野生侧金盏花的花粉活力与柱头可授性
2014-08-10王阿香陈士惠马翠青杨丽丽
孙 颖,王阿香,陈士惠,马翠青,杨丽丽,何 淼
(1.东北林业大学园林学院,黑龙江 哈尔滨 150040; 2.刘家河镇人民政府林业站,辽宁 凤城 118100)
野生侧金盏花的花粉活力与柱头可授性
孙 颖1,王阿香1,陈士惠1,马翠青1,杨丽丽2,何 淼1
(1.东北林业大学园林学院,黑龙江 哈尔滨 150040; 2.刘家河镇人民政府林业站,辽宁 凤城 118100)
以黑龙江省野生草本地被植物侧金盏花(Adonisamurensis)为对象,对其进行了花粉形态的观测、花粉活力检测方法的筛选,同时进行了花粉活力、柱头可授性的研究,探讨了两者与授粉成功的关系。结果表明,侧金盏花花粉萌发的最适培养基为10%蔗糖+0.1 g·L-1H3BO3+0.1 g·L-1Ca(NO3)2。花粉活力在开花前3 d保持在70%以上,开花第2天活力最大。开花第4天花粉活力开始显著降低,至开花第5天花粉活力下降为31.92%。侧金盏花的柱头在开花当天即有微弱可授性,开花第2天进入强可授期,此时期可维持2~3 d。因开花后2~3 d花药才开裂,为异花授粉提供了机会。但因环境条件的限制及自花花粉空间上的优势,不排除其存在一定的自交率。
侧金盏花;花粉活力;柱头可授性;检测方法
侧金盏花(Adonisamurensis)为毛茛科多年生草本植物,又名冰凉花、福寿草、顶冰花,一般生长在东北地区山坡或山脚的灌木丛间、阔叶林下以及林缘地上[1]。花黄色艳,成株株高30~40 cm,3月中下旬即可发芽、开花;适应性强,建植、栽培、养护成本低,是一种有很大开发潜力的珍奇野生花卉。侧金盏花是高寒地区组成缀花草坪、构建护坡、布置花境花坛的优秀植物材料,可广泛应用于公园、庭院、乡村道路、高尔夫球场及待开发地带,形成色彩丰富、层次多样的草坪及园林景观。侧金盏花在国外已经有100多年的栽培历史,培育出100多个花色品种[2],我国则基本处于野生状态[3],针对其的研究主要集中在分类[4-5]、药理成分[6]、抗寒性[7-8]等方面。为更充分地开发侧金盏花野生种质资源,使其能够广泛应用于北方高寒城市绿地,对其进行系统地野外观察及引种试验,认识整个开花过程的自然规律及有性繁殖特性显得十分必要。
传粉是种子植物有性生殖的必经阶段,传粉系统由花粉、柱头、传粉媒介3个部分组成。一般人们把有花植物的传粉过程分成3个阶段:花粉从花药中释放;花粉从父系结构(花药)向母系结构(柱头)传送;花粉成功地落置于柱头表面继而萌发[9]。花粉从花药传送到柱头表面是一个复杂的过程,需要借助一定的载体,经过一定的空间。正是由于花粉携带精子运动的复杂传粉过程,为物种的进化提供了契机,成为植物进化生物学和繁殖生物学关注的焦点[10-12]。正因如此,花粉活力的高低及柱头可授性的强弱就成为植物生殖成功与否、结实率高低的关键因素之一。近年来,花粉活力与柱头可授性的研究引起很多学者的重视[13-17]。在自然条件下侧金盏花以种子进行繁殖,掌握其花粉活力和柱头可授性对于进行侧金盏花的种子生产和良种选育,提高侧金盏花的制种效率,以及进一步进行种质栽培和基地建设具有重要意义。
1 材料与方法
1.1 试验材料
试材为分布于黑龙江省哈尔滨市帽儿山老山人工林试验站(45°20′ N,127°34′ E)的野生侧金盏花。选择具有代表性的落叶松林样地,于样地内取10个2 m×2 m样方,以样方内正常生长的侧金盏植株作为研究对象。
1.2 试验方法
1.2.1 花粉及柱头综合特征观察 在侧金盏花花蕾期,选取10个样地侧金盏花的花朵共100枚进行挂牌标记。花朵开放前,每天观察一次。开花当天及第2天从07:00观察到17:00,随后每天观测一次,直到花瓣萎蔫。记录花朵的各部分器官发生变化的时间、动态过程,注意花粉散出、花柱弯曲的情况。
1.2.2 花粉及柱头微观结构观察 于样地随机选取10枚新鲜的侧金盏花花朵,对花粉及柱头样品经过固定、脱水、粘台、喷金等处理后,置于扫描电镜下观察表面结构并拍照。
1.2.3 花粉活力测定方法的筛选 对比TTC法[18]、I2-KI法[19]和离体萌发法[18]的试验结果,以确定花粉活力的测定方法。其中离体萌发法的具体操作方法为:将盛花期混合均匀的侧金盏花花粉散播在蔗糖浓度100 g·L-1,H3BO30.05 g·L-1,Ca(NO3)20.1 g·L-1的培养液中,置于30 ℃的恒温培养箱培养2 h后,于160×显微镜下统计3个视野花粉的萌发率,3次重复。
萌发率的计算:在显微镜下观察花粉的萌发情况。以花粉管长度大于花粉粒直径为萌发标准,读取一个视野中的花粉总数和萌发花粉数,以此计算萌发率。
1.2.4 花粉离体萌发最适培养液的筛选 采用3因素3水平L9(34)正交设计。蔗糖、Ca(NO3)2、H3BO3为3因素,具体用量见表1。此外,其他元素的配制用量为:琼脂9 g·L-1、KNO30.2 g·L-1、MgSO40.1 g·L-1。试验共设9个处理,每处理3次重复。根据试验结果确定各成分的加入量。
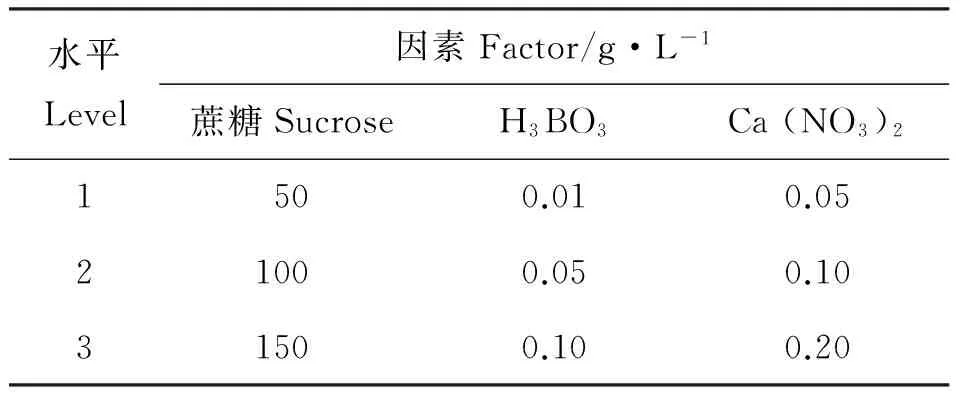
表1 L9(34)正交设计的因素与水平
1.2.5 不同开花天数花粉活力的测定 开花前1天对开放程度大致相同的侧金盏花花蕾进行套袋,同时进行标记。第2天清晨将硫酸纸袋撤掉,使其同时开放,每天10:00-11:00采集10枚花的花粉混匀播于最适培养液中。连续观测5 d,记录花粉萌发率。
1.2.6 柱头可授性检测 采用两种方法进行柱头可授性的检测:1)对开花不同天数的柱头进行粘液分泌情况的观察,主要借助放大镜进行外观的观察,同时用联苯胺-过氧化氢的方法检测柱头的可授性[13]。每个时期检测30枚花的柱头。2)人工授粉。在开花前1天,开花第1、2、3、4、5天分别对60枚单花进行去雄、套袋的人工授粉试验。花粉来源为异花花粉。统计授粉后的座果率情况。
2 结果与分析
2.1 侧金盏花雄蕊及柱头的形态特征
2.1.1 开花过程中侧金盏花雄蕊及柱头的形态变化 侧金盏花花朵初开时直径较小,花瓣为黄绿色。随着花被的增长逐渐变为鲜黄色,花朵直径逐渐变大。花药与花被同步生长,开花后2~3 d外轮花药开始开裂,其余交替开裂,当天气晴朗时所有花药全部开裂大约需要1 h,当天气阴冷时花药散粉持续时间较长,常长达1~2 d。侧金盏花花药平展于花瓣上,散粉过程中花丝逐渐伸长到直立,进而向心发育,包围于雌蕊群之上(表2)。
2.1.2 侧金盏花花粉及柱头的微观形态特征 侧金盏花花粉为单粒花粉,形状为椭球形(图1)。花粉极面观为三裂圆形,赤道面观为纺锤形。具三沟,沟宽4.5~5.0 μm,长至近两极。沟端圆钝,沟沿部整齐不加厚,沟膜具密集小刺。花粉外壁表面亦有均匀分布的刺状凸起,小刺长0.2~0.3 μm。花粉粒大小约为31.57 μm×22.51 μm。侧金盏花的柱头相对表面积较大,柱头表面有很多长柱状凸起。柱头成熟后表面有粘液出现,这有助于黏附和嵌入花粉(图1)。
2.2 侧金盏花花粉活力测定方法的确定
选用I2-KI染色法和TTC染色法测得的侧金盏花花粉活力分别为25.18%和57.41%,离体萌发法测得的侧金盏花花粉活力为70.15%,三者之间差异显著(P<0.05)。因此,选择离体萌发法进行侧金盏花花粉活力测定。
通过不同培养液的花粉萌发试验发现(表3),蔗糖、H3BO3、Ca(NO3)2在不同程度上对侧金盏花的花粉萌发有影响。3个蔗糖水平下的侧金盏花的花粉萌发率分别为20.33%、44.58%、29.79%,3个H3BO3水平下侧金盏花的花粉萌发率分别为30.63%、30.36%、33.71%,3个Ca(NO3)2水平下侧金盏花的花粉萌发率分别为23.02%、43.37%、28.31%。极差对比发现,蔗糖的影响最大,Ca(NO3)2次之,影响最小者为H3BO3。三者对侧金盏花花粉萌发率的影响均不显著(表4)。
2.3 侧金盏花花粉活力检测结果
检测发现,侧金盏花的花粉在开花前即有活力,开花前1天的花粉活力达到62.22%,与开花当天的花粉活力持平。随后花粉活力随开花天数增加升高,至开花第2天花粉活力达到最高的74.09%。开花第4天花粉活力下降至53.55%,开花第5天花粉活力降至31.92%。开花第2天和第3天花粉活力显著高于其他(P<0.05)。侧金盏花花粉活力在开花的前3 d基本保持稳定。

表2 侧金盏花开花过程雄蕊与柱头形态特征变化

图1 侧金盏花花粉、柱头微形态
注:1~2,花粉形态;3,花粉外壁;4~5,柱头形态;6,花粉散落于柱头表面的状态。
Note: 1~2,Pollen morphology;3,Outer layer of pollen;4~5,Stigma morphology;6,Pollination of pollen and stigma (SEM).

表3 不同蔗糖、H3BO3、Ca (NO3)2水平对花粉萌发的影响

表4 蔗糖、H3BO3和Ca (NO3)2在不同水平下花粉萌发率的显著性分析
2.4 侧金盏花柱头可授性检测结果
侧金盏花在开花前1天(此时花药被花被片包被,与柱头存在空间隔离)无粘液分泌,柱头颜色为浅绿色,不具有可授性;开花当天部分花朵柱头开始分泌粘液,柱头进入微弱的可授期;至开花第2天时柱头开始分泌大量粘液,柱头颜色变为黄绿色,进入较强可授期,此状态一直维持3 d;开花第4天时柱头可授性变弱,粘液分泌量减少;开花第5天时已经有部分柱头失去可授性,颜色由黄色变为黄褐色;在花末期,粘液分泌已停止,柱头颜色已转为深褐色,此时侧金盏花的子房开始发育,但花朵仍可开放3~4 d,直至完全枯萎。

表5 侧金盏花的柱头可授性及粘液分泌情况
注:-表示柱头不具有可授性;-/+表示部分柱头具有可授性;+表示柱头具有可授性;++表示柱头可授性较强。
Note: -means stigmas are not receptive;-/+means some stigmas are receptive;+means stigmas are receptive;++means stigma receptivity is strong.
通过对侧金盏花进行不同开花天数的人工授粉检测发现,在开花前1天进行授粉的植株没有成功座果。开花当天的座果率为61.67%,开花第2和3天时的座果率则超过75%;开花第4天时的座果率又降为46.67%;开花第5天时座果率急速下降,仅为6.67%。说明侧金盏花在开花当天即可进行授粉,在开花第2天时柱头可授性开始加强,授粉成功率升高。此结果与上述的联苯胺-过氧化氢检测结果是一致的。
3 讨论
花粉在植物有性繁殖中传递雄性亲本的遗传信息[15,20]。侧金盏花的花粉约为31.57 μm×22.51 μm,按照埃尔特曼的花粉划分标准[21],这种大小的花粉属于小孢粉。侧金盏花的花粉虽然体积小,但其形状为椭球形,且外壁较粗糙,具有刺状突起,有利于黏附在昆虫身上进行传播,这是典型的虫媒花特征[22]。侧金盏花的花粉在开花前即具有活力,在开花前3 d活力基本稳定在70%左右,花后第5天仍有31.92%的花粉具有活力,活力较高。一方面,这是由遗传因素所决定,另一方面也受到环境条件的影响。有研究认为温度、湿度和光照是影响花粉活力的主要因素[23]。很多相关研究也证明了这一点:如刺五加(Eleutherococcussenticosus)在光照条件好的生境中花粉活力在开花之初较高但下降迅速,而在光照条件较差的生境中花粉活力在开花之初较低但下降较慢[20];连翘(Forsythiasuspense)花粉在室温环境下能存活5 d,而在低温干燥环境下能存活10 d[24];地被菊‘毛白玉’(Dendranthemamorifolium‘Maobaiyu’)的花粉活力在全光照环境中的活力超过35%,而在60%遮阴环境中的活力低于20%[25]。侧金盏花的花粉生活环境相对气温低,能减弱花粉的代谢作用,减少营养物质的消耗,保持酶的活性。花粉较长时间保持高活力对授粉具有积极意义。这一特点为短期保存侧金盏花粉提供有益启示,同时可以采集开花后3 d内的花粉进行人工授粉,为侧金盏花的引种栽培提供技术保证。
柱头可授性是花朵成熟过程中一个重要时期,其能在很大程度上影响植物自交率[16]。不同植物的柱头可授期时间从几小时到十几天不等。如刺五加的柱头在开花后5~9 d具有可授性,而其花粉寿命只有4 d,说明其基本没有自花授粉的机会[20];连翘柱头从开花当天至开花第4天均有可授性,花粉活力较强期为花后第2天,说明其存在一定自花授粉能力[24]。同时,花朵开放期间,柱头有无分泌物对其授粉效果也有影响。侧金盏花的柱头在开花当天即分泌少量粘液,开花第2天粘液大量分泌,说明其属于湿型柱头,这种柱头可作为昆虫的诱物黏着花粉,又可以为花粉萌发提供必要的物质。经联苯胺-过氧化氢检测发现,侧金盏花开花当天部分柱头即具备可授性,可授期可维持4 d左右。不同开花天数的人工授粉试验也证明从开花第1天至开花第4天柱头均可成功授粉。但由于花药在开花后2~3 d才开始开裂,这就造成了花药散粉与部分柱头可授期时间上一定的间隔,为异花授粉提供了可能性。花粉和柱头在出现时间上的不同步是一种较为常见的限制自花传粉的机制[26-28]。但植株的自交率不仅取决于自身授粉的花粉量,而且取决于它从种群内其他个体那里接受外来花粉的数量、时机,尤其是取决于传粉媒介对花粉的有效传送[29-31]。对于侧金盏花来说,其花期在北方的3-4月,此时常常冰雪尚未消融,传粉昆虫的数量、访花频率、传粉效率较难得到保证。另外,侧金盏花在开花第2天花药开裂时,花粉活力达到最大,此时也是其柱头的强可授期,对于开花第1天未能成功授粉的植株来说,此时花丝变长,花药与柱头之间间距变短,自花花粉具有空间上的优势,不能排除其存在一定的自花授粉机会。在这种情况下自交可以提高植物的结实率,提高植株的生殖适合度。侧金盏花的自交率是多少,在多大程度上产生繁殖保障效应,及其随环境条件的改变是否产生变化、变化的幅度,需要通过更加详细的人工授粉试验及环境控制试验来进一步明确。
4 结论
侧金盏花花粉体积较小但外壁较粗糙,具有刺状突起,是典型的虫媒花花粉,其适宜的萌发培养基为10%蔗糖+0.1 g·L-1H3BO3+0.1 g·L-1Ca(NO3)2。经萌发培养检测发现,侧金盏花花粉活力在开花前3 d保持在70%以上,开花第2天活力最大。开花第4天开始花粉活力显著降低,至开花第5天花粉活力比前3 d降低50%以上。经联苯胺-过氧化氢法检测和不同开花天数的人工授粉试验发现,侧金盏花的柱头在开花当天即有微弱可授性,开花第2天进入强可授期,此时期可维持2~3 d。因开花后2~3 d花药才开裂,为异花授粉提供了机会。但因环境条件的限制及自花花粉空间上的优势,不排除其存在一定的自交率。
[1] 李欢,肖筝,王本全,李铁松,姜长阳.侧金盏花组织培养及无性系建立的研究[J].特产研究,2010,32(1):22-28.
[2] 万清林,刘鸣远.侧金盏花成株年生长节律生理特性的研究[J].植物研究,1996,16(3):351-355.
[3] 王宗霞,秦荣,张淑艳,刘欣.侧金盏的引种试验研究[J].内蒙古科技与经济,2003(10):102-103.
[4] 王文采.侧金盏花属修订(一)[J].植物研究,1994,14(1):1-31.
[5] 王文采.侧金盏花属修订(二)[J].植物研究,1994,14(2):108-138.
[6] 傅翔,张汉明.侧金盏花属植物成分及药理研究进展[J].植物资源与环境,1996,4(3):56-60.
[7] 曲彦婷,唐焕伟,陈菲.低温胁迫对侧金盏部分生理生化指标的影响[J].国土与自然资源研究,2009(3):82-83.
[8] 曲彦婷.早春花卉侧金盏的抗寒性研究[J].国土与自然资源研究,2009(4):90-91.
[9] 曹坤方.植物生殖生态学透视[J].植物学通报,1993,10(2):15-23.
[10] Fenster C B,Armbruster W S,Wilson P,Dudash M R,Thomson J D.Pollination syndromes and floral specialization[J].Annual Review of Ecology,Evolution and Systematics,2004,35:375-403.
[11] 任明迅.雄蕊合生植物半边莲的花部综合特征与繁育系统[J].植物生态学报,2009,33(2):361-368.
[12] Barrett S C H.Sexual interference of the floral kind[J].Heredity,2002,88:154-159.
[13] 叶要妹,张俊卫,齐迎春,陈天花,包满珠.百日草柱头可授性和花粉生活力的研究[J].中国农业科学,2007,40(10):2376-2381.
[14] 丁春邦,李强,李燕,张利,杨瑞武,周永红.重楼属9种5变种花粉活力与柱头可授性特性研究[J].草业学报,2009,18(4):61-66.
[15] 刘帮龙,张晓慧,干友民,贺啸尘,莫利.野生马蹄金花粉生活力检测方法比较[J].草业科学,2011,28(11):1941-1944.
[16] 赵金花,李青丰,那仁图雅,斯琴.3种野生葱属植物花粉活力和柱头可授性研究[J].草业科学,2010,27(4):93-96.
[17] 刘林德,张萍,张丽,陈磊,高玉葆.锦带花的花粉活力、柱头可授性及传粉者的观察[J].西北植物学报,2004,24(8):1431-1434.
[18] 张志良.植物生理学指导[M].北京:高等教育出版社,1990:237-243.
[19] 胡适宜.植物胚胎学实验方法(一)花粉生活力的测定[J].植物学通报,1993,10(2):60-62.
[20] 刘林德,张洪军,祝宁,申家恒.刺五加花粉活力和柱头可授性的研究[J].植物研究,2001,21(3):375-379.
[21] Erdtman.花粉形态与植物分类[M].王伏雄,钱南芬,译.北京:科学出版社,1962.
[22] 周凌瑜,刘群录,邵邻相.虫媒花与风媒花花粉形态的比较[J].上海交通大学学报:农业科学版,2008,26(3):177-182.
[23] 乔红莲,刘忠华,霍喜颖.贮藏温度对百合花粉生活力的影响[J].北方园艺,2010(5):86-89.
[24] 铁军,金山,茹文明,张志翔.连翘花粉活力和柱头可授性研究[J].山西大学学报自然科学版,2008,31(4):604-607.
[25] 吕晋慧,赵耀,王媛,冯雁梦,李艳锋,孔一昌.地被菊花粉活力和储藏性研究[J].园艺学报,2012,39(12):2483-2490.
[26] Wu Z K,Zhang C Q.Comparative study of pollination biology of two closely related alpinePrimulaspecies, namelyPrimulabeesianaandP.bulleyana(Primulaceae)[J].Journal of Systematics and Evolution,2010,48(2):109-117.
[27] María J A,José M I,Adrián E, Elena T.Dissecting components of flowering pattern:Size effects on female fitness[J].Botanical Journal of the Linnean Society,2008,156(2):227-236.
[28] 任明迅,姜新华,张大勇.植物繁殖生态学的若干重要问题[J].生物多样性,2012,20(3):241-249.
[29] Holsinger K E.Pollination biology and the evolution of mating systems in flowering plants[J].Evolutionary Biology,1996,29:107-149.
[30] Buide M L,Díaz-Peromingo J A,Guitián J.Flowering phonology and female reproductive success inSileneacutifoliaLink ex Rohrb[J].Plant Ecology,2002,163(1):93-103.
[31] Brys R,Jacquemyn H,Endels P,Rossum F V,Hermy M,Triest L,Bruyn L D,Blust G D E.Reduced reproductive success in small populations of the self-incompatiblePrimulavulgaris[J].Journal of Ecology,2004,92:5-14.
(责任编辑 武艳培)
Research on pollen viability and stigma receptivity ofAdonisamurensis
SUN Ying1, WANG A-xiang1, CHEN Shi-hui1, MA Cui-qing1, YANG Li-li2, HE Miao1
(1.College of Landscape and Architecture, Northeast Forestry University, Harbin 150040, China;2.Forestry Station of Liujiahe Town People’s Government, Fengcheng 118100, China)
In order to study pollen morphology, pollen viability and stigma receptivity, an investigation on pollen and stigma ofAdonisamurensisin Heilongjiang, China was conducted. The relationships between pollen viability, stigma receptivity and pollination success were also analyzed. The results showed that the best medium for pollen germination was 10% sucrose + 0.1 g·L-1H3BO3+0.1 g·L-1Ca (NO3)2. Pollen viability maintained more than 70% at the first three days of blossom. The highest pollen viability was observed at the second day and decreased significantly since the fourth day. Pollen viability was 31.92% at the end of florescence. Stigma had weak receptivity ability at the first day and strong receptivity ability since the second day which maintained 2~3 days. Pollen had germination ability at the first day of blossom and anther crack delayed 2~3 days which provided the opportunity for cross-pollination. At the same time, there was the possiblity of self-fertilize for the environmental constraints and space advantage of self pollen.
Adonisamurensis; pollen viability; stigma receptivity; detection method
HE Miao E-mail:hemiao_xu@126.com
10.11829j.issn.1001-0629.2013-0459
2013-08-14 接受日期:2013-09-03
中央高校基本科研业务费专项资金项目(DL13BA08)
孙颖(1979-)女,吉林长春人,副教授,博士,主要研究方向为园林植物种质资源开发与利用。E-mail:littlesuning@126.com
何淼(1975-)女,辽宁本溪人,副教授,博士,主要研究方向为园林植物培育、养护管理。E-mail: hemiao_xu@126.com
Q949.746;Q944.42
A
1001-0629(2014)06-1045-07*1
