胚再到弯曲的子叶胚时期。 其间胚乳细胞经历了内复制和细胞的程序化死亡。 成熟的种子,是由糊粉层及被其包被着的弯曲的子叶胚构成的。 弯曲的子叶胚主要由子叶(cotyledon),茎顶端分生组织(shoot apical meristem)和根顶端分生组织组成。 整个种子被种皮(seed coat)包被并保护着,发育到种子成熟末期时,种子已经失水,蛋白质和RNA 合成停止,胚及种子进入休眠状态(Agarwal et al.,2011)。
拟南芥种子的发育可以分成三个阶段:形态发生,成熟,干燥。 在形态发生的过程中,大多数的细胞处于高度分裂分化的状态,此时,种子的主体是胚胎,主要由子叶、下胚轴、幼根及根和芽的顶端分生组织构成(Bowman and Floyd, 2008; Zhang and Ogas, 2009)。 在成熟的过程中,种子开始积累像脂质和蛋白质这样的储存物质。 当种子进入到干燥阶段时,种子脱水,新陈代谢几近停止,此时是保存种子的最佳时期,当外界有适合萌发的条件出现时,种子进入萌发状态。在胚胎发育的同时,胚乳快速增殖形成一个多核体,经历高度的细胞化过程,最后成熟,只剩下一层胚乳细胞(Zhang and Ogas, 2009)。
2 拟南芥的胚胎发育过程
据研究,植物胚胎发育的过程大致被划分为三个阶段(图2),第一阶段为双受精到原胚阶段, 从两细胞期到八细胞期被称为原胚阶段。合子经过不对称分裂产生两个大小不同的子细胞:一个小的、位于上部的顶细胞。一个大的、位于下部的基细胞,这样在胚胎的纵轴上就建立了极性,经过组织学的研究发现,顶细胞和基细胞发育成成熟胚的不同部位。 两细胞胚在经过各自的细胞分裂发育到四细胞期、八细胞期,完成原胚阶段的发育。第二阶段为球形胚到心形胚的转换阶段,从十六细胞期到心形胚时期被称为球形胚到心形胚的转换阶段,在此阶段胚胎的器官和组织类型发生分化,即纵向上顶部和基部的区分及胚胎的器官系统的产生。 第三阶段为器官的生长和成熟阶段,鱼雷胚期到成熟胚期为器官的生长和成熟阶段,此阶段为模式形成到储存物质积累阶段的一个过渡时期 (Goldberg et al., 1989; Goldberg et al.,1994; West and Harada, 1993)。
3 拟南芥种子及胚胎发育的影响因素
3.1 植物激素
生长素、脱落酸和赤霉素是常见的植物激素,研究表明,它们在种子的发育过程中发挥着重要的作用。在植物的模式建成和分化过程中生长素浓度梯度起着重要作用(Bowman and Floyd, 2008)。 生长素对于胚胎形态发生时模式的建成是必需的(Weijers and Jurgens, 2005)。 赤霉素对种子的发育至关重要, 赤霉素还可以促进种子的萌发(Zhang and Ogas, 2009)。在拟南芥中,使赤霉素失去活性的酶异位表达会导致种子的败育(Singh et al., 2002)。 与种子成熟有关系的是脱落酸,它和种子储存物质的积累和种子的休眠等都有关系 (Finkelstein et al.,2008)。
3.2 转录因子
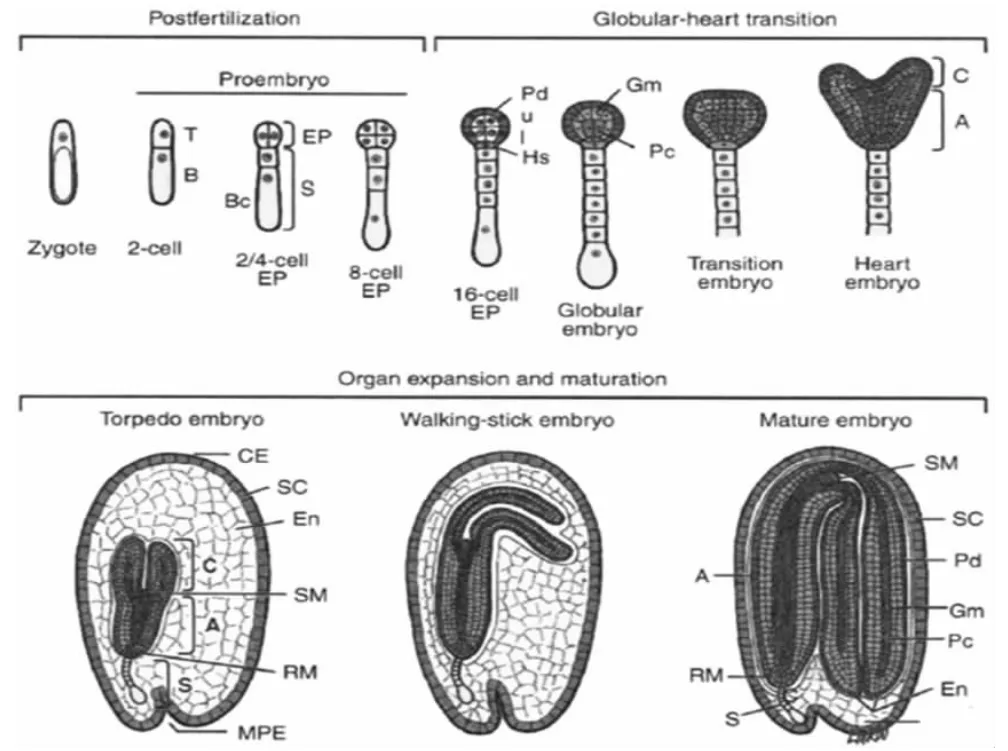
图2 拟南芥胚胎发育过程示意图(Goldberg et al., 1994)Figure 2 A generalized overview of Arabidopsis embryogenesis(Goldberg et al., 1994)
转录因子是植物种子发育过程中的重要调控因子。 在种子、胚胎发育的过程中是否也有转录因子发挥着重要作用呢? 据报道,胚胎发育的各个阶段都有相应的转录因子发挥着作用(图3)。

图3 种子发育中三个重要时期表达的转录因子(Agarwal et al. 2011)Figure 3 TFs from Arabidopsis expressed during the three landmark stages of seed development. (Agarwal et al. 2011)
3.2.1 原胚到球形胚发育的转录因子
通过突变体表型的观察和表达模式的分析,WOX 基因在胚胎发生早期的模式建成和特异区域的调控程序上都发挥着重要的作用。WOX2 和WOX8/STIMPY-LIKE/STPL 在受精的合子中表达。 WOX9/STIMPY/ STIP 先在基细胞中表达, 再在顶细胞中表达,WOX5 在静止中心表达(Abrash and Bergmann, 2009; Agarwal et al.,2011; Dumas and Rogowsky, 2008; Wu et al., 2007)。 MADS 家 族 的 八 个 基 因(AGL70/MAF3, AGL53, 93, 67, FLC, FLM,AGL15 和18) 对于胚胎的发育也很重要(Lehti-Shiu et al., 2005)。LEC1 是一个转录激活因子,主要调控着胚胎形态发生和细胞分化相关的一些基因(Agarwal et al.,2011)。
3.2.2 球形胚到心形胚转变时期发挥作用的转录因子
在拟南芥中,SHOOT MERISTEMLMLE SS/STM 对于SAM 的起始和维持是必须的(Agarwal et al.,2011)。 STM 需要和CUC1,CUC2 一起来调控形成SAM,STM 反过来需要CUC2 的功能, 在发育的胚胎中,CUC1 和CUC2 控制着邻近器官的分化。 它们调控着胚胎发生过程中的细胞分裂,并参与了营养生长阶段和胚胎阶段SAM 的形成。 CUC3和CUC1、CUC2 是同源的, 界定着分生组织和器官边界(Chandler et al., 2008)。 WUSCHEL/WUS 通过ZLL 控制干细胞的命运 (Long and Barton, 1998; Tucker et al., 2008)。
3.2.3 鱼雷胚到弯曲子叶胚时期参与的转录因子
ABI3 特异地在心形胚早期到种子成熟时期表达, 直至受精后16天达到一个稳定水平(Agarwal et al.,2011; Suzuki and McCarty, 2008)。另一个重要的调节因子就是FUS3,它通过抑制胚胎的生长、抑制早期的萌发、子叶细胞命运的建立、种子储存物质的积累等方面来控制种子的休眠(Tiedemann et al., 2008)。
4 结语
综上所述,迄今为止人们对于拟南芥胚胎发育的过程及影响因素的研究已取得了重大进展, 但是植物胚胎发育是一个非常复杂的、由很多基因,很多因素控制的一个过程,因此,进一步研究植物胚胎发育各时期基因表达的时空性及特异蛋白的表达, 对于揭示胚胎发生、发育的机理具有重大的理论和现实意义。
[1]Agarwal, P., Kapoor, S., and Tyagi, A.K.(2011) Transcription factors regulating the progression of monocot and dicot seed development [J]. Bioessays, 33 (3):189-202.
[2]Bowman, J.L., and Floyd, S.K. (2008) Patterning and polarity in seed plant shoots[J]. Annu Rev Plant Biol, 59:67-88.
[3]Zhang, H., and Ogas, J. (2009) An epigenetic perspective on developmental regulation of seed genes[J]. Mol Plant, 2(4):610-627.
[4]Goldberg, R.B., Barker, S.J., and Perez-Grau, L. (1989) Regulation of gene expression during plant embryogenesis[J]. Cell, 56(2):149-160.
[5]Goldberg, R.B., de Paiva, G., and Yadegari, R. (1994) Plant embryogenesis:zygote to seed[J]. Science, 266(5185):605-614.
[6]West, M., and Harada, J.J. (1993) Embryogenesis in Higher Plants: An Overview[J]. Plant Cell, 5(10):1361-1369.
[7]Weijers, D., and Jurgens, G. (2005) Auxin and embryo axis formation: the ends in sight?[J]. Curr Opin Plant Biol, 8(1):32-37.
[8]Singh, D.P., Jermakow, A.M., and Swain, S.M. (2002) Gibberellins are required for seed development and pollen tube growth in Arabidopsis[J]. Plant Cell, 14(12):3133-3147.
[9]Finkelstein, R., Reeves, W., Ariizumi, T., and Steber, C. (2008) Molecular aspects of seed dormancy[J]. Annu Rev Plant Biol, 59:387-415.
[10]Abrash, E.B., and Bergmann, D.C. (2009) Asymmetric cell divisions: a view from plant development[J]. Dev Cell, 16(6):783-796.
[11]Dumas, C., and Rogowsky, P. (2008) Fertilization and early seed formation[J].C R Biol, 331(10):715-725.
[12]Wu, X., Chory, J., and Weigel, D. (2007) Combinations of WOX activities regulate tissue proliferation during Arabidopsis embryonic development [J]. Dev Biol, 309(2):306-316.
[13]Lehti-Shiu, M.D., Adamczyk, B.J., and Fernandez, D.E. (2005) Expression of MADS-box genes during the embryonic phase in Arabidopsis[J]. Plant Mol Biol, 58(1):89-107.
[14]Chandler, J., Nardmann, J., and Werr, W. (2008) Plant development revolves around axes[J]. Trends Plant Science, 13(2):78-84.
[15]Long, J.A., and Barton, M.K.(1998) The development of apical embryonic pattern in Arabidopsis[J]. Development, 125(16):3027-3035.
[16]Tucker, M.R., Hinze, A., Tucker, E.J., Takada, S.,Jurgens,G., and Laux, T. (2008) Vascular signalling mediated by ZWILLE potentiates WUSCHEL function during shoot meristem stem cell development in the Arabidopsis embryo[J]. Development, 135(17):2839-2843.
[17]Suzuki, M., and McCarty, D.R. (2008) Functional symmetry of the B3 network controlling seed development[J]. Curr Opin Plant Biol, 11(5):548-553.
[18]Tiedemann, J., Rutten, T., Monke, G., Vorwieger, A., Rolletschek, H.,Meissner, D., Milkowski, C., Petereck, S., Mock, H.P., Zank, T., and BaumlMLein,H.(2008) Dissection of a complex seed phenotype: novel insights of FUSCA3 regulated developmental processes[J]. Dev Biol, 317(1):1-12.
