横断山区中缅树鼩肥满度的初步研究*
2014-08-02高文荣周庆宏朱万龙张浩姜文秀王政昆
高文荣, 周庆宏, 朱万龙, 张浩, 姜文秀, 王政昆
(1.云南师范大学 生命科学学院,云南 昆明 650500;2.昆明市海口林场,云南 昆明 650114)
自Fulton提出肥满度(relative fatness)后,肥满度就被作为一个判断动物对环境适应的生理状态和营养状况的综合指标,广泛用于动物生长状况与年龄、性别、温度、光照、季节、食物、种群密度关系和种间种内关系的研究[1].因此,肥满度的研究历来受到国内外学者的重视.自夏武平等[2]首次把在鱼类中使用的肥满度应用于红背鼠平(Clethrionomysrutilus)以来,已经在很多属种中得到了广泛应用,如大林姬鼠(Apodemuspeninsulae)[3]、黄毛鼠(Rattusrattoides)[4]、小家鼠(Musmusculus)[5]、灰仓鼠(Cricetulusmigratorius)[6]、达乌尔黄鼠(Spermophilusdauricus)[7]、中华鼢鼠(Myospalaxfontanieri)[8]、黑线仓鼠(C.barabensis)[9]、大仓鼠(C.triton)[10]、布氏田鼠(Microtusbrandti)[11]、黑线姬鼠(A.agrarius)[12]、褐家鼠(R.norvegicus)[13]、社鼠(R.niviventer)[14]、东北鼢鼠(Myospalaxpislurus)[15]、大沙鼠(Rhombomysopimus)[16]等.
横断山区是我国特有的高山峡谷,地势起伏大,哺乳动物种类多,古老种特有种和比例高[17,34-35],被誉为“第四纪冰期动物的避难所”[18].该地区环境复杂多样且地理位置特殊,南部与东南亚热带北端相邻,北部及西北部向高寒的青藏高原地带过渡,此外,该地区南北流向的澜沧江、怒江、金沙江及其支流峡谷,有利于南北生物的交流[19].中缅树鼩(Tupaiabelangeri)为攀鼩目(Scandentia)树鼩科(Tupaiidae)动物,主要分布于亚洲热带亚热带地区,是东洋界特有的小型哺乳动物,中缅树鼩是攀鼩目中分布纬度最高和分布最广的一个种;我国云南、贵州和四川西南部很可能是树鼩目分布的北限[20].2006年,列入IUCN濒危动物,并被作为生物医学模型动物.关于中缅树鼩的生理生态学研究已有较多报道,如体温调节和产热特征的关系[21],蒸发失水[22],光驯化和冷驯化过程中产热、能量收支和血清瘦素变化[23-24]的研究等.本文以生活在横断山区的中缅树鼩为实验对象,研究了中缅树鼩肥满度与性别、年龄和季节变化的关系,以及禁食和重喂食对中缅树鼩肥满度的影响,为探讨横断山区小型哺乳动物的生理生态适应模式和适应对策提供一些理论依据.
1 研究方法
1.1 材料和方法
实验动物于2012年的9月和12月与2013年的3月和6月捕自云南大理州剑川县石龙村的农田和灌丛(26°22′ N,99°48′ E),海拔2 550~2 615 m.该地区温带季风气候特征明显,干湿季节分明.6月份的平均温度最高,为25.04 ℃,比同纬度的平原地区低,随海拔高度的增加气温显著降低.共捕获中缅树鼩80只,其中,雌性36只,雄性44只.对捕获的动物样本分别进行编号,年龄、性别鉴定,逐一测量体长、体重等有关数据.然后将实验动物带回云南师范大学生命科学学院实验动物房(昆明),单笼饲养(饲养笼规格为:120 mm×200 mm×300 mm),无巢材,饲养温度为(25 ℃±1 ℃),光照条件为12L:12D,以标准雏鸡饲料(昆明生产的产蛋雏鸡饲料)(69.1%)、全脂奶粉(2.7%)、白糖(6.9%)以及精面粉(21.3 %),熟喂动物,每隔1天,加喂少许水果(苹果,梨等).动物适应至少2周后,即进行相关的实验.
实验选取体重相近的中缅树鼩56只(测定体重与体长并计算肥满度,其肥满度差异不显著)随机分为7组(每组8只,):对照组(自由取食,F0h);分别禁食12 h(F12h)组、24 h(F24h)组和36 h(F36h)组;以及禁食36 h后重喂食12 h(R12h)组、48 h(R48h)组和7 d(R7d)组.实验结束后测定动物的体重和体长.采用公式 K=100 W/L3计算肥满度,其中,W为体重(g);L为体长(cm);K为肥满度(g/cm3).年龄组用体重与体长相对比较并结合其生物学性状进行划分,包括未成年鼠(幼鼠、亚成年鼠)和成年鼠两大类.一般肥满度值越大,表明鼠类身体状况越好,反之亦然.
1.2 数据分析
数据处理采用SPSS 16.0软件包.采用独立样本t-检验和单因素方差分析(one-way ANOVA)的方法进行数据分析,数据全部表示为平均值±标准误差(Mean±SE),P<0.05表示差异显著,P<0.01表示差异极显著.
2 结果
2.1 肥满度与性别的关系
对80只中缅树鼩的肥满度统计结果显示,其肥满度变动幅度较大,变幅为1.30~3.14 g/cm3,总平均肥满度为(1.98±0.43)g/cm3,其中,雌性肥满度平均为(2.04±0.09)g/cm3,雄性肥满度平均为(1.95±0.06)g/cm3,差异不显著(t=-1.129,P>0.05)(如表1所示).
2.2 肥满度与年龄的关系
对不同年龄组的中缅树鼩肥满度进行比较,其中,未成年中缅树鼩肥满度平均为(2.05±0.04) g/cm3,成年中缅树鼩肥满度平均为(1.95±0.05) g/cm3,成年个体和未成年个体肥满度之间差异不显著(t=-1.511,P>0.05)(如表2所示).

表2 中缅树鼩不同年龄肥满度的变化
2.3 肥满度的季节变化
对2012年9月和12月与2013年3月和6月捕捉到的中缅树鼩进行肥满度统计,结果显示,中缅树鼩平均肥满度以12月最高,6月最低.肥满度的季节变化趋势为:12月最高,平均为(2.20±0.32) g/cm3;6月最低,平均为(1.76±0.27) g/cm3;9月较高,平均为(2.10±0.28) g/cm3;3月较低,平均为(1.86±0.18) g/cm3.单因素方差分析结果表明:中缅树鼩肥满度的季节变化差异极显著(F3,76=5.201,P<0.01,n=80)(如图1所示).

图1 中缅树鼩肥满度的季节变化.数值为平均值±标准误差,柱上不同字母表示组间差异显著(P<0.05)
2.4 禁食和重喂食对肥满度的影响
禁食和重喂食极显著影响中缅树鼩的肥满度(F6,49=5.216,P<0.01),禁食后肥满度降低,禁食24 时达到极显著水平,禁食36 h时最小,平均为(1.45±0.09) g/cm3,重喂食7 d后能恢复到对照组水平(如图2所示).
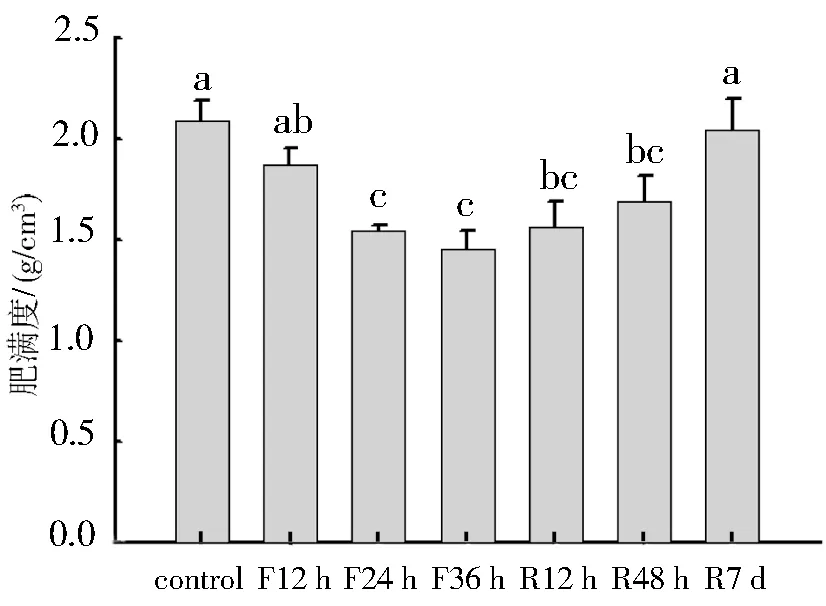
图2 禁食和重喂食对中缅树鼩肥满度的影响.F12 h:禁食12 h组;F24 h:禁食24 h组;F36 h:禁食36 h组;R12h:禁食36 h后重喂食12 h组;R48h:禁食36 h后重喂食48 h组;R7d:禁食36 h后重喂食7 d.数值为平均值±标准误差,柱上不同字母表示组间差异显著(P<0.05).
3 讨 论
3.1 肥满度与性别、年龄的关系
不同性别中缅树鼩的肥满度差异不显著,这可能是因为中缅树鼩个体较小,雌雄个体活动范围差异不大,消耗能量相似的缘故[13].这与大林姬鼠、小家鼠、达乌尔黄鼠、褐家鼠等报道一致[3,5,7,13].还有一些研究表明,有些种类的动物肥满度两性存在差异,如灰仓鼠、黑线仓鼠、大仓鼠、社鼠、东北鼢鼠等[6,9-10,14-15].由表2可以看出,未成年中缅树鼩的肥满度高于成年中缅树鼩,这可能是因为未成年中缅树鼩在亲代中缅树鼩的抚育照料下,营养条件较好,导致生长发育旺盛,性成熟速度快,身体状况略优于成年鼠[13].这与小家鼠、灰仓鼠、达乌尔黄鼠、黑线仓鼠等报道一致[5-7,9].
3.2 肥满度的季节变化
中缅树鼩肥满度的季节变化趋势为:12月最高,6月最低,9月较高,3月较低,平均为(1.98±0.43)g/cm3,这可能与中缅树鼩特殊的生存环境有关.有研究表明冷驯化条件下中缅树鼩体重显著增加,脱冷驯化后降低[23],而中缅树鼩生活的横断山区是典型的高海拔低纬度地区,冬季来临时,环境温度较低(相当于冷驯化),中缅树鼩体重显著增加,但体长变化不大,因此12月肥满度最高;夏季时,环境温度较高(相当于脱冷驯化),中缅树鼩体重显著降低,同样体长变化不大,故6月肥满度最低.9月的食物资源比3月丰富,而环境温度又比较适宜,因而中缅树鼩肥满度表现出9月较高,3月较低.
3.3 肥满度与禁食和重喂食的关系
食物是影响小型哺乳动物体重和能量代谢的重要环境因子[25-26].能量摄入和支出平衡的变化最终决定体重的适应性调节,进而影响动物的肥满度.我们的研究发现,禁食和重喂食极显著影响中缅树鼩的肥满度,禁食后肥满度降低,禁食24 h时达到极显著水平,禁食36 h时最小,平均为(1.45±0.09) g/cm3,重喂食7 d后能够恢复到对照组水平.可能是因为禁食后,中缅树鼩体重下降,重喂食后又回升,而短时间内其体长基本没有变化所致.食物资源变化诱导许多小型哺乳动物体重调节,如布氏田鼠(Lasiopodomysbrandtii)[27],大鼠[28],小鼠[29],叙利亚仓鼠(Mesocricetusauratus)[30-31]和西伯利亚仓鼠(Phodopussungorus)[32-33].另外,中缅树鼩属于杂食性动物,食物资源的可获得性至关重要.基于以上分析,短时间的禁食与重喂食条件下,中缅树鼩体重变化显著,而体长基本保持不变,从而导致其肥满度变化显著.
4 结 论
中缅树鼩雌雄个体肥满度之间差异不显著;不同年龄组之间肥满度差异不显著;不同季节的肥满度差异极显著,12月最高,6月最低;禁食和重喂食极显著影响中缅树鼩的肥满度,禁食后中缅树鼩的肥满度降低,禁食24 h时达到极显著水平,禁食36 h时最小,重喂食7 d后能够恢复到对照组水平.总之,中缅树鼩肥满度在不同条件下表现出来的变化模式,可能与其生存的自然环境密切相关.所以肥满度可以作为预测种群数量变动的一项指标.
参 考 文 献:
[1] 孙儒泳.动物生态学原理(第二版)[M].北京:北京师范大学出版社,1992.
[2] 夏武平,孙崇潞.红背鼠平肥满度的研究[J].动物学报,1963,15(1):33-34.
[3] 夏武平,孙崇潞.大林姬鼠肥满度的研究[J].动物学报,1964,16(4):555-566.
[4] 秦耀亮.黄毛鼠肥满度的研究[A].广东省动物学会论文集[C].广州:广东省动物学会出版社,1981,90-96.
[5] 严志堂.小家鼠肥满度的研究[J].兽类学报,1983,3(2):173-180.
[6] 钟明明,严志堂.灰仓鼠肥满度的研究[J].兽类学报,1984,4(4):273-282.
[7] 刘振才,王成贵,王琛.达乌尔黄鼠肥满度的研究[J].兽类学报,1990,10(1):66-70.
[8] 李晓晨,王廷正,刘加坤,等.中华鼢鼠肥满度的研究[J].陕西师范大学学报,1991,19(增):122-125.
[9] 周延林,候希贤,董维惠,等.黑线仓鼠肥满度的研究[J].兽类学报,1992,12(3):207-212.
[10]李晓晨,王廷正.大仓鼠肥满度的研究[J].兽类学报,1992,12(4):275-279.
[11]刘志龙,李仲来.布氏田鼠肥满度的研究[J].中国媒介生物学及控制杂志,1993,4(5):362-366.
[12]杨再学.黑线姬鼠肥满度的研究[J].兽类学报,1995,15(1):73-74.
[13]杨再学,郭世平.褐家鼠肥满度的研究[J].生态学杂志,1995,14(6):25-28.
[14]高枫,鲍毅新.舟山岛与金华北山社鼠肥满度的比较与分析[J].兽类学报,1995,15(4):272-309.
[15]王俊森,李云芳,程继臻.东北鼢鼠满度的研究[J].国土与自然资源研究,1996,7(4):50-51.
[16]张忠兵,赵天飙,曹广成,等.大沙鼠满度的研究[J].中国媒介生物学及控制杂志,2002,13(4):250-252.
[17]张荣祖.中国动物地理[M].北京:科学出版社,1999.
[18]陈灵芝.中国生物多样性[M].北京:科学出版社,1993.
[19]吴征镒,王荷生.中国自然地理-植物地理[M].北京:科学出版社,1985.
[20]王应祥,李崇云,马世来.树鼩的分类与生态//彭燕章,叶智彰,邹如金.树鼩生物学[M].昆明:云南科学技术出版社,1991.
[21]谢静,朱莉萍,王政昆.冷暴露对中缅树鼩适应性产热特征的影响[J].兽类学报,2009,29(1):50-58.
[22]黄春梅,胡黎娅,杨盛昌,等.中缅树鼩体温、代谢率和蒸发失水日节律[J].动物学杂志,2012,47(2):127-135.
[23]ZHANG L,ZHANG H,ZHU W L,et al.Energy metabolism, thermogenesis and body mass regulation in tree shrew (Tupaia belangeri) during subsequent cold and warm acclimation[J].Comparative Biochemistry and Physiology,Part A,Molecular & Integrative Physiology,2012 ,162(4):437-442.
[24]ZHANG L,ZHU W L,WANG Z K.Role of photoperiod on hormone concentrations and adaptive capacity in tree shrews,Tupaia belangeri[J].Comparative Biochemistry and Physiology,Part A,Molecular & Integrative Physiology,2012,163(3-4):253-259.
[25]VELOSO C,BOZINOVIC F.Dietary and digestive constraints on basal energy metabolism in a small herbivorous rodent[J].Ecology,1993,74(7):2003-2010.
[26]CORK S J.Digestive constraints on dietary scope in small and moderate. small mammals:how much do we really understand? In:Chivers D J,Langer Ped.The Digestive System in Mammals:Food,Form and Function[M]. Cambridge:Candaridge University Press,1994.
[27]ZHAN X M,LI Y L,WANG D H.Effects of fasting and refeeding on body mass,thermogenesis and serum leptin in Brandt’s voles(Lasiopodomys brandtii)[J].Journal of Thermal Biology,2009,34(5):237-243.
[28]FRIEDMAN J M,HALLAS J I.Leptin and the regulation of body weight in mammals[J].Nature,1998,395(6704):763-770.
[29]WOODS S C,SEELEY R J,PORTE J D,et al.Signals that regulate food intake and energy homeostasis[J]. Science,1998,280(5368):1378-1383.
[30]BARTNESS T J.Food hoarding is increased by pregnancy,lactation,and food deprivation in Siberian hamsters[J]. American Journal of Physiology,1997,272(1Pt2):R118-R125.
[31]SCHNEIDER J E,BLUM R M,WADE G N.Metabolic control of food intake and estrous cycles in syrian hamsters.I.Plasma insulin and leptin[J].American Journal of Physiology,2000,278(2):R476-R485.
[32]DAY D E,BARTNESS T J.Fasting-induced increases in food hoarding are dependent on the foraging-effort level[J].Physiology & Behavior,2003,78(4-5):655-668.
[33]DAY D E,MINTZ E M,BARTNESS T J.Diet self-selection and food hoarding after food deprivation by Siberian hamsters[J].Physiology & Behavior,1999,68(1-2):187-194.
[34]杨盛昌,王玫昆,杨晓楠,等.瘦素抵抗在妊娠期大绒鼠体重调节中的作用[J].云南师范大学学报:自然科学版,2013,33(5):59-63.
[35]余婷婷,杨盛昌,朱万龙,等.高山姬鼠瘦素抵抗模式的初步研究[J].云南师范大学学报:自然科学版,2014,34(1):75-78.
