孕哺期暴露NP对子代大鼠脑组织基因表达谱的影响*
2014-08-15杨孟雪蒋智钢万齐友庹芳旭封校亚
许 洁,汪 洋,杨孟雪,蒋智钢,万齐友,庹芳旭,封校亚,俞 捷
(1.重庆医科大学 公共卫生学院, 重庆 400016;2. 遵义医学院 公共卫生学院, 贵州 遵义 563099;3. 遵义医学院附属医院 内分泌科,贵州 遵义 563099 )
基因芯片技术是20 世纪 90 年代迅速发展起来的分子生物学技术,它在疾病发生机制和临床诊断等许多生命科学领域已得到广泛的应用,具有快速、准确、多样性、微型化和自动化等优点[1-4]。目前,基因芯片技术在神经毒理学中的应用还属于初始阶段,本研究应用基因芯片技术筛选暴露仔鼠模型海马组织的差异表达基因,拟在基因水平揭示Nonylphnol (NP) 引起仔鼠学习记忆能力降低的可能作用机制。
结合在前期实践过程中遇到的若干问题,遵循对照长江经济带发展和生态文明建设的新要求,本文从顶层设计、系统治理、绿色发展、科技与制度改革创新、水生态文明意识培育五个方面探索提出对城市水生态文明建设的对策建议。
1 材料与方法
1.1 实验动物分组及染毒 选取清洁级Sprague Dawley大鼠性成熟个体,雄鼠15只,雌鼠30只,按雌:雄2:1同笼交配(由第三军医大学实验动物中心提供:SCXK(渝)20070005),获取妊娠雌性SD鼠后随机分为NP组与对照组,于妊娠第6天到出生后21 d灌胃NP和花生油, NP 纯度99%(购于东京化成株试会社)。
1.2.2 差异表达基因的筛选 原始数据用FastQC检测数据质量,使用Tophat进行“序列比对”,再由Cufflink系列软件分析阴性对照组和miR-1254模拟体组的DEG。其中,以倍数变化(fold change,FC)表示单个基因表达变化的倍数。以q<0.05且|log FC|≥1考虑差异有统计学意义。
1.2 样品的合并 NP实验组和对照组21 d仔鼠累积取样各10个,于盛有液态氮的金属溶器中进行样品合并,放液氮罐中备用。
2.3.1.1 分子功能(molecular function)差异表达基因主要涉及的分子功能:结合活性(binding activity)有158个基因上调、催化活性(catalytic activity)有83个基因上调、核酸结合活性(nucleotide binding)有55个基因上调,嘌呤核苷酸结合活性(purine ribonucleotide binding)有50个基因上调等(见图1)。
1.5 差异基因GO(Gene Ontology)分析 GO是基因及基因产物的功能分类体系,分为分子功能(Molecular Function)、生物过程(Biological Process)和细胞组成(Cellular Component)3个部分。
1.4 表达谱芯片杂交及扫描 芯片杂交与洗涤委托上海康成科技有限公司完成。大鼠表达谱芯片采用Roche NimbleGen(NimbleGen Systems, Inc., USA)的12x135K芯片,每张芯片可检测26,419个基因,芯片晾干后,结果采用Axon GenePix 4000B 芯片扫描仪,扫描文件导入GenePix Pro 6.0分析软件。
2 结果
2.1 脑组织RNA提取及质量鉴定结果 对照组和实验组样本提取的总RNA,经核酸浓度测定仪检测,特定波长的光密度值A260/A280比值在1.9~2.1,A260/A230 比值大于1.8,表明提取的总RNA无降解,符合实验要求,且mRNA总量达到芯片分析要求(见表1)。
表1壬基酚暴露和未暴露组脑组织RNA提取及质量鉴定结果

组别OD260/280OD260/230浓度 (ng/μL)对照组2.042.351721.97处理组 2.062.282024.5
2.2 差异表达基因 暴露组与对照组共检测26 419个基因,芯片结果有1 254个差异表达基因,其中619个基因上调,635个基因下调。与神经系统功能相关差异表达基因大致为几方面:周围神经系统发育相关基因,神经胶质细胞发育基因,细胞增殖与分化,细胞凋亡,免疫应答,炎症反应因子,细胞信号传导等方面。
2.3.1.3 细胞组成(cellular component) 表达上调基因的表达产物所涉及细胞成分:胞外区(extracellular region)有27个基因、高尔基体部分(Golgi apparatus part)有8个基因、中间纤维(intermediate filament)有5个等(见图3)。
2.3 基因功能分类
2.3.1 上调基因功能分类
有一个大胡子照相师曾经骑着一匹枣红马来我们村。他第一次来,我以为是您回来了。那天,我远远看见他,不知道怎么就跑上去,问他认不认识一杭。我很希望他说认识。但他说不认识。还反问我认识吗?不过,我还是很喜欢他,他有很粗很黑的胡子,看上去特别威风。爸爸,您也要为我留一把大胡子。
1.3 总RNA的提取及鉴定 用TRIzol提取NP暴露组和对照组脑组织总RNA,紫外分光光度计检测0D值,要求A260/A280比值介于1.8~2.1,A260/A230 比值大于1.8。
2.3.2 下调基因功能分类
小儿急性上呼吸道感染是临床常见的小儿疾病,可见于任何季节,鼻病毒、流感病毒、呼吸道合胞病毒、副流感病毒等是引起本病的主要病原菌,患儿临床多表现为喷嚏、咽部充血、发热等症状。
然后,饲料厂要注重生物安全。在不使用时做好密封,有效地控制害虫,防止灰尘收集系统中的灰尘再循环回到饲料中,并管理好整个工厂的人员移动。
2.3.1.2 生物过程(biological process)表达上调基因参与的主要生物学过程:大分子代谢过程(macromolecule metabolic process)有102基因上调,细胞大分子代谢过程(cellular macromolecule metabolic process) 有92基因上调,蛋白代谢过程(protein metabolic process)有62个基因上调等(见图2)。
2.3.2.1 分子功能(molecular function)表达下调基因经GO分析,分子功能主要分类:结合活性(binding activity)有266个基因下调、催化活性(catalytic activity)有153个基因下调、金属离子结合活性(metalion binding)有95个基因下调、离子结合活性(ion binding)有95个基因下调,阳离子结合活性(cation binding)有95个基因下调等(见图4)。

图1 差异表达上调基因分子功能的分类 图2 差异表达上调基因参与的生物过程分析
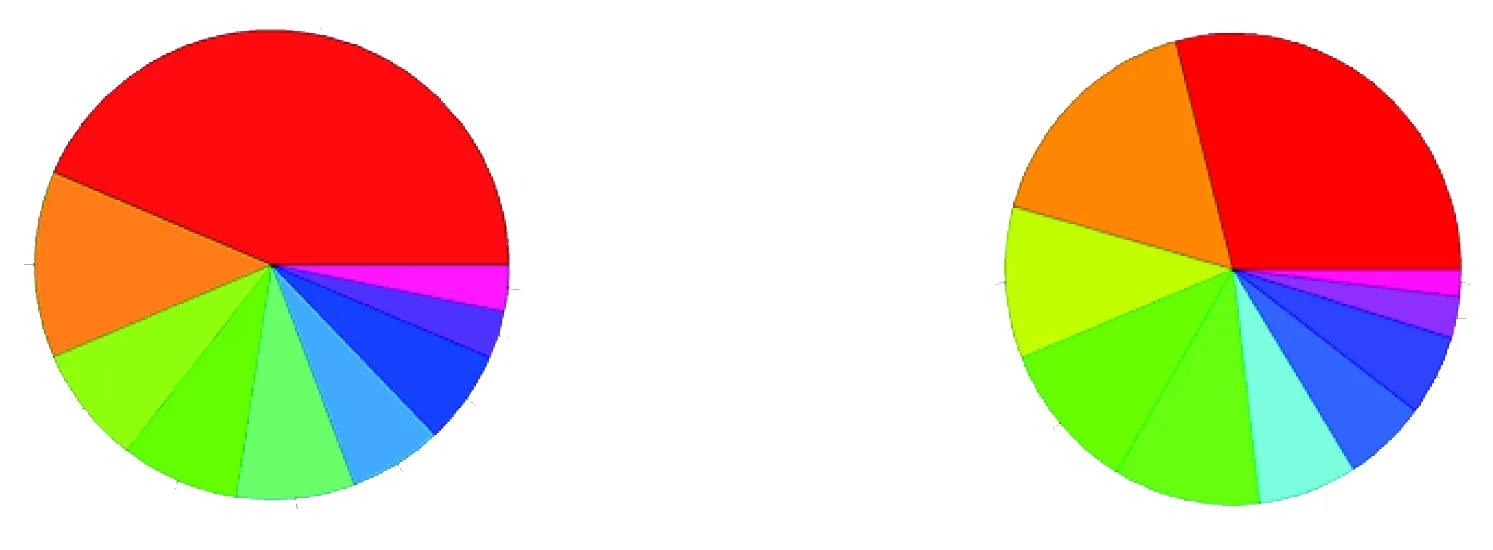
图3 差异表达上调基因表达产物的细胞组成分析 图4 差异表达下调基因的分子功能分类
2.3.2.2 生物过程(biological process)从下调表达基因参与的生物学过程分析结果来看,下调基因参与的主要生物学过程: 细胞代谢过程(cellular metabolic process)有179个基因下调,代谢过程(metabolic process) 有209个基因,氮化合物代谢过程(nitrogen compound metabolic process)有110个基因,细胞氮化合物代谢过程(cellular nitrogen compound metabolic process) 有108个基因,核苷碱基、核苷、核苷酸及核酸代谢过程(nucleobase, nucleoside, nucleotide and nucleic acid metabolic process)有100个基因,发育过程(developmental process)有88个基因等(见图5)。
2.3.2.3 细胞组成(cellular component)表达下调基因的表达产物所涉及细胞成分:细胞内(intracellular)有280个基因,细胞内部分(intracellular part)有264个基因,细胞器(organelle)有231个基因,细胞器部分(intracellular organelle)有228个基因,膜结合细胞器(membrane-bounded organelle)有204个基因,细胞质(cytoplasm)有203个基因等(见图6)。

图5 差异表达下调基因参与的生物过程分析 图6 差异表达下调基因表达产物的细胞组成分析
3 讨论
胚胎期和哺乳期是中枢神经系统发育和功能形成期,外界有害因素的影响可导致脑功能严重缺损。有研究报道NP可导致对小鼠神经毒性作用[5-6],NP和雌二醇[7]或双酚A[8]孕哺期联合染毒能使猴和鼠子代行为发生改变,但有关NP影响脑发育的机制目前还不十分清楚。目前国外有报道将基因表达谱芯片分析技术引入环境内分泌干扰物的体外实验研究[9-11],有关环境内分泌干扰物NP对仔鼠神经发育影响的基因表达谱分析的研究报道较少,关于NP对5日龄子代睾丸以及脑组织的芯片表达谱的研究表明[12-13]:NP对发育中的脑组织有关酶类、神经递质会产生不利影响,也可通过激活caspase-3诱导神经干细胞的凋亡。本实验采用表达谱芯片检测两组差异基因,根据筛选出来的差异表达基因参与的信号通路,为我们进一步有针对性的探索NP神经毒性的机制研究提供方向。
GO分析即基因本体论分析,表达谱芯片筛选的差异表达基因通过GO分析,可得知差异表达基因主要功能。本实验的GO(Gene Ontology)分析显示:差异表达基因主要涉及的分子功能有结合活性、金属离子结合活性、催化活性、核酸结合活性等;参与的主要生物学过程有细胞大分子代谢过程, 细胞氮化合物代谢过程、核苷酸及核酸代谢过程、蛋白代谢过程;所涉及细胞成分包括胞外区、细胞器、高尔基体部分、细胞质、中间纤维等。目前尚无有关NP及其它的内分泌干扰物对成熟期仔鼠脑组织的基因表达谱的报道,故无借鉴资料进行比较,只能对差异表达基因中,现已明确功能的部分基因及表达产物的相关蛋白进行分析,对定位和功能不明的基因,暂不能进行分析,但可为NP对神经系统发育影响机制的研究提供了极为重要的线索,随着未知基因功能逐步明确,将对逐步弄清NP对神经系统分化发育的损伤机制具有重要的意义。
[参考文献]
[1] Guo W,Zhang K M, Tu K, et al. Adipogenesis licensing and execution are disparately linked to cell proliferation [J]. Cell Research, 2009, 19(2):216-223.
[2] Naciff J M, Overmann G J, Torontali S M, et al. Gene expression profile induced by 17 alpha-ethynyl estradiol in the prepubertal female reproductive system of the rat[J]. Toxicol Sci, 2003, 72(2):314-330.
[3] Simunovic F, Yi M, Wang Y, et al. Evidence for gender specific transcriptional profiles of nigral dopamine neurons in Parkinson disease [J]. PLoS One, 2010, 5(1):e8856.
[4] Zhang Y Q,Mao Z,Zheng Y L.Elevation of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Expression in the Mouse Brain after ChronicNonylphenol Exposure [J]. Int J Mol Sci, 2008, 9(10):1977-1988.
[5] 俞捷,张镖,许洁. 环境内分泌干扰物壬基酚对神经系统影响的研究进展[J]. 环境与职业医学, 2013, 30(3): 226-228.
[6] 许洁,汪洋,张镖,等. 壬基酚对子代大鼠皮质部神经生长相关蛋白的影响[J]. 遵义医学院学报, 2014, 37(1):77-80.
[7] Yu J, Fan Q Y, Binli H, et al. Joint neurodevelopmental and behavioral effects of nonylphenol and estradiol on F(1) male rats[J]. Int J Environ Health Res, 2013, 23(4):321-330.
[8] Negishi T, Kawasaki K, Suzaki S, et al. Behavioral alterations in response to fear-provoking stimuli and tranylcypromine induced by perinatal exposure to bisphenol A and nonylphenol in male rats[J]. Environ Health Perspect, 2004 ,112(11):1159-1164.
[9] Naciff J M, Overmann G J, Torontali S M. Gene expression profile induced by 17 alpha-ethynyl estradiol in the prepubertal female reproductive system of the rat[J]. Toxicol Sci, 2003, 72(2):314-330.
[10] Adachi T, Komiyama M, Ono Y. Toxicogenomic effects of neonatal exposure to diethylstilbestrol on mouse testicular gene expression in the long term: a study using cDNA microarray analysis[J]. Mol Reprod Dev, 2002, 63(1):17-23.
[11] 詹平,李元锋,吴德生. 壬基酚对大鼠F1代脑发育相关基因表达的影响[J]. 预防医学情报杂志, 2005, 21(3):257-258.
[12] 张浩. NP雄性生殖发育毒性机理研究[D].成都:四川大学,2006:126-130.
[13] Fitch R H, Denenberg V H. A role for ovarian hormones in sexual differentiation of the brain[J]. Behav Brain Sci, 1998, 21(3):311-327.
