稻瘟病菌中己糖载体蛋白的生物信息学分析
2014-06-27许俊洁郑凯玲鲁国东福建农林大学植物保护学院福建福州350002
许俊洁, 陈 键, 郑凯玲, 鲁国东(福建农林大学植物保护学院, 福建 福州 350002)
1 引言
己糖载体蛋白的功能在许多生物中已经有报道,其中,哺乳类动物体内发现了13种己糖转运蛋白(Rogersetal., 2003)。在人体内,葡萄糖能调控基因转录、酶活,激素分泌和受葡萄糖调节的神经细胞的活性(Thorens1etal., 2012;Wetal., 1993)。糖感应途径和糖信号途径如果不能正常调控,则会引发代谢的严重无序(Kimetal., 2013)。但是,细胞是如何感受并转运葡萄糖的,目前这种机制尚不清楚(Karhumaaetal., 2010)。酿酒酵母(Saccharomycescerevisiae)的己糖载体蛋白研究较为深入,已鉴定出20个己糖载体蛋白(Kimetal., 2013;Dietzeletal., 2012;Leandroetal., 2009)。Reifenberger 等通过构建酿酒酵母hxt1-7缺陷突变体,发现此突变菌株在葡萄糖、果糖、甘露醇和蔗糖培养基上不能生长,而将基因Hxt1,Hxt2,Hxt3,Hxt4,Hxt6,Hxt7和Gal2分别互补hxt1-7缺失突变菌株均能在含有葡萄糖的培养基上生长(Maieretal., 2002)。研究表明,酿酒酵母中葡萄糖的传感、吸收调节及葡萄糖利用率之间有着复杂的网络结构(Liuetal., 2013 )。此外,在Neurosporacrassa,Aspergillusnidulans和Colletrotrichumgraminicola中也都鉴定出数个己糖载体蛋白基因(Madietal.,1997;Lingneretal., 2011;Reisetal., 2013;Lietal., 2014 )。
稻瘟病是世界性重大病害(Klausetal., 2012),研究稻瘟病菌的致病机理有着重要意义。目前,稻瘟病菌和水稻互作过程中糖代谢调控途径的研究还很少有报道,尤其是稻瘟病菌摄取糖类的途径和机理目前几乎还是空白(Ebbole,2007)。最近,Hiromasa Saitoh 等首次发现了稻瘟病菌的一个己糖载体蛋白基因MOST1( Saitohetal., 2014)。另有报道称,稻瘟病菌侵染水稻被证明是依赖于稻瘟病菌的6-P葡萄糖的传感器Tps1,它能融合碳源和氮源代谢,并对可利用的葡萄糖做出反应(Reisetal., 2013)。本文将通过生物信息学的方法,对稻瘟病菌中的己糖载体蛋白家族进行初步分析,为进一步研究它们的功能奠定基础。
2 方法
2.1 序列选择和序列比对
为查找稻瘟病菌中可能存在的己糖载体蛋白,利用该载体蛋白的保守结构域Sugar_tr的氨基酸序列搜索稻瘟病菌数据(http:∥www.broadinstitute.org/annotation/genome/magnaporthe_comparative/MultiHome.html)。同时,选择酿酒酵母、构巢曲霉和粗糙脉胞酶已经鉴定功能的部分己糖载体蛋白,并与稻瘟病菌中预测的己糖载体蛋白进行比对分析。相关蛋白序列搜索数据库:构巢曲霉(http:∥www.broadinstitute.org/annotation/genome/aspergillus group/MultiHome.html)粗糙脉胞酶(http:∥www.broadinstitute.org/annotation/genome/neurospora/MultiHome.html)酿酒酵母(http:∥www.broadinstitute.org/annotation/genome/saccharomyces_cerevisiae)。炭疽病菌数据库(http:∥www.broadinstitute.org/annotation/genome/colletotrichum_group/MultiHome.html)。
2.2 系统进化树分析
根据氨基酸序列,使用DNAMAN软件中的Multiple Alignment分析得到Phylogenetic_Tree。
2.3 跨膜螺旋结构和结构域分析
分别用结构域分析在线软件(http:∥string-db.org/newstring_cgi/show_input_page.plUserId =eNQqcNmAi0rN&sessionId=18b__pDB_cxA)和跨膜结构数据库(http:∥www.cbs. dtu.dk/services/TMHMM/)对预测蛋白进行单独分析。
2.4 基因表达量分析
稻瘟病菌的附着胞形成阶段的基因表达量数据主要来源于(http:∥cogeme.ex.ac.uk/supersage/)。侵染阶段基因表达量来自于实验室之前所做的基因芯片杂交试验。
3 结果
3.1 稻瘟病菌己糖载体蛋白系统发育树的建立
通过Blast搜索比对,在稻瘟病菌数据库中发现sugar_sr家族有67个同源蛋白,推测其为己糖载体蛋白。选择酿酒酵母的糖信号受体蛋白Rgt2、Snf3;构巢曲霉和粗糙脉胞酶中分别已经鉴定为葡萄糖载体的部分蛋白NCU_02582、NCU_04963、NCU_06133、NCU_06138、NCU_10021、hxtC(ANID_10891)、hxtD(ANID_08737)、hxtE (ANID_01797)、ANID_06923 ;炭疽病菌中鉴定出的己糖载体蛋白 CgHXT1、CgHXT2、CgHXT3、CgHXT4、CgHXT5与稻瘟病菌中这67个蛋白一起,共同建立了进化树(图1)。通过进化树分析可以看出,稻瘟病菌己糖载体蛋白与N.crassa、A.nidulans和C.graminicola中已鉴定出的部分己糖载体蛋白有一定的同源性(从42%到74%)。MGG_06203分别与N.crassa的NCU_04963和A.nidulans的葡萄糖载体蛋白hxtC(ANID_10891)、hxtE (ANID_01797)以及C.graminicola的己糖载体蛋白CgHXT1同源最高,其中与CgHXT1同源性可达72%。MoST1与NCU_02582和CgHXT4同源。MGG_15700(MGG_01446.6)分别与NCU_10021、CgHXT3和hxtD(ANID_08737)同源。MGG_00040与CgHXT2高度同源,而与A.nidulans、C.graminicola个别蛋白有较低的同源性。MGG_13651只与葡萄糖载体蛋白ANID_06923有43%的同源性。MGG_05946与NCU_06138同源性最高达74%。稻瘟病菌中没有与酿酒酵母中的葡萄糖信号受体Rgt2和Snf3高度同源的蛋白,但是有几个蛋白与其有一定的同源性(39%)。

图1稻瘟病菌中67个己糖载体蛋白与酿酒酵母、N.crassa、A.nidulans
和C.graminicola中已知功能的部分己糖载体蛋白的系统进化关系
Figure 1 The phylogenetic relationship of 67 hexose transporter inM.oryzaeand some identified hexose transporters inS.cerevisiae,N.crassa,A.nidulansandC.graminicola
3.2 稻瘟病菌己糖载体蛋白跨膜结构的分析
通过在线软件TMHMM Server v. 2.0对预测的67个蛋白进行跨膜分析,发现它们在蛋白质结构上存在一定的差异。根据蛋白质的跨膜结构螺旋分为6类,其中MGG_03620等26个蛋白具有12个跨膜结构螺旋,图2所示的为MGG_00040的12个跨膜结构,前100个氨基酸残基构成2个螺旋结构,100到200的氨基酸位点之间有4个螺旋结构,氨基酸位点从200到300之间无螺旋结构,而剩下的约200个氨基酸位点构成了6个跨膜螺旋结构,这些峰值的准确性约为90%以上。MGG_02783 等12个蛋白具有11个跨膜结构螺旋;MGG_00346等22个蛋白具有10个的跨膜结构螺旋;4个蛋白有9个跨膜结构螺旋; MGG_07546只有8个跨膜结构螺旋;而最为特殊的是MGG_07844有16个跨膜结构螺旋结构。不具有12个跨膜结构的蛋白,不代表其预测的跨膜结构不足12个,而是有些跨膜螺旋的峰值的可能性低于80%;当然,也有部分蛋白预测的跨膜螺旋没有12个。根据Maria Jos′e Leandro等预测分析,酵母中的己糖载体蛋白的12层跨膜结构模型见图3。它们的跨膜螺旋结构镶嵌在膜上, 蛋白质的C端和N端都伸入到细胞内部。
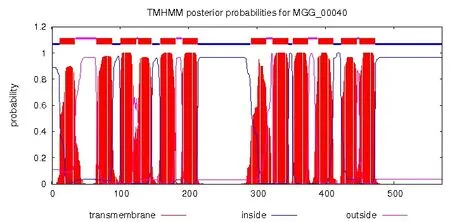
图2 稻瘟病菌基因MGG_00040的12个跨膜螺旋结构图Figure 2 The twelve transmembrane domains of gene MGG_00040 in M.oryzae

图中的5个位点(A-E)分别代表在糖转运蛋白中已经鉴定出的基序 图3 酿酒酵母已糖载体跨膜结构模型Figure 3 Transmembrane model of S.cerevisiae
由结构域在线分析软件STRING 9.1分析预测稻瘟病菌67个蛋白。其中有66个的蛋白由于氨基酸序列的重叠都有两种形式的结构域:Sugar_tr和MFS_1,如图4所示,这类蛋白所具有的此类结构域的数目以及形成的结构域氨基酸位点不同。而MGG_15700蛋白只有一种Sugar_tr式的结构域。另外,较为特殊的是MGG_09307和MGG_13930蛋白各有一个TRI12结构域,此结构域存在于有些致病真菌中,与单端孢毒素的生物合成有关。同时,MGG_13651含有一个DUF791结构域,此结构域存在于真核生物中,但是具体的功能目前未知。

图4基因MGG_03620的两种不同形式的结构域
Figure 4 Two different forms of domains in geneMGG_03620
3.3 稻瘟病菌己糖载体蛋白基因的表达分析
3.3.1 稻瘟病菌附着胞形成过程中己糖载体蛋白基因的表达分析 预测的67个蛋白中,可以从数据库中搜索到稻瘟病菌菌株Guy11对应的63个蛋白在附着胞形成过程中的表达量数据。根据Xu等研究,稻瘟病菌分生孢子着落在寄主植物表面,30min内会萌发形成芽管,8h后大部分细胞会分化形成附着胞,24 h后,90%以上的分生孢子会形成带有芽管的成熟分化的附着胞,之后产生侵染钉侵入植物细胞(Xuetal., 1996)。因此,附着胞的形成对于稻瘟病菌是否能成功侵染寄主细胞起着关键作用。文章中选出表达量变化差异性较大同时又具有代表性的14个基因进行分析。其中,MGG_15700的表达量极高,在基本培养基MM上生长,其表达量达到5033.34。相反,基因MGG_09307的表达量为0,而在CM培养基上也只有1.965。基因MGG_00040和MGG_03360的表达量在附着胞形成过程中不断升高,分别从9.61和74.61升高到72.24和186.37。基因MGG_03620、MGG_06203从4 h到6 h表达量有升高,而后,表达量持续下降。而其他基因表达量不规律,尤其在6 h、8 h和14 h之间波动较大(表1)。
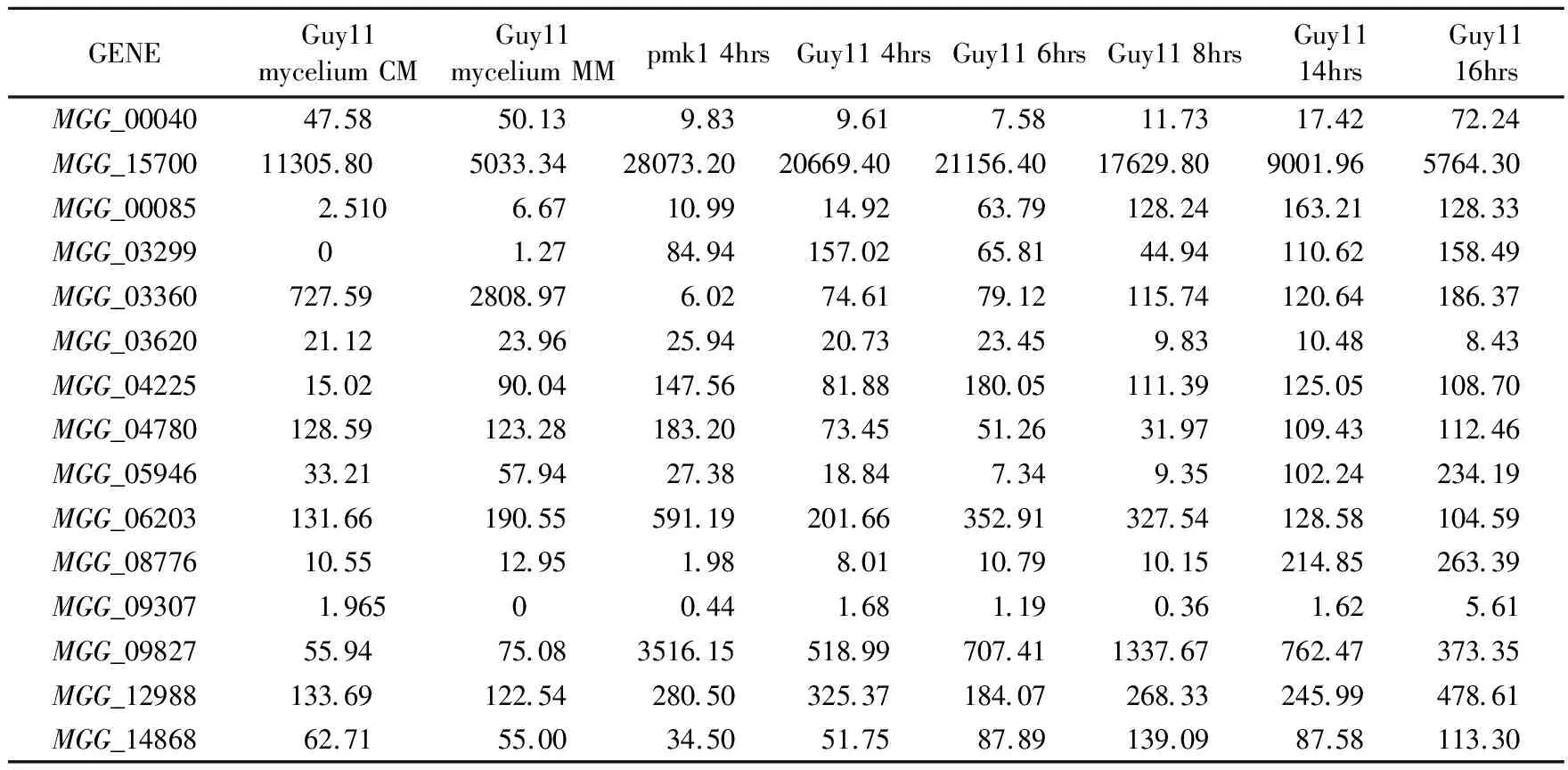
表 1 稻瘟病菌己糖载体蛋白基因在附着胞形成过程中不同时段的表达量Table 1 The hexose transporter genes expression in M.oryzae during appresorium develpment
3.3.2 稻瘟病菌侵染水稻48h后的己糖载体蛋白基因的表达分析 预测的67个己糖载体蛋白中,有25个蛋白基因在稻瘟病菌侵染水稻48h后有明显上调表达。其中,基因MGG_09607, MGG_09680, MGG_09852, MGG_10530上调最为明显,表达量倍数超过6;基因MGG_07884, MGG_09307, MGG_10816有上调,但表达量倍数未超过3;其他17个基因表达量倍数处于3倍到6倍之间(图5)。
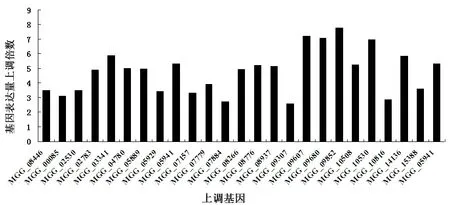
图5 稻瘟病菌己糖载体蛋白基因在侵染阶段的表达量Figure 5 The hexose transporter genes expression in M.oryzae during infection process
4 讨论
本研究通过同源性比对预测稻瘟病菌数据库中有67个己糖载体蛋白,这与Saitoh等的研究一致(Saitohetal., 2014)。稻瘟病菌中,MGG_06203分别与NCU_04963、hxtC(ANID_10891)、hxtE (ANID_01797)、 CgHXT1同源,且与CgHXT1同源性高达72%。MGG_00040与CgHXT2高度同源,而与A. nidulans、C.graminicola的相应己糖载体蛋白有较低的同源性。此外,稻瘟病菌中有的蛋白与酿酒酵母的己糖信号受体有一定的同源性。由此推测,己糖载体蛋白在不同生物中存在特定碱基的保守性。
对67个己糖载体蛋白跨膜结构进行分析,其跨膜螺旋数目分为8、9、10、11、12、16共六个类型。其中,有些蛋白存在更多的跨膜螺旋预测峰形,只是由于峰值的低于80%,不算在其中。采用软件STRING9.1进行在线分析,预测的67个蛋白都具有Sugar_tr结构域,此结构域为己糖载体蛋白的特征结构域。
基因MGG_15700、MGG_09307分别在附着胞形成初期和整个阶段表达量极高;MGG_00040和MGG_03360表达量呈不断升高趋势;MGG_03620和MGG_06203表达量在4 h到6 h表达量有升高,而后持续下降;其它基因的表达量在6 h、8 h和14 h之间波动。总之,根据附着胞形成阶段基因的表达量高低,可以推测其是否参与致病过程,以及在附着胞形成的不同阶段所起作用的差异。
预测的67个己糖载体蛋白中,有42个蛋白的基因在稻瘟病菌侵染水稻的48 h后表达没有明显上调,而另外25个表达上调的基因,可能参与侵染致病过程。目前已经鉴定出的首个稻瘟病菌己糖载体蛋白基因MGG_03620(MoST1)在侵染阶段表达量不高,表明它可能不参与侵染致病过程,这与Hiromasa Saitoh等的研究一致,即此基因的敲除不会影响病菌致病性的变化( Saitohetal., 2014)。当然,上述预测的己糖载体蛋白的生物学和生理生化功能还需要通过实验进行验证。
Couch BC,Fudal I,Lebrun MH,Tharreau D,Valent B,Kim P v,Notteghem JL,Kohn LM.2005.Origins of host-specific populations of the blast pathogenMagnaportheoryzaein crop domestication with subsequent expansion of pandemic clones on rice and weeds of rice.Genetics,170:613-630.
Couch BC,Fudal I,Lebrun MH,Tharreau D,Ebbole DJ.2007.Magnaportheas a model for understanding the host-pathogen interactions.Phytopathol,45:437-56.
Dietzel KL,Ramakrishnan V,Murphy EE,Bisson LF.2012.MTH1 and RGT1 demonstrate combined haploinsufficiency in regulation of the hexose transporter genes inSaccharomycescerevisiae.BMC Genet,13:107.
Fernandez J,Yang KT,Cornwell KM,Wright JD,Wilson RA.2013.Growth in rice cells requires denovo purine biosynthesis by the blast fungusMagnaportheoryzae.Science Report,3:1-9.
Gould GW,Holman GD.1993.The glucose transporter family: structure,function and tissue-specific expression.Biochem,295:329-341.
Karhumaa K,Wu BQ,Kielland-Brandt M C.2010.Conditions with high intracellular glucose inhibit sensing through glucose sensor Snf3 inSaccharomycescerevisiae.J.Cell Biochem,110:920-925.
Scheuermann KK,Raimondi JV,Marschalek R,Andrade AD,Wickert E.2012.Magnaportheoryzaegenetic diversity and its outcomes on the search for durable resistance.The Molecular Basis of Plant Genetic Diversity,InTech Press,
Kim JH,Roy A,Jouandot D.2013.The glucose signaling network in yeast.Biochimica et Biophysica Acta,1830:5204-5210.
Leandro MJ,Fonseca C,Gonçalves P.2009.Hexose and pentose transport in ascomycetous yeasts:an overview.FEMS Yeast Research,9:511-525.
Madi L,McBride SA,Bailey LA,Ebbole DJ.1997.rco-3,a Gene Involved in glucose transport and conidiation inNeurosporacrassa.Genetics,146:499-508.
Li JG,Lin LC,Li HY,Tian CG,Ma YH.2014.Transcriptional comparison of the filamentous fungusNeurosporacrassagrowing on three major monosaccharides D-glucose,D-xylose and L-arabinose .Biotechnology for Biofuels.,7:31.
Lingner U,Münch S,Sode B,Deising .B Sauer N.2011.Functional Characterization of a Eukaryotic Melibiose Transporter.Plant Physiol.,156:1565-1576.
Liu,TB,Wang YN,Xue CY.2013.The Glucose Sensor-Like Protein Hxs1 Is a High-Affinity Glucose Transporter and Required for Virulence in Cryptococcus neoformans.PLoS One,8:1-24.
Maier A,Volker B,Boles E.2002.Characterisation of glucose transport in Saccharomyces cerevisiae with plasma membrane vesicles (countertransport) and intact cells (initial uptake) with single Hxt1,Hxt2,Hxt3,Hxt4,Hxt6,Hxt7 or Gal2 transporters.FEMS Yeast Research,539-550.
Reis TF,Menino JF,Bom VLP,Brown NA,Colabardini AC.2013.Identification of glucose transporters inAspergillusnidulans.PLoS ONE,11:1-15.
Rogers S,Chandler JD,Clarke AL.2003.Glucose transporter GLUT12-functional characterization inXenopuslaevisoocytes Biochem.Biophys.Res.Commun,308:422-426.
Saitoh H,Hirabuchi A,Fujisawa S.2014.MoST1 encoding a hexose transporter-like protein is involved in both conidiation and mycelial melanization ofMagnaportheoryzae.FEMS Microbiology,352:104-113.
Thorens B,Mueckler M.2012.Glucose transporters in the 21st Century.Physiol Endocrinol Metab,298:E141-E145.
Xu JR,Hamer JE.1996.MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungusMagnaporthegrisea.Genes & Development,10:2696-2706.
Xue YK,Shui GH,Wenk MR.2014.TPS1 drug design for rice blast disease inMagnaportheoryzae.Springer Plus,3:18.
