廉州湾红树蚬的繁殖生物学研究
2014-12-15周浩郎张俊杰杨明柳邢永泽
周浩郎, 张俊杰, , 杨明柳, , 邢永泽, 吴 斌, 阎 冰
(1.广西红树林研究中心广西红树林保护与利用重点实验室, 广西北海 536007; 2.广西大学动物科学技术学院, 广西南宁 530004)
红树蚬(Polymesoda erosa(Solander, 1768)), 广泛分布于热带、亚热带沿海红树林生境的双壳贝类,其软体部蛋白含量高, 味道鲜美, 是红树林周边居民日常赶海的常见渔获种类和家常菜肴。红树蚬营埋栖生活, 生长快, 个体大, 耐干露, 对环境适应能力强, 除了食用价值外, 还有通乳、利尿、清热和抗病毒等药用功效, 是值得发展养殖的贝类[1-3]。20世纪 90年代末在中国台湾云林县试养红树蚬获得成功,现已发展成为台湾云林的特色土产, 商品名“马蹄蛤”, 鲜贝及其深加工产品不仅在台湾本地销售, 也销往大陆地区和出口日本。近年来, 广东和广西红树林区已经出现了小规模的利用天然贝苗进行的红树蚬养殖。由于采捕红树蚬天然苗种较为困难而且往往会破坏红树林根系, 开展红树蚬人工育苗势在必行。然而关于红树蚬繁殖生物学的研究报道极少,Morton[4]研究了中国香港地区红树蚬的繁殖策略,Gimin等[5]研究了澳大利亚北部红树蚬的繁殖特性,Clemente等[6]研究了印度果阿Chorao岛红树蚬的性腺发育和生殖周期。本实验采集广西北海廉州湾红树蚬的周年样品进行性腺发育和生殖周期研究, 并通过人工授精观察其胚胎发育过程, 以期为北部湾地区红树蚬的人工繁殖和资源保护提供基础资料。
1 材料与方法
2011年8 月至2012年7月, 每月在广西北海市廉州湾红树林挖掘采集红树蚬。潮水高于平均海平面 360cm时采样点才能被海水淹没, 在采样点土壤10cm深处埋设防水温度记录仪(HOBO UA-002-08),设定每小时记录温度1次。
从每次采集的样品中随机抽取 20~30个个体,洗净擦干贝壳表面后用游标卡尺测量壳长、壳宽和壳高, 用解剖刀贴贝壳内壁剥离闭壳肌并开壳后观察性腺的饱满度, 将软体部与贝壳分开后放置于60~70℃烘箱中恒温24 h, 用电子天平(精确度0.001g)分别称量软体部干质量和贝壳干质量, 计算肥满度指数(软体部干质量/贝壳干质量×100%)。每月固定标本 10~15个, 切取不同部位的性腺, 用 Bouin’s液固定, 石蜡包埋, 切片厚7 µm, H.E 染色, 光学显微镜观察并拍照。2011年 9月底采集红树蚬样品, 壳长40 mm以上的个体开壳后用肉眼和显微镜进行性别判定和性腺形态观察, 壳长50~70 mm的个体, 选取性腺饱满、成熟度高的个体, 分别吸取精子和卵子,进行人工授精, 光学显微镜观察精卵形态、定时观察胚胎发育过程并拍照。
每月用温度记录仪自带的软件读取温度记录,统计计算当月表层土温的平均值。对照潮汐表(国家海洋信息中心编)统计采样期间采样点每月被海水浸淹的总时间(潮高大于 360 cm)和完全不能被海水浸淹的天数(全天潮高小于 360 cm), 计算每月红树蚬被海水浸淹时间的百分比。
2 结果
2.1 性别及性腺形态
红树蚬为雌雄异体, 322个成体红树蚬中雌性164个(50.93%), 雄性 156个(48.45%), 雌性同体 2个(0.62%), 雌雄比约为1∶1。从外形不能区分性别,即红树蚬的外形不具备性别二态性。成体性别可以通过性腺颜色来辨别, 雄性性腺淡乳黄色, 雌性性腺黑褐色(图1)。
红树蚬的生殖腺位于足上方、内脏团两侧, 属于滤泡型, 由滤泡、生殖输送管、生殖管3部分构成。精子属鞭毛型, 呈钉子状, 全长约 55~65 µm, 其中鞭毛长约 40~50 µm; 成熟卵呈圆球形, 属均黄卵,外被一层很厚的胶质膜, 胶质膜的内侧与卵膜形成很明显的分界, 卵黄直径约80~120 µm, 胶质膜直径约 170~220 µm(图2)。
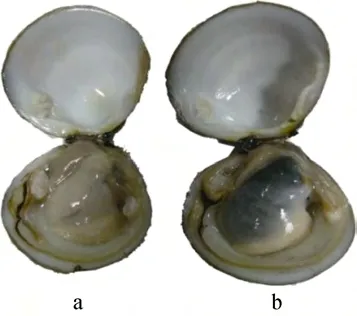
图1 红树蚬的性腺Fig.1 The gonads of P.erosa
2.2 性腺发育周年变化及分期
根据周年的活体和性腺组织切片观察, 红树蚬的性腺发育可以分为以下5个阶段。
2.2.1 增殖期
2月至3月, 滤泡出现并逐步增大, 滤泡壁增厚,呈不规则形状, 性腺开始形成, 出现卵原细胞和精原细胞排列于滤泡内壁(图3a, 图4a)。
2.2.2 生长期
4月至5月, 性腺明显增大, 基本覆盖整个内脏团, 部分个体肉眼能辨雌雄。滤泡增大增多, 滤泡间空隙逐渐缩小。精子堆积于滤泡内, 卵母细胞增大,附于滤泡内壁(图3b, 图4b)。
2.2.3 成熟期
5月至6月, 性腺非常饱满, 性别在外观颜色上区别明显, 雌性呈黑褐色, 雄性呈淡乳黄色。滤泡更大, 滤泡间的空隙基本消失, 滤泡腔充满成熟的生殖细胞。精子密集堆积, 占满滤泡, 头朝滤泡壁, 尾朝滤泡腔。卵细胞成熟, 近圆形, 大部分不再附于滤泡壁, 而是堆积在滤泡腔内(图3c, 图4c)。
2.2.4 排放期
6月至11月, 持续有部分个体产卵排精, 性腺饱满度降低, 颜色变淡。7月, 大部分个体集中出现产卵排精, 形成1次高峰, 11月出现一次小高峰。滤泡内的精子和卵细胞显著减少, 滤泡腔内出现较大的空隙, 部分成熟的精子或卵子尚未排放, 不同滤泡中存留的精卵数量差异较大, 滤泡壁上出现精原细胞和卵原细胞(图3d, 图4d)。
2.2.5 休止期
12月至翌年 1月, 大部分个体肉眼不能观察到性腺。精子和卵子几乎释放殆尽, 滤泡萎缩, 间隙增大, 滤泡壁变厚, 滤泡内仍有残留的少量精细胞、卵子或成熟的卵母细胞(图3e, 图4e)。

图3 红树蚬精巢组织的周年变化Fig.3 Annual testicular histological changes of P.erosa

图4 红树蚬卵巢组织的周年变化Fig.4 Annual ovary histological changes of P.erosa
2.3 生殖周期与肥满度
红树蚬每年一个生殖周期, 雌雄性腺发育同步,繁殖期从6月一直延续到11月。红树蚬的肥满度、栖息环境温度和受海水浸淹时间周年变化情况见图5。2月性腺发育开始进入增殖期, 肥满度指数最低(1.98), 1~2月温度为全年最低。4月进入生长期, 5~6月进入成熟期, 5月肥满度指数达到最大(5.19), 3~6月温度逐渐升高达到全年最高值。7月为繁殖高峰,肥满度指数显著降低, 10月为第2次繁殖高峰, 肥满度指数明显降低, 但降幅小于 7月。12月进入休止期, 肥满度指数较 11月略微上升, 然后迅速降低,温度从11月的 23.29℃降到 12月的 15.76℃再到翌年1月的13.14℃。廉州湾属不规则全日潮, 采样点位于高潮带, 每天被海水浸淹时间 0~7h, 海水浸淹时间占比 15.8%(2月)至 20.7%(8月), 每月有 4~8d完全不能被海水浸淹。
2.4 胚胎发育
红树蚬的卵为沉性卵, 在 pH 7.8, 水温 26℃的海水中, 其胚胎发育进程如表1和图6所示。在0.06‰氨水处理过的卵子中加入适量精子后, 精子与卵子迅速结合, 形成受精卵, 经约 0.3~1.47h, 出现第一极体, 第二极体。接着受精卵伸出极叶, 准备进行第一次卵裂。受精卵发育经约7h后形成囊胚, 9h后进入胚胎阶段, 胚胎表面出现纤毛以及主鞭毛,胚胎在卵膜内转动, 不断撞击卵膜, 之后成为担轮幼虫。经约 27~40h, 担轮幼虫顶部凹陷, 形成面盘,进入D形幼虫期。

图5 红树蚬肥满度、表层土温和受海水浸淹时间周年变化(2011年8月~2012年7月)Fig.5 Annual variation of P.erosa Condition index, surface soil temperature, and tidal inundation from Aug 2011 to July 2012

表1 红树蚬胚胎发育过程Tab.1 The Embryogenesis of P.erosa
3 讨论
3.1 红树蚬的繁殖期
廉州湾红树蚬的繁殖期从6月延续到11月, 较长的繁殖期与香港[4]、澳大利亚北部[5]和印度果阿Chorao岛[6]红树蚬较一致, 也与常见的滩涂双壳贝类文蛤[7]和青蛤[8]等相似。滩涂贝类的生活环境因子日变化较大, 特别是红树蚬有着半陆生的性质, 每天被海水浸淹又露空, 温度、盐度和溶氧等变化剧烈。较长的繁殖期可能是红树蚬应对剧烈变化的环境条件, 提高繁殖成功率的一种适应。在繁殖期内一直有个别个体少量排放精卵, 可能与环境因子剧烈变化的刺激有关。廉州湾红树蚬在一个生殖周期内有2次繁殖高峰(7月和10月), 这与香港红树蚬相一致, 但印度果阿Chorao岛红树蚬仅有1次繁殖高峰,说明不同地理种群的繁殖特性存在差异, 我们的前期研究也表明廉州湾红树蚬与澳大利亚北部红树蚬在形态上存在差异[9]。
3.2 环境因子、性腺发育和肥满度的关系
温度和饵料是影响双壳贝类生殖周期最重要的外因。在温度最低的1月, 性腺处于休止期, 肥满度次低。2月温度开始上升, 性腺发育启动, 进入增殖期, 肥满度最低。3~5月温度快速上升, 性腺迅速发育, 经生长期到达成熟期, 充满整个内脏团, 非常饱满, 肥满度达到最高。6月温度达到全年最高, 性腺发育进入排放期, 肥满度微降。7~11月温度维持在相对较高的水平(30.73~23.29℃), 性腺发育持续处于排放期, 肥满度波动但维持在相对较高的水平。12月温度快速下降到 15.76℃, 性腺开始萎缩, 进入休止期, 肥满度略微上升后迅速下降。性腺发育和肥满度随温度变化而变化的规律明显, 肥满度的变化相对滞后。
红树蚬为滤食性, 只有在被海水浸淹时才能摄食, 海水浸淹时间长短可以间接地反映其摄食量。1至 2月海水浸淹时间略微降低, 肥满度降低而性腺发育启动。2~4月海水浸淹时间略微升高但幅度极小(占比 15.8%至 16.4%), 性腺发育迅速, 肥满度显著升高, 5月海水浸淹时间小幅增加, 性腺发育最快,肥满度最大。6~12月海水浸淹时间占比在19%左右波动, 性腺持续发育并产卵排精直至萎缩, 肥满度波动。海水浸淹时间变化与性腺发育和肥满度之间没有明显的规律, 暗示红树蚬具有较强的摄食能力,较短的摄食时间即能满足发育需要。

图6 红树蚬胚胎发育Fig.6 Embryogensis of P.erosa
红树蚬的肥满度与性腺发育有密切的关系。性腺发育迅速的时期(3~5月)肥满度快速增加, 精卵集中排放的时期(7月, 10月)肥满度急剧下降。
3.3 红树蚬的胚胎发育特点
红树蚬的繁殖方式为非孵育型, 沉性卵, 卵被一层厚厚的胶质膜, 卵裂及早期胚胎发育在胶质膜内进行, 至担轮幼虫期开始以很快的速度撞击卵膜,冲破卵膜后孵化出来, 与大洋河河蚬比较相似, 但大洋河河蚬胚胎发育至D形幼虫阶段才孵出卵膜于水中浮游[10]。
[1]蔡英亚, 黄翔鹄, 吴洞科.红树蚬的生态观察[J].热带海洋, 1995, 14(1): 94-98.
[2]Morton B.The biology and functional morphology of the Southeast Asian mangrove bivalve,Polymesoda(Geloina)erosa(Solander1786) (Bivalvia: Corbiculidae)[J].Can JZool, 1976, 54: 482-500.
[3]Chatterji A, Ansari Z A, Ingole B S, et al.Indian marine bivalves: Potential source of antiviral drugs[J].Current Science, 2002, 82(10): 1279-1282.
[4]Morton B.The reproductive strategy of the mangrove bivalvePolymesoda(Geloina)erosa(Bivalvia: Corbiculidae) in Hong Kong [J].Malacological Review, 1985,18(1-2): 83-89.
[5]Gimin R, Luong-Van T, Mohan R et al.Aspects of the reproductive biology ofPolymesoda erosa(Solander,1786) (Bivalvia: Corbiculidae) in northern Australia [J].Beagle, 2005, 21: 37-46.
[6]Clemente S, Ingole B S.Gametogenic development and spawning of the mud clam,Polymesoda erosa(Solander,1876) at Chorao Island, Goa [J].Marine Biology Research, 2009, 5(2): 109-121.
[7]林志华, 单乐州, 柴雪良, 等.文蛤的性腺发育和生殖周期[J].水产学报, 2004, 28(5): 510-514.
[8]曾志南, 李复雪.青蛤的繁殖周期[J].热带海洋,1991, 10(1): 86-91.
[9]张俊杰, 周浩郎, 吴斌, 等.红树蚬表型性状对秋季软体部重的影响效果分析[J].广西科学, 2012, 19(4):384-386.
[10]王庆志, 常亚青.大洋河河蚬的繁殖生物学研究[J].大连水产学院学报, 2010, 2(1): 8-13.
