日本佳盾蜾蠃的生物学特性观察
2014-11-25黄敦元余江帆许晓风
黄敦元 ,何 波,谷 平,余江帆,彭 飞,许晓风
(1.江西环境工程职业学院,江西赣州 341000;2.南京师范大学生命科学学院动物学国家重点学科,江苏生物多样性重点实验室,南京 210046;3.江西省林业科技培训中心,南昌 330038)
蜾蠃亚科Eumeninae 是胡蜂科中最大的一个类群(Carpenter,1982;Kim et al.,2005),目前研究发现,全世界共有3585 种,我国现有记录161 种(Yamane 1990;Kim and Yamane,2001;周鑫等,2011,2012,2013;黄敦元等,2013;You et al.,2013)。蜾蠃亚科多为独栖物种,无固定居住巢穴,一般交配后开始营巢产卵繁育后代,其巢穴有两种类型:一种是呈中空壶状,上部有细颈,开口于颈端部的泥质巢穴 (李铁生,1982);另一种巢是利用开口于竹管、芦苇管等中空材料,衔泥分隔成多个虫室,直接将卵产于竹管或者苇管内壁上,产卵后外出捕捉蛾类幼虫带回虫室中以供其卵孵化出的幼虫食用,最后用泥块密封虫室和巢穴口 (Cowan,1991;Fateryga,2013)。
日本佳盾蜾蠃Euodynerus nipanicus (Schulthess)属膜翅目Hymenoptera 胡蜂科Vespidae 蜾蠃亚科Eumeninae 佳盾蜾蠃属Euodynerus Dalla Torre 1904(李铁生,1982),该蜂主要分布在中国的江苏、浙江、江西和台湾等地(黄敦元等,2013),以及日本和韩国等地(Kim,2012),是一种重要的独栖性天敌昆虫,其捕食凶猛,飞翔迅速,对林区鳞翅目蛾类害虫的控制作用明显 (李铁生,1985)。本研究利用人工巢管收集并室内观察该蜂,旨在进一步明确其生物学习性,以期为日本佳盾蜾蠃的保护与利用提供生物学依据。
1 材料与方法
1.1 研究地区概况
调查的时间从2011年2月到2013年11月。调查地点为江西省赣州市赣县沙地镇 (N:26°06';E:114°46')、江西省赣州市南康市谭东镇(N:25°76';E:114°87')和江西省赣州市湖边镇 (N:25° 53';E:114°54')共3个典型样地。
江西省赣州市气候特点:气候温暖、光照充足、雨量充沛、四季分明,春季天气多变且雨量充沛,回暖较早;夏季高温;秋季凉爽干燥;冬季阴雨天气较多,年平均气温在19.1℃至20.4℃,年平均无霜期287 天,年平均降雨量1605.6 mm,4-6月为雨季,占全年雨量的47%,年平均日照时数为1774.4 小时。
1.2 研究方法
1.2.1 幼虫的收集与观察
利用人工巢穴(trap-nesting) (图版Ⅰ-1)野外收集日本佳盾蜾蠃,实验室解剖观察并记录各虫态的发育历期;记录日本佳盾蜾蠃的生活史,同时对其寄生性天敌的寄生行为进行观察。用照相机(Canon Power Shot S3IS)拍摄各个虫态,自蛹期起每12 h 拍照一次,直至羽化,并用游标卡尺测量各虫态的长度大小。
1.2.2 羽化及性比的观察
野外每15 天采集一次筑巢巢管,解剖后统计羽化期每个巢穴中羽化出的雌雄个体数量,据此计算不同世代的性比。
1.2.3 成虫交配行为的观察
制作规格为3 m×3 m×3 m的网罩安置在蜂巢附近,将羽化出巢的雌雄个体人为放置在网罩内观察其交配行为。
1.2.4 成虫活动习性与寿命观察
用白色涂改液标记刚筑巢的巢管,对筑巢雌蜂的活动规律进行跟踪观察,同时通过野外观察和室内解剖雌蜂观察卵巢有无产卵斑相结合的方法(贺春玲等,2011)来确定成虫的寿命。
1.2.5 雌蜂访花行为观察
在日本佳盾蜾蠃成虫活跃期(2013年4、7、10月),每月随机选择4个晴好天气,巢区周边800 m 范围内观察该蜂到访的植物种类,辅以拍照和摄像的方法记录该蜂的访花行为。
1.3 数据统计与分析
实验数据存储和图形制作采用SPSS 13.0 和Excel 2003 软件进行处理。
2 结果与分析
2.1 形态特征(成虫)
日本佳盾蜾蠃雌性个体体长在9.68-12.35 mm 之间,平均11.17 ±0.25 mm (n=15)(图版Ⅰ-8)。头部宽略窄于胸部。额部除额沟处有1 黄斑外,全呈黑色,密布粗糙刻点,覆以浅色短毛。褐色单眼呈倒三角形排列于两复眼顶部之间,触角支角突黑色,柄节前缘黄色,余黑色,梗节及鞭节均黑色。颅顶部及颊部均黑色,仅复眼后缘上部有1 黄色斑,呈点状,刻点较浅,覆短毛。唇基半部黄色,端半部黑色,密布刻点及短毛,上唇棕色,上颚楔状,基部黑色,向端部渐呈暗棕色,近基部边缘有1 黄斑,齿尖锐。前胸背板前缘截状,肩角明显,前缘两侧各有1 黄色横带状斑,余均黑色,密布粗糙刻点,覆浅色短毛。并胸腹节全呈黑色,中部凹陷,有横皱褶,中央有纵沟,两侧刻点粗糙,覆短毛。中胸侧板全星黑色,中部有粗糙刻点,后胸侧板全呈黑色,布有横皱褶,均覆短毛。翅基片黄色,中央有1棕色斑,光滑无毛。腹部第1-4 节背板黑色,仅沿端部边缘有1 黄色横带,密布刻点及短毛。5-6 节背板及各节腹板全呈黑色,仅第2 节腹板两侧常有1 黄色斑,各节有略浅的刻点及短毛。
雄性个体体长在7.98-10.74 mm 之间,平均9.13 ±0.33 mm (n=15) (图版Ⅰ-8)。体色近似于雌蜂,唇基全呈黄色,中、后足基节前缘黄色,触角末端节呈钩状,腹部共7 节。
2.2 生物学特性
早在2009年8月,我们在江西宜春、赣州等地野外采集过程中发现有日本佳盾蜾蠃的分布,营巢地点多为枯萎的树枝和竹竿。为了便于研究该蜂的生物学及营巢生物学规律,我们尝试使用人工巢管(芦苇管)来收集该蜂,在2010年野外放置的巢管中发现该蜂有大量筑巢现象,2011年开始系统研究该蜂的生物学和营巢规律。日本佳盾蜾蠃的各虫态形态图版Ⅰ
2.2.1 生活史
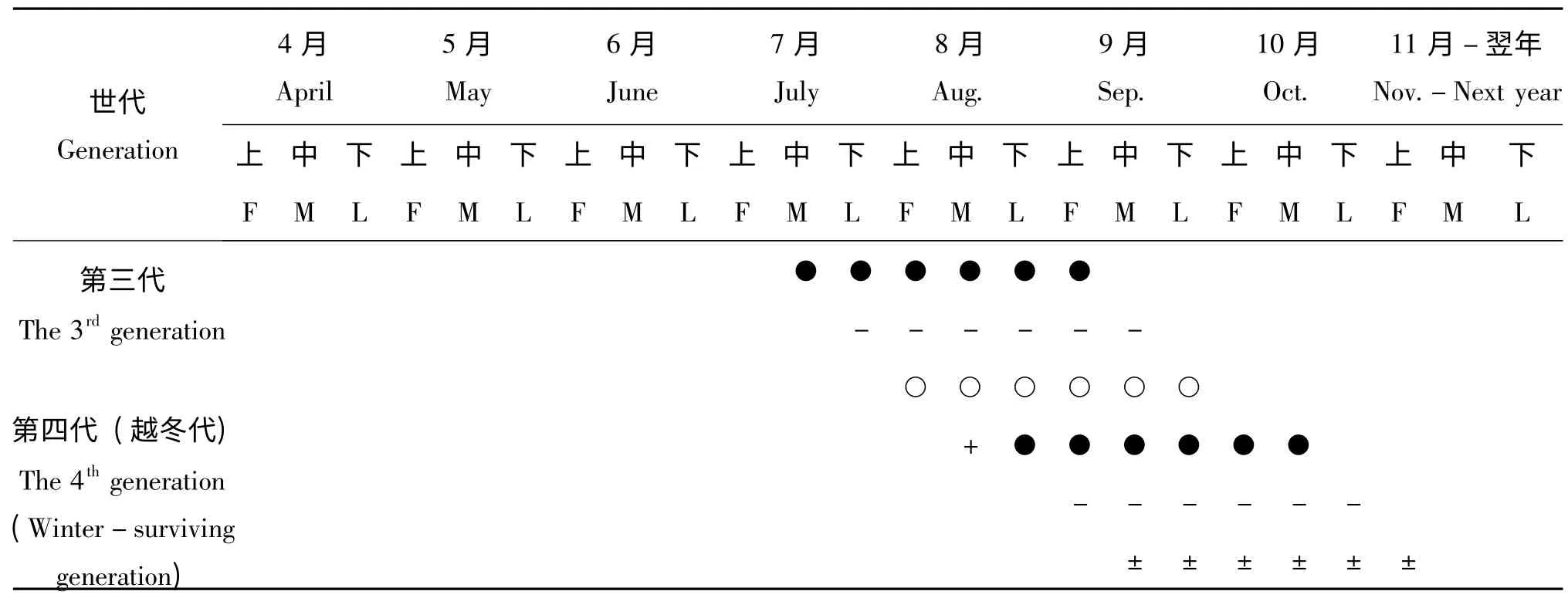
日本佳盾蜾蠃在江西赣州地区1年4 代,并有世代重叠现象,最后一代成虫10月下旬数量开始减少,11月上旬基本不见其活动,以滞育状态下的老熟幼虫在虫室内越冬,越冬成虫4月底开始羽化并陆续出巢活动,最后一代的成虫10月下旬数量开始减少,11月上旬基本不见其活动。自然条件下,越冬幼虫翌年4月中旬至5月初开始陆续化蛹,蛹期6-9 d。单个巢管中所有成虫的出巢历期约3-5 d,一般雄蜂先于雌蜂1-2 d 出巢,雌蜂羽化出巢后即可交配,交配后雌蜂多数在羽化地点附近选址筑巢,雌蜂寿命约25 d。雄蜂寿命约15 d,1 头雄蜂可与多头雌蜂交尾。该蜂卵期2-4 d,幼虫期6-10 d,幼虫取食完所在虫室中的食物后开始吐丝做茧,进入老熟幼虫期,约4-5 d,越冬代滞育期约190 d。日本佳盾蜾蠃在江西赣州地区的年生活史见表1。

表1 日本佳盾蜾蠃的生活史(江西,赣州)Table1 The life cycle of Euodynerus nipanicus (Schulthess)(Ganzhou,Jiangxi)
(续上表)
2.2.1.1 卵
卵近似微弯的长圆柱形(腊肠形),长度一般在2.74-3.06 mm 之间,平均2.91 ±0.13 mm (n=10),直径在0.78-0.92 mm 之间,平均0.84±0.04 mm (n=10),卵期2-4 d,平均2.5 ±0.99 d (n=10)。刚产的卵无色透明略带乳白色,表面光滑,24 h 后渐变成浅黄色(图版Ⅰ-2),以一根1-3 mm 长的白色丝状物悬挂在虫室最内端的上壁。大约1-2 d 后,卵的中部会有明显的膨大,2 d 后卵壳内幼虫的形态发育完成。体视镜(40×)下可观察到虫体在缓慢地蠕动,然后卵壳从背中线逐渐开裂,以头部破卵而出,掉落于食物上,幼虫不取食卵壳。
2.2.1.2 幼虫
初孵幼虫呈浅乳白色,体长一般在2.08-3.51 mm 之间,平均2.75 ±0.15 mm (n=10),无足,光滑无毛,体躯柔软弯曲呈“C”型,头部小,呈乳白色(图版Ⅰ-3)。幼虫吸附在虫室中的鳞翅目蛾类幼虫上,先用大颚咬开鳞翅目幼虫的虫体,通过身体的蠕动来吸食幼虫的体液至只剩躯壳。取食过程中能明显的观察到低龄幼虫体液的流动情况,食物在中肠后端开始积累,消化道的颜色也因此逐渐由后向前加深,逐渐能透过身体看到一条浅黄色的消化道。低龄幼虫(1-3 龄)只取食不排便,幼虫的体色与被取食幼虫的体色相似(图版Ⅰ-4)。4-5 龄幼虫节间多皱褶,粗肥,啃食鳞翅目幼虫的躯体,此时幼虫体色均转为浅绿或黄白色(图版Ⅰ-5),取食量明显增大。5-7 天幼虫进入吐丝阶段,茧的大小一般与虫室的大小一致,也可能略小于虫室。幼虫做完茧后,进行一次排便,排泄物为棕黑色粘稠。之后体色渐为浅黄色,体形由“C”型转为扁平且直,进入老熟期(图版Ⅰ-6),幼虫期6-10 d,平均8.1±0.99 d (n=10)。
老熟幼虫体粗胖、两端略尖呈梭形,体表光滑无毛,体节明显可见且呈深黄色(图版Ⅰ-7),体长1.209 ±0.261 mm,宽0.412 ±0.098 mm。头部宽圆,骨化弱,上颚具简单齿,下颚须及下唇须明显可见。日本佳盾蜾蠃老熟幼虫期一般为4-5 d (越冬代老熟幼虫期及滞育期较长,约190 d)(图版Ⅰ-7)。
2.2.1.3 蛹
日本佳盾蜾蠃的蛹型为被蛹(图版Ⅰ-10)。雌雄个体蛹的大小差异较大,雌蛹体长在10.64-13.92 mm 之间,平均12.31 ±1.08 mm (n=15);雄蛹体长在8.84-10.82 mm 之间,平均10.07 ±0.56 mm (n=15)。体色初期乳白色,逐渐由乳白色-淡黄色-土黄色-褐色-黑色转变(图版Ⅰ-10);头胸部的颜色先于腹部变暗;腹部节间处随头胸部同时变黑,其余部位由褐色逐渐至黑色。复眼1 对,单眼3 只,位于额顶两复眼中间,呈倒三角形排列;初期单、复眼颜色与虫体相近,约2 d 后变为浅粉色,并逐渐由浅红色转红色,再由红色转暗红色后转黑色。整个蛹期约6-9 d,平均7.4 ±0.84 d (n=10),羽化完成后变成成虫,新成虫打通虫室隔离层及前厅堵塞物,爬出巢口。

图版Ⅰ 日本佳盾蜾蠃的形态特征PlateⅠ The morphology of Euodynerus nipanicus
2.2.2 成虫的羽化及性比
利用人工巢管筑巢的日本佳盾蜾蠃在江西赣州地区每年4月底至5月初开始陆续羽化出巢,雄蜂先于雌蜂1-2 天羽化出巢,出巢后在巢区附近抖动双翅清理虫体等待其它雌蜂羽化出巢。越冬代成虫从4月底开始羽化,5月初羽化数量逐渐上升并达到高峰,至5月中旬开始数量逐渐降低,其他世代的羽化时间见表1。成蜂羽化时间主要集中在晴好天气下的上午7∶00-9∶00,其次是下午15∶00-17∶00 时。
日本佳盾蜾蠃不同世代的性比略有不同,全年共收集巢管200 枝,累计羽化出巢1020 头,其中雌蜂762 头,雄蜂258 头。通过表2 可以看出,不同时期收集的巢管,其出蜂的雌雄比有一定的差异,其中5月15 收集的巢管,其出蜂的雌雄比最大,为3.67∶1;雌雄比最小的是在8月15 日收集的巢管,仅为2.31∶1;10 次收集的巢管成虫羽化后的平均性比为2.95∶1。
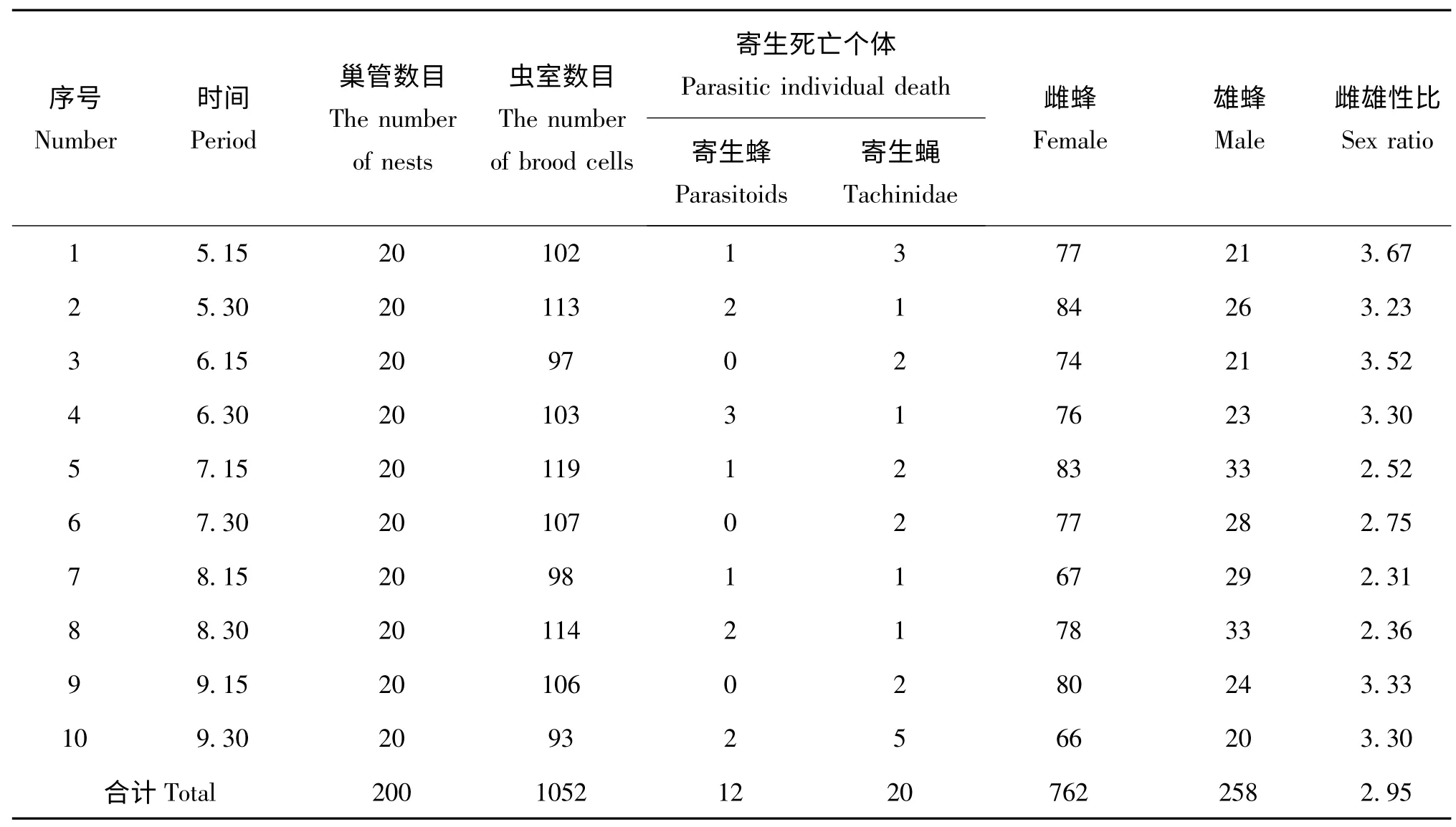
表2 日本佳盾蜾蠃的性比及寄生情况(江西,赣州)Table 2 The sex ratio and parasitism of Euodynerus nipanicus (Schulthess)(Ganzhou,Jiangxi)
2.2.3 成虫的交配行为
成虫在羽化出巢当天即可进行交配,交配率以羽化后次日上午最高,4 d 后显著下降。晴好天气下,该蜂全天均可进行交配,交配高峰一般在上午的8∶00-10∶00 之间和下午的16∶00-18∶00之间。雄蜂羽化较雌蜂早且主要集中在巢口附近的杂草灌木周围或是蜜源植物附近等待羽化出巢的雌蜂,寻找交配机会,一旦发现雌蜂,雄蜂立即扑上去与之交尾。交尾时雄蜂爬到雌蜂体背用上颚和足抓住雌蜂,不断用前足拨动雌蜂的前胸背部,待雌蜂腹部末端上翘接受交尾时,雄蜂伸长并向下弯曲腹部,翻出阳茎,插入雌蜂的生殖孔。交配时间在60-250 s 之间,平均141.1 ±46.2 s (n=10)。一头雄蜂可先后可与多头雌蜂交配,而雌蜂一生只接受一次交配。
2.2.4 成虫的营巢活动
雌蜂羽化出巢后寻找蜜源植物补充营养,期间完成交配。交配后的雌蜂在羽化巢区附近选择合适的巢管并清理好巢管后,开始采集湿润的泥土筑巢。筑巢时从巢管的末端开始,一般按照构筑末端预封塞物(deep plug)、构建虫室(brood cell)、产卵、准备蜂粮、构建室间隔离层 (cell partition)、构建空室 (deep cell)构建下一个虫室、最后用泥块密封管口意味前庭 (vestibular cell)构筑完成。雌蜂在晴好天气下每天出巢次数在45-66 之间,平均55 ±5.33 次(n=10),其中捕食次数约26 次,采集泥块次数约21 次,其他为外出采集花蜜补充自身营养或是采集不成功。雌蜂晚上栖息于距巢口1-2 cm 主道内。
雄蜂羽化出巢后寻找蜜源植物补充营养,白天潜伏在巢口附近的杂草灌木周围或是蜜源植物附近寻求交配机会,不参与新巢的构建和蜂粮的采集。在同一个巢管雌蜂没有羽化之前,雄蜂夜宿巢管并帮助打通虫室通向巢口的隔离层;在雌蜂筑巢期间,雄蜂一般不夜宿自己羽化的巢管和雌蜂新构建的巢管,而是寻找废弃的老巢或是杂草灌木的叶面背部夜宿。雄蜂寿命短于雌蜂,一般只有15 d 左右。
2.2.5 蜜源植物
日本佳盾蜾蠃为捕食性天敌昆虫,雌蜂捕食鳞翅目昆虫幼虫作为繁育后代的食物。同时,该蜂也为杂食性昆虫,观察表明,该蜂访问紫藤Wisteria sinensis Sweet、黄 荆Vitex negundo Linn.、红蓼Polygonum orientale Linn.、野棉花Anemone hupehensis Lemoine.(黄敦元等,2013)、盐肤木Rhus chinensis Mill.、空心菜 Ipomoea aquatica Forsk、野棉花Anemone hupehensis Lemoine.等植物的花朵,取食花蜜补充自身营养。
日本佳盾蜾蠃雌性个体的额、颅顶、胸部等部位覆有短毛,其访花主要目的是取食花蜜维持个体生存需要,在访问蜜源植物花时,前足、中足攀附在花萼或花枝上,将头伸入花柱底部取食花蜜,在取食花蜜过程中常会对花部结构造成一定的破坏,但同时体壁上会粘附少量花粉,对到访植物可能具有一定的传粉功能。
2.2.6 寄生性天敌
通过对定期收集回实验室的巢管进行室内饲养和观察,在200个巢管中,累计有1052个虫室,每个虫室有1 头日本佳盾蜾蠃幼虫,因寄生而死亡的幼虫有32 头,其中有12 头幼虫被寄生蜂寄生,20 头幼虫被寄生蝇寄生(表2)。本研究中涉及的寄生蜂和寄生蝇具体物种需要进一步鉴定。
3 结论与讨论
3.1 雌雄比在不同世代之间的差异
巢管营巢的独栖野生蜜蜂和胡蜂类群,其不同世代雌雄比的差异主要受以下因素的影响:首先是巢区附近食物资源的丰富度是影响雌雄比的主要因素之一 (Jason et al.,2006;Peterson and Roitberg,2006),Jason 等 (2006)研究显示:Megachile rotundata (F.)巢区附近食物资源的丰富度通过影响其后代个体幼虫期的食物多少而决定其后代的雌雄比;其次是受到雌蜂所选择巢管的规格及制作虫室的大小影响 (Carlo et al.,2011;Fateryga,2013),Fateryga (2013)通过野外观察和解剖Euodynerus quadrifasciatus (F.)的巢管提出:巢管中单个虫室大小是影响不同虫室中卵和幼虫的发育情况进而影响到个体大小和雌雄性比,因为虫室的大小制约虫室中幼虫发育所需的食物数量;再次是季节的更替和温度的变化也是导致雌雄个体差异及性比不同的因素之一(Fye,1965;Longair,1981),Fye (1965)通过巢管诱集大量Rygchium leucomelas (Sauss.)并对其生物学进行研究发现,该蜂一年2 代,越冬代雄性个体的数目偏多,夏季一代的个体雌性个体偏多。本研究中的日本佳盾蜾蠃在每年的第1 代和最后1 代的雌雄比比较大,7-8月份羽化出巢的第3 代雌雄比明显偏小,我们初步认为该时期的食物资源紧缺和巢区内的温度过高是导致这种现象的主要因素,有待于进一步的验证。
3.2 巢室内成蜂羽化顺序
雌蜂在营巢构建虫室时,发育成雄性成虫的虫室一般在最外层的几个虫室中。雄蜂先于雌蜂1-2 d 羽化,羽化后雄蜂打通虫室通向巢口的室间隔离层外出取食,雌蜂没有羽化之前回巢管过夜,等待后续羽化的雌蜂并帮助及时打通巢管末端的室间隔离层。雄蜂是如何确定雌蜂的羽化我们目前还尚不清楚,初步推测是雄蜂听到巢管虫室内雌蜂羽化后翅膀产生的振动以便确定雌蜂的羽化。
References)
Bruun HH.Patterns of species richness in dry grassland pat ches in an agricultural landscape[J].Ecography,2000,23,641-650.
Carlo P,Roberto B,Walter B.Few,small,and male:multiple effects of reduced nest space on the offspring of the solitary wasp,Euodynerus (Pareuodynerus)posticus (Hymenoptera:Vespidae)[J].Comptes Rendus Biologies,2011,334 (1):50-60.
Carpenter JM.The phylogenetic relationships and natural classification of the Vespoidea (Hymenoptera)[J].Systematic Entomology,1982,7 (1):11-38.
Cowan DP.The solitary and presocial Vespidae.In Ross KC,Matthews RW,eds.The Social Biology of Wasps[C].Ithaca and London:Comstock Publishing Associates,1991,678.
Fateryga AV.The nest structure in four wasp species of the genus Euodynerus Dalla Torre (Hymenoptera,Vespidae:Eumeninae)[J].Entomological Review,2013,99 (4):412-421.
Fye RE.The biology of the Vespidae,Pompilidae and Sphecidae(Hymenoptera)from trap nests in northwestern Ontario [J].Canadian Entomol.,1965,97 (7):716-744.
He CL,Ji BZ,Liu SW.Morphology and biology of Xylocopa tranquebarorum[J].Chinese Journal of Applied Entomology,2011,48 (6):1751-1758.[贺春玲,嵇保中,刘曙雯.长木蜂的形态和生物学观察[J].应用昆虫学报,2011,48 (6):1751-1758]
Huang DY,He B,Yu JF,et al.Nesting biology of Euodynerus nipanicus Schulthess (Hymenoptera:Vespidae)[J].Journal of Environmental Entomology,2013,(35)6:778-787.[黄敦元,何 波,余江帆,等.日本佳盾蜾蠃营巢生物学研究[J].环境昆虫学报,2013,35 (6):778-787]
Jason HP,Bernard DR,Peterson JH.Impacts of flight distance on sex ratio and resource allocation to offspring in the leafcutter bee,Megachile rotundata [J].Behavioral Ecology and Sociobiology,2006,59 (5):589-596.
Ju Y,Chen B,Li TJ.Two new species of the genus Ancistrocerus Wesmael (Hymenoptera:Vespidae:Eumeninae) from China,with a key to the Oriental species[J].Zookeys,2013,303:77-86.
Kim JG,Choi YC,Choi JY,et al.Environmental evaluation by using Hymenoptera induced by Bamboo Pipe Traps indicated by Eumenid wasp (Hymenoptera:Eumenidae)[J].Korean Journal of Applied Entomology,2005,44:307-315.
Kim JK,Yamane S.A revision of Eumenes Latreille (Hymenoptera:Vespidae)from the Far East Asia,with descriptions of one new species and one new subspecies[J].Entomological Science,2001,4 (2):139-155.
Kim JK.Taxonomic Review of the genus Euodynerus (Hymenoptera:Vespidae:Eumeninae)in the Korean Peninsula[J].Anim.Syst.Evol.Divers.,2012,28 (3):161-167.
Peterson JH,Roitberg BD.Impact of resource levels on sex ratio and resource allocation in the solitary bee,Megachile rotundata [J].Environmental Entomology,2006,35 (5):1404-1410.
Lee TS.Wasps in Rural Region of China[M].Beijing:Agricultural Publishing House,1982,64-115.[李铁生.中国农区胡蜂.北京:农业出版社.1982,64-115]
Lee TS.Economic Insect Fauna of China,Fasc.30,Hymenoptera:Vespoidea[M].Beijing:Science Press,1985,1-159.[李铁生.中国经济昆虫志·第三十册· 胡蜂总科[M].北京:科学出版社,1985,1-159]
Longair RW.Sex-ratio variations in xylophilous aculeate Hymenoptera[J].Evolution,1981,35:597-600.
Zhou X,Chen B,Li TJ.Two new species of the genus Discoelius Latreille(Hymenoptera:Vespidae:Eumeninae)from China,with a key to the Chinese species[J].Journal of Hymenoptera Research,2013,32:45-54.
Yamane S.A revision of the Japanese Eumenidae (Hymenoptera,Vespoidea)[J].Insecta Matsumurana,New Series,1990,43:1-189.
Zhou X,Chen B,Li TJ.Two new species and a key to species of the genus Eumenes Latreille (Hymenoptera:Vespidae:Eumeninae)from southwestern China[J].Entomotaxonomia,2012,34 (2):467-474.
Zhou X,Li TJ,Chen B.The Taxonomic research progress of Eumeninae(Hymenoptera:Vespidae)[J].Journal of Chongqing Normal University (Natural Science),2011,28 (6):22-29.[周鑫,李廷景,陈斌.蜾蠃亚科(膜翅目:胡蜂科)分类研究进展[J].重庆师范大学学报(自然科学版),2011,28 (6):22-29]
