光照增强对林下云锦杜鹃树苗光合作用的影响
2014-11-24柯世省张云生
柯世省,张云生
(1.台州学院生命科学学院,浙江 台州 318002;2.浙江省台州市椒江区林业特产总站,浙江 台州 318000)
云锦杜鹃(Rhododendron fortunei)为杜鹃花科(Ericaceae)杜鹃花属常绿灌木或小乔木,是常绿杜鹃亚属中最原始的一个类群[1],自然分布于长江流域各省,生于海拔400 ~ 1 900 m的沟谷阔叶林或山顶灌草丛,生境阴凉潮湿,雨水充沛。云锦杜鹃为常绿树种,叶片寿命2 ~ 3 a,每年3月底新叶开始长出,枝条和叶片生长迅速,5月中上旬开花,花期约半个月,6月中上旬叶片基本成熟,新叶不再长出。云锦杜鹃适宜作观花或观叶植物,是一种观赏价值高、开发潜力大的园林树种[2]。在自然条件下,云锦杜鹃在林窗、林缘和林下均可生长,但林下生长缓慢,开花数目极少。云锦杜鹃种子掉落后萌发成为幼苗,绝大部分的幼苗见于林下。
在良好的森林生态系统中,林下的光照水平往往不足冠层的 5%,成为植物生长的严重制约因素。由于森林生境片段化、冠层树木倒伏或断枝等原因形成新的林窗,使得原林下的光照突然和剧烈地增强,而原林下生长的植物能否适应这样的变化将成为影响植株生长的关键因素。当植物生长环境从低光强转变为高光强时,正在展开或新长出的叶片可能会适应高光强,但在林下已发育成熟的叶片则往往会遭受一段时间的光抑制,导致光合速率的降低和光合器官的损伤[3]。随着强光照射时间的延长(两周或更久),不同种类植物在林下发育成熟的叶片的光合作用光抑制的持续时间差异甚大,有些种类会遭受长期的光抑制,而有些种类的光合速率会回复到初始值或者更高,也有一些植物在强光照射下甚至不会出现明显的光抑制[4]。对于具有明显光抑制的植物而言,耐荫植物往往比通常生长在高光环境中的植物更容易发生光抑制。因此,林下植物的种类和其叶片的发育程度决定着光抑制的严重程度并成为该植物能否适应光照增强这种新生长环境的重要限制因素。本文以林下生长的云锦杜鹃树苗为材料,模拟林窗和林缘生境,研究不同光照强度对其成熟叶片光合作用的影响,并与长期生长在这三种光环境下的云锦杜鹃进行比较,探讨其光适应能力和机制,也为园林建设中云锦杜鹃树苗的移栽提供理论依据。
1 材料和方法
1.1 样地选择和材料处理
研究在浙江省天台县华顶国家森林公园云锦杜鹃林进行。海拔约900 m,属亚热带季风性湿润气候,年降水量1 700 mm,年平均相对湿度85%以上,年平均气温13℃,无霜期约230 d。山地土壤系水成岩及火成的花岗岩母质上发育的黄壤土,土层厚30 ~ 100 cm,尚属湿润肥沃。与云锦杜鹃伴生的主要种类有金钟花(Forsythia viridissima)、灰白蜡瓣花(Corylopsis glanduliferavar.hypoglauca)、绿叶甘橿(Lindera fruticosavar.fruticosa)、半边月(Weigela japonicavar.sinica)、白檀(Symplocos paniculata)、山橿(Lindera reflexa)、黄山松(Pinus taiwanensis)、柳杉(Cryptomeria fortunei)、檫木(Sassafras tzumu)等。2012年3月上旬,即云锦杜鹃新叶未长出前,在云锦杜鹃林林下挖取4 ~ 5年生云锦杜鹃树苗(约50 cm高)移栽到塑料花盆中,一盆一株,共50盆。各将10盆放置于云锦杜鹃林的林窗和林缘生境中,另30盆置于林下。林窗生境能接受到约7 h/d的阳光直接照射(高光强,H),林缘生境在中午前阳光直接照射约4 h(中光强,M),林下生境的光强约为林窗的5%(低光强,L)。当年7月初,当年生叶片成熟后,分别将置于林下的树苗各10盆转移到林窗(L-H)和林缘(L-M)生境中,留下10盆仍放置于林下(L)作为对照。
1.2 光合作用光响应测试
于植株转移前和转移后每隔5 d对各生境植株当年生成熟叶片进行持续30 d的光响应测试,测试在控制条件下进行。用Li-6400XT型光合测定仪(Li-Cor,Lincoln,Nebraska,USA)控制叶室温度为25℃,进气CO2浓度为350 μmol/mol,由仪器配备的LED提供光源,测试光强分别为2 000、1 500、1 200、1 000、800、600、400、200、150、100、50、20和0 μmol·m-2·s-1。测得从低到高一系列光合有效辐射下叶片的净光合速率。利用光合助手软件(LI-COR Application Note #105)拟合光响应曲线,求得表观量子效率(AQY)、光补偿点(LCP)、光饱和点(LSP)、暗呼吸速率(Rd)和最大净光合速率(Pnmax)。
1.3 叶绿素荧光参数测定
凌晨,用OS30P型叶绿素荧光仪(OPTI-sciences,USA)采用快速动力学法测定最大光化学效率(Fv/Fm),所测叶片与测定光合作用光响应的相同,测定前将叶片夹入暗适应夹2 h。
转移后第30 d,按Pastenes等[5]的方法用FMS-2便携式调制荧光仪(Hanstech, UK)测定叶绿素荧光参数,计算实际光化学效率(PhiPSⅡ)、光化学淬灭系数(qP)和非光化学淬灭系数(qN),所测叶片与测定光合作用光响应的相同。
1.4 其他参数测定
光合作用测定结束后,采集样品叶带回实验室。用WinFOLIA系统(Regent,Canada)测定叶面积,在60℃烘箱中干燥48 h后称重,计算比叶重(LMA)。干燥样品粉碎后用微量凯氏定氮法测定叶片单位重量氮含量,根据叶片的LMA换算为单位面积氮含量(Narea)。叶绿素但用丙酮提取,比色法测定[6]。
1.4 数据的统计分析
试验设5个重复,用DPS软件对实验结果进行方差分析,显著性差异运用Duncan’s检验法进行多重比较。
2 结果与分析
2.1 光合作用光响应参数的动态变化
通过光合速率的光响应测试,可以计算出Pnmax、Rd、LCP、LSP和AQY等光合参数。不同光环境下生长的云锦杜鹃树苗光合作用的光响应参数如图1所示。一直分别生长在三种光环境下的云锦杜鹃树苗在30 d的实验期间没有新叶长出,也没有叶片掉落,它们的Pnmax在实验前后变化不大,但三者之间的Pnmax差异显著。林下云锦杜鹃(以下称L)的Pnmax最低,林缘云锦杜鹃(M)的Pnmax最高,是L的2.4倍,而林窗云锦杜鹃(H)Pnmax显著降低,只是M的77%,表明云锦杜鹃虽然在三种光环境下都能生长,但过强(全光照)或过弱的光环境都不利于其光合作用的进行。将林下云锦杜鹃转移到林窗(L-H)和林缘(L-M)生境后,它们Pnmax的动态变化发生了明显差异。转移后10 d内,L-H的Pnmax降至最低,以后逐渐恢复并达到稳定水平,但仍显著低于转移前水平(即L的Pnmax)。L-M在转移后5 d其Pnmax就有所增加,在以后的15 d内增加较为迅速并接近H的Pnmax,接着继续慢速增加,转移后30 d达到L-H的2.6倍,但仍显著低于M(图1)。三种光环境下,L的Rd最低,H最高,是L的1.8倍,M降低幅度较大,仅为L的1.2倍。林下的云锦杜鹃在转移后,其Rd即逐渐增高,L-M增加较为平缓,L-H的增加则较为迅速,至实验结束时,L-M的Rd达到M水平,L-H则是M的1.26倍,是L的1.43倍,但显著低于H。M的LCP虽然高于L,但差异并不显著,H的LCP是M的1.86倍。林下云锦杜鹃转移至另二种光环境后(L-M,L-H)其LCP均增加,转移后5 d即基本上已接近相应环境中的对照植株。三种光环境中云锦杜鹃的LSP差异显著,L最低,H最高,H是L的1.74倍,M是L的1.33倍。林下云锦杜鹃转移后,L-M的LSP变化较为平缓,稍高于L值;L-H的LSP变化幅度较大,转移后前15 d均低于L值,后逐渐增加,至实验结束时接近M值。M的AQY稍高于L,但差异不显著,两者均显著高于H。林下云锦杜鹃在转移后第5 d其AQY即显著降低,L-M的AQY略低于H,而L-H的AQY仅为H的68.9%;第10 d后,L-M和L-H的AQY有所恢复并维持稳定,至实验结束时,L-M的AQY达到L和M的水平,但L-H的AQY仍显著低于H,是H的85.5%,是L-M的74.4%。

图1 云锦杜鹃树苗Pnmax、Rd、LCP、LSP和AQY的动态变化Figure1 Dynamics of Pnmax, Rd, LCP, LSP and AQYin the leaves of R.fortuneisaplings
2.2 叶绿素荧光参数
2.2.1 光系统Ⅱ最大量子效率的动态变化 暗适应后三种光环境下的云锦杜鹃(L,M,H)的最大光化学效率(Fv/Fm)差异不显著,尽管 H相对较低(图2)。从图2还可以看出,林下的云锦杜鹃在转移后其Fv/Fm显著降低,L-H降低幅度大于L-M,两者在第5 d后开始恢复,至实验结束时,仍显著低于相应光环境中的对照植株,L-M的Fv/Fm是M的89.1%,L-H的Fv/Fm是H的68.2%。

图2 云锦杜鹃树苗光系统Ⅱ最大光化学效率的动态变化Figure2 Dynamic of maximum photochemical efficiency of
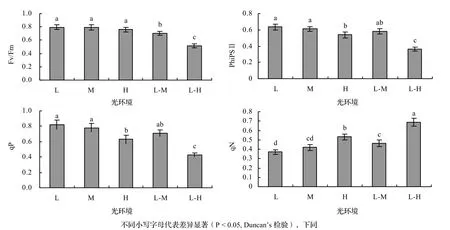
图3 云锦杜鹃树苗最大量子效率、实际量子效率、光化学淬灭系数和非光化学淬灭系数Figure3 ComparisonsofFv/Fm,PhiPSII,qPandqNintheleavesof R.fortunei saplings
2.2.2 不同光环境下叶绿素荧光参数的比较 至实验结束时,各种光环境中云锦杜鹃(L,M,H,L-M,L-H)的叶绿素荧光参数如图3所示。L、M和H具有较高和相似的Fv/Fm,L-M和L-H显著降低,L-M是L的88.3%,L-H是L的65.1%。L和M具有较高和相似的PhiPSⅡ,显著高于H,L-M的PhiPSⅡ低于L和M,高于H,但差异均不显著,而L-H的PhiPSⅡ最低,仅为L-M的62.3%和H的67.7%。各种光环境中云锦杜鹃的qP与PhiPSⅡ的变化趋势相似。L-H的qN显著高于其他光环境中的云锦杜鹃,L、M和L-M的qN则相对较低。

图4 云锦杜鹃树苗叶片比叶重、叶氮含量、叶绿素含量和叶绿素a/b值Fig ure 4 Comparisons of LMA, Narea, Chlmass, Chlareaand ratio of Chl a/b in the leaves ofR.f ortunei saplings
2.3 叶片的其他性质
从图4可以看出,实验结束时,M和H具有相似的单位面积叶重量(LMA),并显著高于其他三种类型,后三者的LMA差异不显著。从叶氮含量(Narea)来看,M最高,L最低;L-M的Narea显著高于L,但显著低于M;而L-H的Narea则显著低于H,与L差异不大。随着环境光强的增加(L,M,H),云锦杜鹃以叶片重量计量的叶绿素含量(Chlmass)显著降低;L-M的Chlmass低于L,高于M,但差异不显著;L-H的Chlmass显著低于H,为各生境中的最低值。与Chlmass不同,以叶片面积计量的叶绿素含量(Chlarea)M最高,其次为L,并显著高于其他三种类型;L-M的Chlarea显著低于M,与L差异不显著;L-H的Chlarea最低。随着环境光强的增加(L,M,H),云锦杜鹃叶绿素a/b值逐渐增高,H显著高于L;在转移实验过程中(L-M,L-H),林下发育成熟的云锦杜鹃叶片的叶绿素a/b值变化不大,与L差异不显著。
3 讨论
光照强度是影响植物光合作用、生长和繁殖的主要环境因素之一。本研究结果表明,不同的光环境(林下、林缘、林窗)明显影响云锦杜鹃树苗的光合作用,导致光合能力的显著差异,其中林缘生境中最高。这与对一些植物的研究结果相同[7],但也有一些植物在全光照条件下其光合能力最强[8]。本研究结果可能与云锦杜鹃在高光强下产生轻度的光抑制有关。将叶片已在林下发育成熟的云锦杜鹃树苗分别转移到林缘和林窗生境,其光合作用发生了明显分化。在实验持续期间,转移到林缘生境的云锦杜鹃的 Pnmax显著增加,并最终超过了林窗生境的对照植株,而转移到林窗生境的云锦杜鹃的 Pnmax则显著降低,最终也没有恢复到原来的水平,这表明突然增强的光照能否促进成熟阴生叶的光合作用取决于叶片所接受到的光的强度。从本文的结果来看,林缘的光环境有利于云锦杜鹃成熟阴生叶光合水平的提高,但林窗的光环境则严重抑制了光合作用的进行。
AQY和Fv/Fm的明显降低是光抑制发生的可靠判据[9]。林窗对照植株的AQY和PhiPSⅡ明显低于其他两个生境的结果表明,在全光照条件下,云锦杜鹃阳生叶的光合作用已受到了光抑制的影响,尽管Fv/Fm降低不明显,光抑制是林窗云锦杜鹃Pnmax低于林缘的一个重要原因。当林下云锦杜鹃接受的光照突然增加时(L-M,L-H),AQY和Fv/Fm立即降低随后不同程度地逐渐恢复,表明发生了光抑制和光抑制不同程度地解除。一方面,光抑制的程度取决于叶片接受的光照强度。另一方面,在强光下是否易于产生光抑制又决定着植物对生长环境光强的耐受性。同时,植物转移先后所在生长环境光强的变化情况也是决定植物光抑制程度的重要因素,因为光能的过剩会有很大的差异。林下云锦杜鹃转移到林窗后(L-H),光饱和点降低,虽然后来有所增加,但远低于H,光能严重过剩,其平均AQY和Fv/Fm不仅明显低于L-M,还低于H,表明L-H的光抑制比其他植株更严重,导致其Pnmax低下,生长状况恶化,难以适应林窗的高光强环境。林下云锦杜鹃转移到林缘后(L-M),其Pnmax即迅速增加,其AQY、PhiPSⅡ和Fv/Fm接近正常值,表明L-M基本上没有发生光抑制。植物叶片的光合特性对光环境具有明显的适应性,与阴生叶相比,通常阳生叶的叶片较厚,以容纳更多的叶绿体,并表现出较高的光合能力,但高光合能力并不总是伴随着较厚的叶片。叶片厚度主要取决于叶片发育时的光照强度,一旦叶片已成熟,其厚度就基本上不会发生大的变化[10]。当阴生叶成熟后转移至合适光强的环境中后,其光合能力通常会增加,但叶片厚度的变化则很小。Oguchi等[11]的相关研究表明,当阴生成熟叶转移到光线较强的生境时,其叶片厚度决定着 Pnmax的上限,因为没有足够的空间容纳伸长的叶绿体,尽管在这样的生境中叶片的LMA和厚度会稍有增加。本文的研究结果也表明,L-H和L-M的LMA虽然比L有所增加,但差异并不显著,L-M的Pnmax比L有了显著提升,但仍然显著低于M。叶片氮含量取决于其生长环境的光强,叶片中1,5-二磷酸核酮糖羧化酶(Rubisco)和光系统Ⅱ的数量随着生长环境光强的增加而增加,并与叶氮含量和Pnmax有密切的正相关[12]。L-M云锦杜鹃的叶氮含量比L有了显著增加,使得L-M叶片的Rubiso和光系统Ⅱ的数量得以增加,光合能力增强,较高的氮含量似乎是维持叶片较高的Pnmax的重要因素。叶绿素a/b值的变化与两个光系统的光吸收能力的平衡有关,通常情况下,阳生叶的叶绿素含量较低,但叶绿素a/b值较高,增加叶绿素a/b值可减少光系统Ⅱ集光天线色素的数量,以减少光能的过剩并减轻光抑制程度[13]。本文的结果与上述观点一致,随着生长环境光强的增加,云锦杜鹃(L,M,H)叶绿素含量显著降低;转移后,L-M和L-H的叶绿素含量也显著降低,但L-H降低的程度更大。生长在不同光环境下云锦杜鹃叶片叶绿素a/b值的变化可能受到光的量和质的共同调节,这样的变化有助于纠正两个光系统在不同光条件下对光能吸收的失衡,既可以提高光合效率,又可以减轻光损伤。叶片吸收的光能可通过光化学和非光化学途经耗散。随着光照强度的增加,过剩光能的比率也随之增加,而在强光条件下过剩的光能就会引起光抑制和光损伤。不同光环境中云锦杜鹃的光饱和点尽管差异显著,但最高也不超过 800 μmol·m-2·s-1(H),但林窗生境中午的光照强度可达 1 800 μmol·m-2·s-1,远远超过云锦杜鹃的光饱和点。所以,林窗云锦杜鹃较低的光合能力、AQY和PhiPSⅡ应归因于光抑制的发生和光系统Ⅱ的部分关闭。通过叶黄素循环进行的非光化学淬灭是植物应付过剩光能最常用的方式。林窗生境云锦杜鹃(H,L-H)的非光化学淬灭系数(qN)显著高于其他植株(L,M,L-M),并分别达到了 0.53(H)和0.69(L-H),而对许多植物而言,0.5左右的qN是判断是否发生了光抑制的阈值[5]。
4 结论
生长在不同光环境下云锦杜鹃的光合特性反映了其对光照强度变化的生理适应性,光合作用对不同光强的适应状况与LMA、叶氮含量、叶绿素含量及叶绿素a/b值、热耗散方式等有关,云锦杜鹃光合作用对较大范围光照条件的成功适应是其地理分布较广的重要原因,但林下生长的云锦杜鹃对光照突然增强的适应能力较弱。为了使林下云锦杜鹃能最终成功地适应强烈的光照水平(如全光照),减轻光抑制,加速其生长,应逐渐增加其生境的光强。本文的研究结果为野外云锦杜鹃的管理策略(伐疏、整枝等)和云锦杜鹃树苗的园林化移栽提供有用的指导。
[1]方瑞征,闵天禄.杜鹃属植物区系研究[J].云南植物研究,1995,17(4):359-379.
[2]边才苗,金则新.天台山云锦杜鹃的开花与结实特性[J].园艺学报,2006,33(1):101-104.
[3]Coopman R E, Reyes-Díaz M, Briceño V F,et al.Changes during early development in photosynthetic light acclimation capacity explain the shade to sun transition in Nothofagus nitida [J].Tree Physiol, 2008(28):1 561-1 571.
[4]Pandey S, Kushwaha R.Leaf anatomy and photosynthetic acclimation inValeriana jatamansiL.grown under high and low irradiance [J].Photosynthetica, 2005(43):85-90.
[5]Pastenes C, Santa-Maria E, Infante R,et al.Domestication of the Chilean guava (Ugni molinae Turcz.), a forest understorey shrub, must consider light intensity[J].Sci Hor, 2003(98):71-84.
[6]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[7]Zhang S R, Ma K P, Chen L Z.Response of photosynthetic plasticity of Paeonia suffruticosa to changed light environments[J].Environ Exp Bot,2003(49):121-33.
[8]Coopman R E, Fuentes-Neira F P, Briceño V F,et al.Light energy partitioning in photosystems I and II during development of Nothofagus nitida growing under different light environments in the Chilean evergreen temperate rain forest[J].Trees, 2010(24):247-259.
[9]Niyogi K K, Li X P, Rosenberg V,et al.Is PsbS the site of non-photochemical quenching in photosynthesis? [J].J Exp Bot, 2005(56):375-382.
[10]Oguchi R, Hikosaka K, Hirose T.Does the photosynthetic light-acclimation need changes in leaf anatomy? [J].Plant Cell Environ, 2003(26):505-512.
[11]Warren C R, Adams M A.Distribution of N, Rubisco and photosynthesis in Pinus pinaster and acclimation to light[J].Plant Cell Environ, 2001(24):597-609.
[12]Evans J R, Poorter H.Photosynthetic acclimation of plants to growth irradiance: the relative importance of specific leaf area and nitrogen in maximizing carbon gain[J].Plant Cell Environ, 2001(24):755-767.
