石蒜Mg2+转运体基因LrMGT的克隆与分析
2014-05-11蔡黎丽李晓丹
汪 仁,蔡黎丽,徐 晟,李晓丹,夏 冰
〔江苏省·中国科学院植物研究所(南京中山植物园),江苏南京210014〕
石蒜Mg2+转运体基因LrMGT的克隆与分析
汪 仁,蔡黎丽,徐 晟,李晓丹,夏 冰①
〔江苏省·中国科学院植物研究所(南京中山植物园),江苏南京210014〕
从石蒜〔Lycoris radiata(L’Hér.)Herb.〕叶片全长cDNA文库中克隆获得Mg2+转运体(MGT)基因LrMGT。序列分析结果显示:LrMGT基因的cDNA序列全长1 726 bp,其中开放阅读框(ORF)长度921 bp,编码306个氨基酸。石蒜LrMGT基因编码的氨基酸序列的理论相对分子质量为33 635,理论等电点为pI 5.14,为疏水性膜蛋白,不具有信号肽。序列比对结果表明:石蒜LrMGT基因编码的氨基酸序列与小米〔Setaria italica(Linn.)Beauv.〕、水稻(Oryza sativa Linn.)和拟南芥〔Arabidopsis thaliana(Linn.)Heynh.〕等植物的MGT基因编码的氨基酸序列的相似性较高,相似度达到72%~76%;石蒜LrMGT基因与其他植物MGT基因编码的氨基酸序列的保守区域较大,均具有较高的保守性。在NJ系统树上石蒜LrMGT基因编码的氨基酸序列与禾本科(Gramineae)植物二穗短柄草〔Brachypodium distachyum(Linn.)Beauv.〕、水稻、高粱〔Sorghum bicolor(Linn.)Moench〕和小米MGT基因编码的氨基酸序列聚为同一个分支,表明它们可能具有较近的进化关系。实时荧光定量PCR结果表明:石蒜LrMGT基因在根和鳞茎中的相对表达量较高,在叶片和花中的相对表达量较低,具有明显的组织特异性。
石蒜;Mg2+转运体;基因克隆;序列分析;同源性;表达特性
Mg2+是高等植物细胞中最丰富的二价阳离子,也是植物生长发育必需的核心营养元素之一,参与各种生理生化反应,例如,Mg2+是叶绿素的组成成分,可促进光合作用的碳同化[1];参与维持类囊体膜两侧的电荷平衡[2];调节叶绿体不同光系统之间激发能的分配;维持叶绿体结构的稳定等[3]。此外,Mg2+不仅可以作为ATP的辅助因子,还是细胞内许多酶(包括RNA聚合酶、磷酸化酶、激酶及羧化酶等)的活化剂,调节酶活性;Mg2+还参与跨膜电子梯度的建立,以维持细胞内的渗透势、稳定细胞膜[4]。近年来的研究结果还显示:Mg2+可调控叶绿体mRNA的稳定性[5],并在植物抗病[6]和抗金属Al胁迫中起作用[7-8]。
为了适应土壤中Mg2+含量的变化以及应付其他形式的胁迫,植物体必须严格控制Mg2+的转运。在生物界中已知结构类型并各具特点的Mg2+转运体(MGT)家族主要包括5大类:Mg2+/H+交换体、钴抗性蛋白家族(CorA,Cobalt resistance A)、P型磷酸酶(P-type phosphatase)、离子通道和Mg2+转运体基因家族(MgtE,magnesium transport E)[9]。高等植物中已鉴定出Mg2+/H+交换体和CorA-like镁离子转运蛋白两类具有Mg2+转运活性的转运蛋白[10-12]。
石蒜〔Lycoris radiata(L’Hér.)Herb.〕是重要的观赏和药用植物,其鳞茎中含有多种生物碱,包括石蒜碱、石蒜胺碱、石蒜伦碱、伪石蒜碱和加兰他敏等,其中,加兰他敏具有强效的可逆乙酰胆碱酯酶(AChE, acetylcholinesterase)抑制活性和神经元烟碱受体构象调节作用,临床上用于治疗阿尔茨海默氏病和小儿麻痹症等[13]。近年来,对石蒜属(Lycoris Herb.)植物功能基因的研究取得了一定进展,已对多个功能基因进行了克隆、测序及表达特征的分析和验证[14-20]。在前期工作的基础上,作者从石蒜全长cDNA文库中克隆获得1个Mg2+转运体基因,并利用生物信息学对该基因的核苷酸序列及其编码的氨基酸序列进行分析,以期为石蒜功能基因研究提供更全面的实验依据,并为更加全面了解石蒜MGT基因(LrMGT)的生物学功能奠定理论基础。
1 材料和方法
1.1 材料
供试的石蒜新鲜叶片采自江苏省·中国科学院植物研究所苗圃,石蒜叶片全长cDNA文库为本实验室保存[21]。
植物总RNA提取试剂Trizol reagent购自美国Invitrogen公司;Taq DNA聚合酶、10×PCR buffer(Mg2+free)、MgCl2、dNTPs mixture、DL2000 DNA marker、pMD18-T载体、T4 DNA连接酶和SYBR Premix Ex Taq试剂盒等购自宝生物工程(大连)有限公司;DNA纯化回收试剂盒购于上海华舜生物工程有限公司;大肠杆菌菌株DH5α为本实验室保存;其他常规试剂均为国产分析纯。
1.2 方法
1.2.1 LrMGT基因的克隆和测序 参考GenBank已公布的水稻(Oryza sativa Linn.)OsMGT(登录号NM_ 001051488)、大豆〔Glycine max(Linn.)Merr.〕GmMGT (登录号XM_003526843)、Solanum lycopersicum Linn. SlMGT(登录号XM_004250314)、葡萄(Vitis vinifera Linn.)VvMGT(登录号XM_002272566)、二穗短柄草〔Brachypodium distachyum(Linn.)Beauv.〕BdMGT(登录号XM_003564692)、野草莓(Fragaria vesca Linn.) FvMGT(登录号XM_004288586)、小米〔Setaria italica (Linn.)Beauv.〕SiMGT(登录号XM_004970679)以及拟南芥〔Arabidopsis thaliana(Linn.)Heynh.〕AtMGT (登录号XM_004970679)的保守序列设计引物。根据核苷酸序列比对结果获得1对PCR引物PMGT,上游引物序列为5′-AGCTTCATCATCAAGAAGAAGGG-3′、下游引物序列为5′-CCAGACCAATCCTTGAACAT TAT-3′。
取保存于-75℃超低温冰箱中的石蒜叶片全长cDNA文库20 μL,用灭菌LB液体培养基将菌液稀释10倍,均匀涂布在含有30 μg·mL-1氯霉素的LB固体培养基上,于恒温(37℃)条件下倒置培养过夜(或培养16 h)。随机挑取分散均匀的单菌落,利用引物PMGT进行菌落PCR检测。选取阳性克隆交由上海英骏生物技术有限公司进行测序;结合序列比对,获得该克隆的全长cDNA序列。
1.2.2 LrMGT基因的核苷酸序列及其编码的氨基酸序列分析 检测并去除所测序列两端的载体序列(http:∥www.ncbi.nlm.nih.gov/tools/vecscreen/),利用NCBI-ORF Finder软件(http:∥www.ncbi.nlm. nih.gov/projects/gorf/)分析出目标序列的开放阅读框(http:∥www.ncbi.nlm.nih.gov/gorf)。
采用ProtParam(http:∥www.expasy.org/tools/ protparam.html)分析蛋白质的氨基酸组成;利用GenBank数据库的BLASTp及DNAMAN软件进行氨基酸序列比对,相关植物种类的MGT氨基酸序列均来自NCBI数据库;利用DNAMAN软件进行疏水性/亲水性鉴定;利用SignalP软件分析N末端信号肽序列(http:∥www.cbs.dtu.dk/services/SignalP-4.0/);利用TMHMM软件进行LrMGT蛋白跨膜结构预测(http:∥www.cbs.dtu.dk/services/TMHMM-2.0/);利用SOPMA(http:∥npsa-pbil.ibcp.fr/cgi-bin/npsa_ automat.pl?page=npsa_sopma.html)预测氨基酸序列的二级结构;使用MEGA5.0软件中的邻接法(NJ, neighbor-joining)构建系统进化树。
1.2.3 LrMGT基因的组织表达特性分析 参照高纯度总RNA提取试剂盒的操作说明,分别从石蒜成熟植株的根、鳞茎、叶片和花等组织中提取总RNA,并根据DNA消化酶说明书中的方法对总RNA中的DNA进行消化。取1 μg已消化的总RNA,按照反转录说明书的操作步骤、利用Oligo(dT)18和反转录酶MMLV(RNase H-)合成cDNA第1链。根据克隆获得的LrMGT基因序列设计特异引物,上游引物序列为5′-TATAGAAATAGATGAGCGTGTC-3′、下游引物序列为5′-CTTGAAGATGAAGCTGGATC-3′;以石蒜Actin为内参设计表达检测引物,上游引物序列为5′-CATCCCTCAGCACCTTCCAG-3′、下游引物序列为5′-CTGGGATGCAAAAACCGCC-3′。采用SYBR Premix Ex Taq试剂盒进行实时荧光定量PCR反应;相对定量使用参照基因的ΔCt法,表达差异采用2-ΔΔCt表示[22]。
2 结果和分析
2.1 石蒜LrMGT基因全长cDNA序列克隆及分析
根据本实验室构建的石蒜叶片的全长cDNA文库,利用特异引物从中分离出LrMGT基因。通过对阳性克隆测序并去除载体序列,获得了石蒜Mg2+转运体基因LrMGT的序列。该序列已提交GenBank数据库,登录号为KJ651960。
LrMGT基因的序列分析结果表明:石蒜LrMGT基因cDNA序列全长1 726 bp,包括1个长度为921 bp的开放阅读框(ORF,open reading frame),编码306个氨基酸(图1)。ProtParam分析结果表明:LrMGT基因编码的氨基酸序列的理论相对分子质量为33 635,理论等电点为pI 5.14。

图1 石蒜LrMGT基因cDNA序列及其编码的氨基酸序列Fig.1 cDNA sequence of LrMGT gene from Lycoris radiata(L’Hér.)Herb.and its encoded amino acid sequence

图2 石蒜LrMGT基因与其他植物MGT基因编码的氨基酸序列的多重比对结果Fig.2 Result of multiple alignment of amino acid sequences encoded by LrMGT gene from Lycoris radiata(L’Hér.)Herb.and MGT gene from other species
2.2 石蒜LrMGT基因及其他植物MGT基因编码的氨基酸序列比较及同源性分析
BLASTp分析结果显示:石蒜LrMGT基因编码的氨基酸序列属于EmrE超基因家族,与小米、水稻、二穗短柄草、高粱〔Sorghum bicolor(Linn.)Moench〕、葡萄、野草莓、Solanum lycopersicum、鹰嘴豆(Cicer arietinum Linn.)、大豆和拟南芥的MGT基因编码的氨基酸序列相似性较高,相似度达到72%~76%。
将石蒜LrMGT基因编码的氨基酸序列与其他植物MGT基因编码的氨基酸序列进行多重比对(图2),结果显示:石蒜LrMGT基因及其他植物MGT基因编码的氨基酸序列的保守区域均较大,且连续的相同氨基酸数目较多,表明MGT基因编码的氨基酸序列具有高度的保守性。
为进一步分析从石蒜中分离的LrMGT基因编码的氨基酸序列与其他植物MGT基因编码的氨基酸序列的亲缘关系,构建了NJ系统树(图3)。结果表明:石蒜LrMGT基因编码的氨基酸序列与同属于禾本科(Gramineae)的二穗短柄草、水稻、高粱和小米的MGT基因编码的氨基酸序列亲缘关系较近,聚在同一个分支上。
2.3 石蒜LrMGT基因编码的LrMGT蛋白的生物信息学分析
使用ProtParam对石蒜LrMGT基因编码的LrMGT蛋白的一级结构及理化性质进行预测,结果显示:石蒜LrMGT蛋白的分子式为C1558H2427N367O426S18,半衰期(half-life)为30 h,不稳定指数(instability index)为35.29,脂肪族氨基酸指数(aliphatic index)为118.50。
利用DANMAN软件对石蒜LrMGT基因编码的LrMGT蛋白进行疏水性/亲水性分析(图4),结果表明:该蛋白质的第21位缬氨酸(Val)疏水性最强,第74位亮氨酸(Leu)和第13位苯丙氨酸(Phe)的疏水性次之;第286位组氨酸(His)和第287位赖氨酸(Lys)的亲水性最强。总体上看,石蒜LrMGT蛋白属于疏水性蛋白。
二级结构预测结果显示:α螺旋、延伸链和无规则卷曲是LrMGT蛋白中较多的结构元件,还含有少量的β转角。统计结果表明:LrMGT蛋白的二级结构包含43.79%的α螺旋、26.80%的延伸链、21.57%的无规则卷曲和7.84%的β转角。而跨膜结构预测结果(图5)表明:LrMGT蛋白拥有7个跨膜区,分别是12~34、38~60、67~86、101~123、136~158、173~192和199~221,无信号肽。

图3 石蒜LrMGT基因与其他植物MGT基因编码的氨基酸序列的NJ系统树Fig.3 NJ phylogenetic tree of amino acid sequences encoded by LrMGT gene from Lycoris radiata(L’Hér.)Herb.and MGT gene from other species
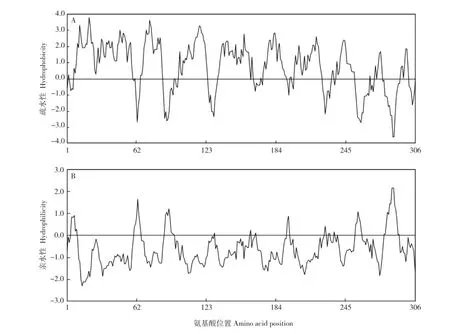
图4 石蒜LrMGT基因编码的LrMGT蛋白的疏水性(A)和亲水性(B)Fig.4 Hydrophobicity(A)and hydrophilicity(B)of LrMGT protein encoded by LrMGT gene from Lycoris radiata(L’Hér.)Herb.

图5 石蒜LrMGT基因编码的LrMGT蛋白的跨膜结构分析Fig.5 Transmembrane structure analysis on LrMGT protein encoded by LrMGT gene from Lycoris radiata(L’Hér.)Herb.
2.4 石蒜LrMGT基因的组织表达特性分析
石蒜不同组织中LrMGT基因相对表达量的实时荧光定量PCR检测结果(图6)显示:石蒜LrMGT基因在石蒜根、鳞茎、叶片和花中的相对表达量差异较大。其中,LrMGT基因在石蒜根中的相对表达量最高,其次为鳞茎;而在叶片和花中的相对表达量较低。由此可见,LrMGT基因在石蒜中的表达具有一定的组织特异性。

图6 石蒜不同组织中LrMGT基因相对表达量的比较Fig.6 Comparison on relative expression of LrMGT gene in different tissues of Lycoris radiata(L’Hér.)Herb.
3 讨 论
随着分子生物技术的发展和应用,已经从不同植物中克隆到多个MGT基因,并对其编码的蛋白质的结构、生化和生理功能等进行了研究[2]。作者以前期构建的高质量石蒜叶片全长cDNA文库为基础,克隆获得了石蒜LrMGT基因的全长cDNA序列。石蒜LrMGT基因与其他植物MGT基因编码的氨基酸序列的比对结果表明:LrMGT基因编码的氨基酸序列与小米、水稻和拟南芥等植物MGT基因编码的氨基酸序列的同源性较高,相似性达到72%~76%,说明MGT基因序列在物种间具有保守性。另外,在NJ系统树上,石蒜与禾本科植物二穗短柄草、水稻、高粱和小米聚为同一个分支,在进化上具有相对较近的关系,说明石蒜与禾本科植物在进化上可能具有一定的关系。通过实时荧光定量PCR分析,石蒜根、鳞茎、叶片和花中LrGMT基因的相对表达量有明显差异,根中LrGMT基因的相对表达量最高、鳞茎中次之、花中最低。可见,该基因的表达具有较明显的组织特异性。
拟南芥Mg2+/H+交换体AtMHX是第1个从多细胞有机体中发现的Mg2+转运蛋白,有11个跨膜区域,长度为100个氨基酸[23];AtMHX定位在液胞膜上,主要存在于木质部薄壁细胞中[2];此外,拟南芥MGT家族(AtMGTs/AtMRS2s)属于CorA类Mg2+转运家族,有10个成员,其氨基酸序列有15%~89%相同,整个家族有6个修饰保守区[11];这些转运体在不同的组织和细胞器中起作用,且大多是膜蛋白。根据这些功能结构特点可将这个大家族分为5个亚家族: AtMGT1和AtMGT2,AtMGT3和AtMGT4,AtMGT5和AtMGT6,AtMGT7、AtMGT8和AtMGT9,AtMGT10[11],其中AtMGT10属于单独的亚家族。在本研究中,膜结构预测结果显示石蒜LrMGT蛋白拥有7个跨膜区,同样属于膜蛋白。因此,LrMGT蛋白究竟属于哪种类型的Mg2+转运蛋白,还需要进一步的验证。植物的MGT蛋白除了具有Mg2+转运功能外,在Al胁迫抗性中也有一定作用[24],而且MGT蛋白还参与了花粉管的发育[25]。因此,石蒜LrMGT基因在植物体内具有的功能还需要进一步研究。
[1] LILLEY R McC,HOLBOROW K,WALKER D A.Magnesium activation of photosynthetic CO2-fixation in a reconstituted chloroplast system[J].New Phytologist,1974,73:657-662.
[2] SHAUL O.Magnesium transport and function in plants:the tip of the iceberg[J].BioMetals,2002,15:309-323.
[3] WILLIAMS L,SALT D E.The plant ionome coming into focus[J]. Current Opinion in Plant Biology,2009,12:247-249.
[4] MAATHUIS F J.Physiological functions of mineral macronutrients [J].Current Opinion in Plant Biology,2009,12:250-258.
[5] HORLITZ M,KLAFF P.Gene-specific trans-regulatory functions of magnesium for chloroplast mRNA stability in higher plants[J].The Journal of Biological Chemistry,2000,275:35638-35645.
[6] HUBER D M,JONES J B.The role of magnesium in plant disease [J].Plant and Soil,2012,368:73-85.
[7] SILVA I R,SMYTH T J,ISRAEL D W,et al.Altered aluminum inhibition of soybean root elongation genotypes in the presence of magnesium[J].Plant and Soil,2001,230:223-230.
[8] YANG J L,YOU J F,LI Y Y,et al.Magnesium enhances aluminum-induced citrate secretion in rice bean roots(Vigna umbellata)by restoring plasma membrane H+-ATPase activity[J]. Plant and Cell Physiology,2007,48:66-73.
[9] 郭照辉,张德元,毛丹丹,等.植物Mg2+转运蛋白与相关基因及其功能的研究进展[J].生命科学研究,2008,12(3):207-210.
[10] SCHOCK I,GREGAN J,STEINHAUSER S,et al.A member of a novel Arabidopsis thaliana gene family of candidate Mg2+ion transporters complements a yeast mitochondrial groupⅡintronsplicing mutant[J].The Plant Journal,2000,24:489-501.
[11] LI L,TUTONE A F,DRUMMOND R S,et al.A novel family of magnesium transport genes in Arabidopsis[J].The Plant Cell, 2001,13:2761-2775.
[12] 丛悦玺,骆东峰,陈坤明,等.生物镁离子转运体研究进展[J].农业生物技术学报,2012,20(7):837-848.
[13] HARVEY A L.The pharmacology of galanthamine and its analogues [J].Pharmacology and Therapeutics,1995,68:113-128.
[14] WANG R,XU S,JIANG Y M,et al.De novo sequence assembly and characterization of Lycoris aurea transcriptome using GS FLX titanium platform of 454 pyrosequencing[J].PLoS One,2013,8: e60449.
[15] JIANG Y M,XIA B,LIANG L J,et al.Molecular and analysis of a phenylalanine ammonia-lyase gene(LrPAL2)from Lycoris radiata [J].Molecular Biology Reports,2013,40:2293-2300.
[16] LI X D,XIA B,WANG R,et al.Molecular cloning and characterization of S-adenosylmethionine synthetase gene from Lycoris radiata[J].Molecular Biology Reports,2013,40:1255-1263.
[17] JIANG Y M,XIA N,LI X D,et al.Molecular cloning and characterization of a phenylalanine ammonia-lyase gene(LrPAL) from Lycoris radiata[J].Molecular Biology Reports,2011,38: 1935-1940.
[18] LI X D,XIA B,JIANG Y M,et al.A new pathogenesis-related protein,LrPR4,from Lycoris radiata,and its antifungal activity against Magnaporthe grisea[J].Molecular Biology Reports,2010, 37:995-1001.
[19] LI Y Z,LI X D,JIANG Y M,et al.Cloning and heterologous expression of a new 3′-hydroxylase gene from Lycoris radiata[J]. Zeitschrift für Naturforschung C,2009,64:138-142.
[20] 梁丽建,江玉梅,夏 冰,等.石蒜儿茶酚氧位甲基转移酶基因克隆与原核表达[J].西北植物学报,2012,32(1):23-28. [21] 王春燕,夏 冰,李晓丹,等.石蒜叶片全长cDNA文库的构建[J].江苏农业学报,2009,25(3):542-546.
[22] 田 畅,蒋 倩,王 枫,等.3个芹菜品种Agnp-G3PDH基因的克隆及其序列和表达特性分析[J].植物资源与环境学报, 2014,23(2):1-10.
[23] SHAUL O,HILGEMANN D W,De-ALMEIDA-ENGLER J,et al. Cloning and characterization of a novel Mg2+/H+exchanger[J]. The EMBO Journal,1999,18:3973-3980.
[24] CHEN Z C,MA J F.Magnesium transporters and their role in Al tolerance in plants[J].Plant and Soil,2013,368:51-56.
[25] MUELLER-ROEBER B,ARVIDSSON S.Fertility control:the role of magnesiumtransportersinpollendevelopment[J].Cell Research,2009,19:800-801.
(责任编辑:张明霞)
Cloning and analysis on Mg2+transporter gene LrMGT from Lycoris radiata WANG Ren,CAI
Lili,XU Sheng,LI Xiaodan,XIA Bing①(Institute of Botany,Jiangsu Province and the Chinese Academy of Sciences,Nanjing 210014,China),J.Plant Resour.&Environ.2014,23(4):1-7
Mg2+transporter(MGT)gene LrMGT was cloned from the full-length cDNA library in leaf of Lycoris radiata(L’Hér.)Herb.Result of sequence analysis shows that the full-length of cDNA sequence of LrMGT gene is 1 726 bp with an open reading frame(ORF)of 921 bp,which encodes 306 amino acids.The theoretical relative molecular weight of the amino acid sequence encoded by LrMGT gene from L.radiata is 33 635 and its theoretical isoelectric point is pI 5.14,and which is hydrophobic membrane protein without signal peptide.The result of sequence alignment shows that amino acid sequence encoded by LrMGT gene from L.radiata possesses higher similarity to those encoded by MGT gene from Setaria italica(Linn.)Beauv.,Oryza sativa Linn.and Arabidopsis thaliana(Linn.)Heynh.,etc,with similarity degree of 72%-76%.There is large conserved region in all of the amino acid sequences encoded by LrMGT gene from L.radiata and MGT gene from other species,which possess high conservation.In NJ phylogenetic tree,the amino acid sequences encoded by LrMGT gene from L.radiata and by MGT gene from other species in Gramineae such as Brachypodium distachyum(Linn.)Beauv.,O.sativa,Sorghum bicolor(Linn.)Moench and S.italica are clustered together,meaning that these species have the closer evolution relationship.The real-time fluorescence quantitative PCR result shows that the relative expression of LrMGT gene from L.radiata is higher in root and bulb and lower in leaf and flower with an obvious tissue specificity.
Lycoris radiata(L’Hér.)Herb.;Mg2+transporter;gene cloning;sequence analysis; homology;expression characteristics
Q785;Q943.2
A
1674-7895(2014)04-0001-07
10.3969/j.issn.1674-7895.2014.04.01
2014-05-04
国家自然科学基金资助项目(31301798);江苏省盐土生物资源研究重点实验室开放课题(JKLBS2012010)
汪 仁(1975—),男,安徽宿松人,博士,副研究员,主要从事植物栽培生理等方面的研究。
①通信作者E-mail:xiabingnbg@sina.com
