9个秋菊品种叶片脂肪酸组成及其抗寒性评价
2014-03-27王翠丽李永华
王翠丽,李 永,崔 洋,李永华
(河南农业大学 林学院,河南 郑州 450002)
温度是影响植物生长、发育及产量的重要环境因子。植物遭受低温胁迫时,生物膜系统最先受到伤害。植物的抗寒性与膜脂的组分和结构有关[1-2],与质膜中不饱和脂肪酸含量的关系更为密切。一般认为,膜脂不饱和脂肪酸含量越高,脂肪酸不饱和指数(Index of Unsaturated Fatty Acid,IUFA)越大,植物的抗寒性就会越强,膜脂不饱和脂肪酸含量及不饱和脂肪酸与饱和脂肪酸的比值(Unsaturated/Saturated Fatty Acid Ratio,USFA/SFA)与植物的抗寒能力呈正相关。在我国北方地区,抗寒性是多年生宿根草本植物的重要性状。秋菊作为一种宿根草本花卉,在园林应用中经常遭受低温伤害,极大地影响了其观赏和经济价值,因此秋菊抗寒性及抗寒品种的选育就成为当前研究的重要方向。郑路等[3]比较了菊花抗寒性与营养特性的关系;Anderson等[4]通过田间统计秋季菊花脚芽的数量,确定了不同菊花品种的抗寒性;Kim等[5]以离体脚芽和植株为材料,通过2种低温处理方法确定了菊花抗寒性的评价方法;许瑛等[6-7]则比较了8个菊花品种的低温半致死温度及其抗寒性,并建立了可靠的菊花耐寒性评价体系。本项目组前期研究了干旱胁迫对不同品种菊花叶片光合生理特性的影响[8]、水杨酸对菊花低温胁迫抗性的影响[9],并探讨了低温胁迫下秋菊膜脂脂肪酸在叶片和根系中抗性机制的差异[10]。但目前的研究多集中在菊花抗寒生理方面,对低温胁迫下秋菊膜脂脂肪酸组分及利用脂肪酸评价抗寒性的研究较少。本研究选用9个秋菊品种为试验材料,通过气相色谱法对其脂肪酸组分与含量进行测定,确定了9个秋菊品种的半致死温度(Lethal Temperature,LT50),并从脂肪酸角度对秋菊的抗寒能力进行研究,以期为菊花抗寒品种的筛选及抗寒性评价提供理论依据。
1 材料与方法
1.1 材 料
供试材料选取菊展和园林绿化中常用的9个秋菊品种,其中包括早花品种(10月上、中旬开花):‘檀香狮子’、‘日出东方’、‘铜雀春深’、‘早粉盘’、‘金锋铃’;晚花品种(11月下旬开花):‘关东新侠’、‘云龙凤舞’、‘星光灿烂’、‘墨宝’。供试秋菊均由开封市园林菊花研究所提供。
1.2 试验设计
将供试材料分为2组,其中一组用于盆栽试验,盆栽基质为V(草炭)∶V(蛭石)∶V(珍珠岩)=1∶1∶1。将生长健壮的菊花幼苗置于人工气候室中培养,温度(25±2) ℃,空气湿度(50±5)%,光照强度(36±0.36) μmol/(m2·s),光照时间10 h/d,常规管理。待株高达(20±2) cm、叶片数量达(13±1)片时进行低温处理:16 ℃(秋菊生长的适宜温度)处理1周,然后5 ℃(植株体内出现明显生理响应的温度[10])低温处理1周。在16和5 ℃条件下,取菊花植株顶端下第4~5片叶,用自来水洗净,蒸馏水反复冲洗,滤纸吸干叶片表面的水分,用于脂肪酸组分及含量测定。另一组则是将9个秋菊品种于2012-05露地定植,常规管理,2012-12-11(9月、10月、11月、12月份的平均温度分别为22,17.5,8.5和2 ℃,最低平均温度分别为17,12,4,-3 ℃;秋菊经过一定时间的自然低温后,抗寒性能得以充分显现,所以于12月份进行采样)取植株顶端下第4~5片叶,测定相对电导率,用于LT50的确定。试验重复3次。
1.3 方 法
1.3.1 脂肪酸的测定 将叶片样品置105 ℃烘箱中杀青5 min,50 ℃恒温烘干。准确称取叶片0.2 g,置100 mL干燥具塞磨口锥形瓶中,加入新配制的体积分数10%的硫酸甲醇溶液10 mL,置70 ℃水浴锅中加热30 min(水面以稍高于瓶中样品液面为宜),分离残渣后,取上清液,移入分液漏斗中,加入20 mL蒸馏水轻轻摇匀,用5 mL无水乙醚萃取。萃取液加适量无水硫酸钠干燥,用0.25 μm微孔滤膜过滤后待用。利用岛津GC2010气相色谱仪测定脂肪酸组分含量,管柱为DB-WAXFTLP毛细管柱(60.0 m×0.53 mm ID)。宽口径进样器(Wide Bore Injector,WBI)温度300 ℃,氢火焰检测器(Flame Ionization Detector,FID)温度350 ℃。载气:氮气,总流量14.4 mL/min,尾吹流量10.0 mL/min,氢气流量40.0 mL/min,空气流量400.0 mL/min。采用脉冲不分流进样,脉冲压109.0 kPa,进样量3 μL。柱温采用程序升温:140 ℃维持2 min,以40 ℃/min升至200 ℃,维持1 min,以5 ℃/min升至220 ℃,维持20 min。以样品保留时间确定不同的脂肪酸组分。采用外标法进行定量计算。
相关指标计算公式如下:饱和脂肪酸(SFA)=肉豆蔻酸(C14∶0)+棕榈酸(C16∶0)+硬脂酸(C18∶0)+山嵛酸(C22∶0);不饱和脂肪酸(USFA)=油酸(C18∶1)+亚油酸(C18∶2)+亚麻酸(C18∶3);脂肪酸不饱和指数(IUFA)=C18∶1+C18∶2×2+C18∶3×3。其中各种脂肪酸的含量为相对含量,即每种脂肪酸占总脂肪酸含量的百分比。
1.3.2 LT50的确定 参照郭海林等[11]的方法,并略做改动测定叶片电导率。每个品种采集生长一致的脚芽叶片,自来水冲洗干净后用去离子水漂洗3次,滤纸吸干。将叶片分成6份,每份约2 g,湿纱布包好置于试管中,4 ℃冰箱中过夜。将样品在低温恒温槽(GDH-1006,scientz公司)中处理,设6个温度梯度:0,-4,-8,-12,-16和-20 ℃,将供试材料在1 h内匀速降温到处理温度,再冷冻1.5 h,处理后置于4 ℃冰箱解冻4 h,将每处理分成3份,每份1 g,置于50 mL三角瓶中,加入去离子水30 mL,在室温下浸提15 h。用电导仪(DDS-307A,上海精密科学仪器有限公司)测定电导率,然后置于沸水浴中15 min,冷却后测定其煮沸电导率。相对电导率(REC)=冷冻电导率/煮沸电导率×100%。
用Logistic方程Y=k/(1+ae-bx)拟合处理温度与相对电导率的关系,其中Y代表细胞伤害率,x代表处理温度,k为细胞伤害率的饱和容量,a、b为方程参数。将处理温度与相对电导率用SPSS统计软件进行非线性回归分析,求得a、b值及相关系数R2,利用公式X=lna/b求出拐点温度X,即为LT50。
1.4 数据分析
试验数据用Microsoft Excel、Word 2003软件进行统计和作图,用SPSS 17.0软件进行方差分析和主成分分析,用SAS 9.2软件进行聚类分析。
2 结果与分析
2.1 叶片脂肪酸组分分析
2.1.1 脂肪酸组分变化 从表1可以看出,菊花叶片中的脂肪酸组分主要有SFA C14∶0、C16∶0、C18∶0、C22∶0和USFA C18∶1、C18∶2、C18∶3。不同品种菊花的脂肪酸组分虽然相同,但其含量存在差异。SFA中以C16∶0为主,16 ℃时其相对含量为11.06%~16.20%,5 ℃时相对含量为9.88%~11.28%。与16 ℃相比,5 ℃条件下除‘檀香狮子’中C14∶0和C18∶0相对含量增加外,其余品种饱和脂肪酸含量均呈下降趋势。USFA中以C18∶3为主,16和5 ℃相对含量分别为51.79%~64.63%,59.99%~64.31%,而C18∶2和C18∶1相对含量分别仅为15.71%~21.84%和2.26%~5.79%。随处理温度的降低,9个秋菊品种叶片中SFA相对含量均呈下降趋势,其中‘关东新侠’下降幅度最大,达35.90%;USFA相对含量均呈上升趋势,其中主要是C18∶3含量增加。
2.1.2 脂肪酸相关指标分析 由表2可见,随着温度的降低,各品种秋菊USFA/SFA与IUFA的变化趋势大致相同,USFA/SFA呈上升趋势,其中‘关东新侠’上升较为明显,上升幅度达70.39%;除‘檀香狮子’外,其余8个秋菊品种的IUFA 值均有不同程度的增加, 5 ℃与16 ℃相比,早花秋菊‘金锋铃’、‘铜雀春深’、‘日出东方’、‘早粉盘’上升幅度分别达7.86%,9.84%,6.23%和13.11%,晚花秋菊‘墨宝’、‘星光灿烂’、‘云龙凤舞’、‘关东新侠’上升幅度分别为6.79%,7.80%,6.28%和10.15%,可见早花秋菊品种的IUFA受温度影响较大。C18∶3/(C18∶1+C18∶2+ C18∶3)明显高于C18∶2/(C18∶1+C18∶2+C18∶3),说明在叶片低温抗性中,C18∶3的作用大于C18∶2。除'檀香狮子'外,其余8个秋菊品种的C18∶3/(C18∶1+C18∶2)和C18∶3/(C18∶1+C18∶2+ C18∶3) 均随温度的降低而呈增加趋势,且C18∶3/(C18∶1+C18∶2)增加幅度较大,最大可达32.51%(‘早粉盘’),而C18∶3/(C18∶1+C18∶2+C18∶3)最大增幅仅为8.96%(‘早粉盘’)。
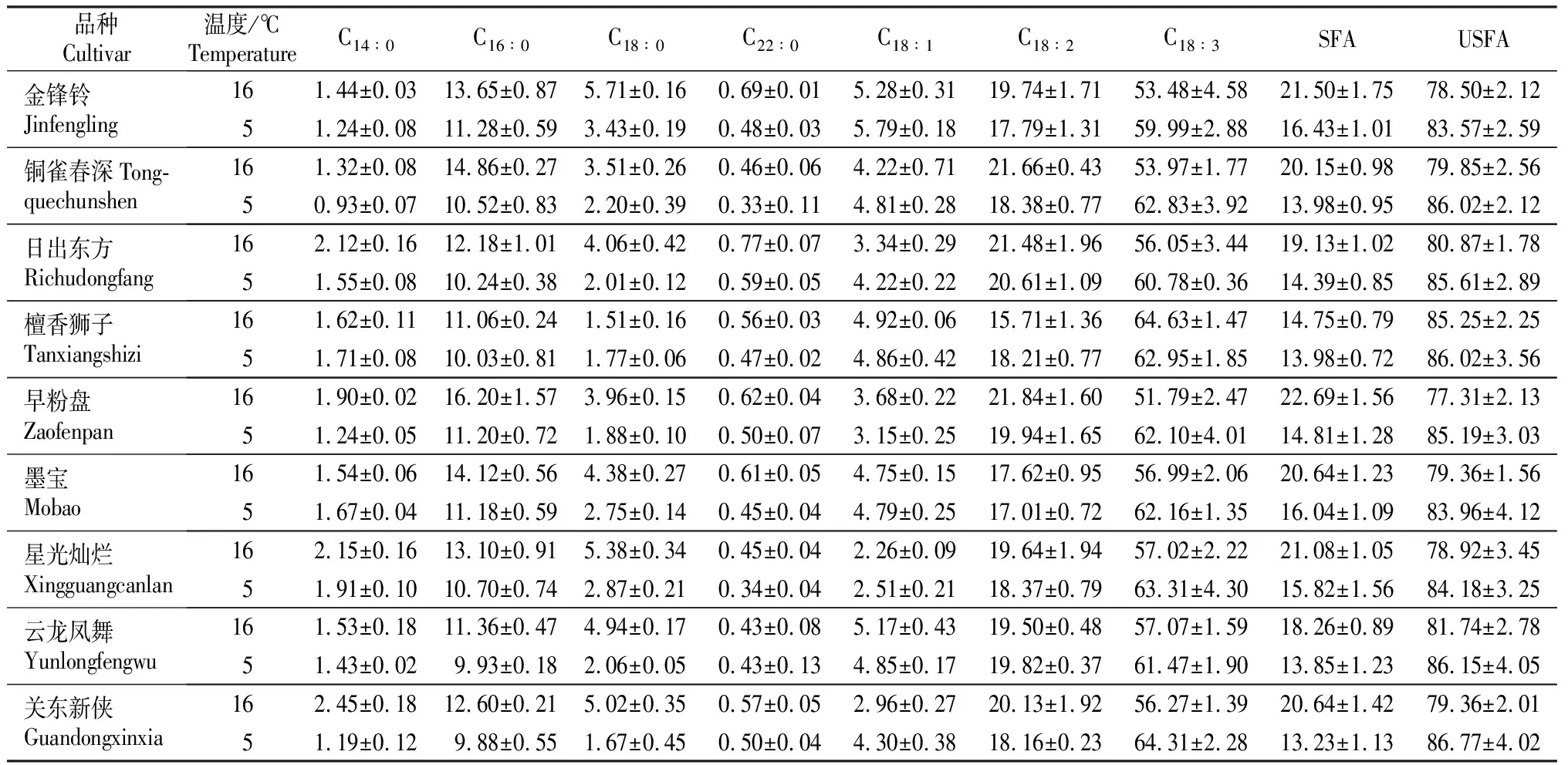
表1 不同温度处理下9个秋菊品种叶片中脂肪酸组分及相对含量

表2 不同温度处理下9个秋菊品种叶片中不饱和脂肪酸相关指标分析
2.2 9个秋菊品种的LT50
以处理温度为横坐标、相对电导率为纵坐标进行作图(以早秋菊‘檀香狮子’和晚秋菊‘云龙凤舞’为代表,其他7个品种与此相似),结果见图1。由图1可知,9个秋菊品种叶片的相对电导率随温度的降低先缓慢增加,当温度降至-4 ℃左右时,相对电导率急剧增加,-12 ℃以后各品种相对电导率增加缓慢。细胞伤害率随处理温度的变化呈典型的“S”型变化。

图1 低温胁迫下秋菊叶片相对电导率与处理温度的关系
由表3可知,处理温度与相对电导率回归方程的相关指数R2均大于0.9,说明回归方程拟合性较好;秋菊品种‘墨宝’、‘星光灿烂’、‘铜雀春深’LT50较低,均在-9 ℃以下,而‘日出东方’的LT50较高,为-6.916 ℃;‘早粉盘’与‘日出东方’、‘墨宝’与‘星光灿烂’之间LT50差异不显著,但其与其他5个品种之间LT50差异均达显著水平。从LT50来看,9个秋菊品种的抗寒能力由高到低依次为‘墨宝’、‘星光灿烂’、‘铜雀春深’、‘金锋铃’、‘檀香狮子’、‘关东新侠’、‘云龙凤舞’、‘早粉盘’、‘日出东方’。

表3 低温胁迫下9个秋菊品种的相对电导率回归方程及半致死温度(LT50)
2.3 9个秋菊品种抗寒性的评价
2.3.1 脂肪酸指标间及其与LT50的关系 对9个与脂肪酸密切相关的生理指标进行相关性分析,结果见表4。由表4可见,C18∶3与IUFA、C18∶3/(C18∶1+C18∶2)与C18∶3/(C18∶1+C18∶2+C18∶3)呈极显著正相关,可共同反映细胞膜系统的变化;C18∶3/(C18∶1+C18∶2+C18∶3)与 C18∶2/(C18∶1+C18∶2+C18∶3)呈极显著负相关;C18∶2与C18∶3呈显著负相关,与IUFA呈显著正相关;C18∶3/(C18∶1+C18∶2)与C18∶2/(C18∶1+C18∶2+C18∶3)呈显著负相关。LT50与USFA/SFA 、C18∶2/(C18∶1+C18∶2+C18∶3)呈正相关,而与C18∶1、C18∶2、C18∶3、 IUFA、C18∶3/(C18∶1+C18∶2)和C18∶3/(C18∶1+C18∶2+C18∶3)均呈负相关。
2.3.2 抗寒性主成分分析 为了全面准确地对各秋菊品种的抗寒性进行评价,对本试验中的C18∶1、C18∶2、C18∶3、USFA/SFA、IUFA、C18∶3/(C18∶1+C18∶2)、C18∶3/(C18∶1+C18∶2+C18∶3)、C18∶2/(C18∶1+C18∶2+C18∶3)等8个指标进行主成分分析。根据特征值大于1的原则(表5、表6),提取了2个主成分,各主成分携带的信息如下:第1主成分有C18∶3、C18∶2/(C18∶1+C18∶2+C18∶3)、C18∶3/(C18∶1+C18∶2)、C18∶3/(C18∶1+C18∶2+C18∶3)、IUFA;第2主成分有C18∶2、USFA/SFA、C18∶1。表7为各菊花品种在2个主成分上的得分,第1主成分得分较高的为‘檀香狮子’、‘墨宝’、‘日出东方’、‘铜雀春深’、‘云龙凤舞’、‘星光灿烂’;第2主成分得分较高的为‘金锋铃’、‘早粉盘’、‘关东新侠’。

表4 低温胁迫下9个秋菊品种脂肪酸指标间及其与LT50的相关系数

表5 9个秋菊品种抗寒性评价的主成分特征值及相应的贡献率

表6 9个秋菊品种抗寒性评价主成分荷载矩阵

表7 9个秋菊品种的主成分得分
2.3.3 秋菊品种聚类分析 通过聚类分析(图2)可见,秋菊品种按抗寒性强弱分为3类:第1类包括‘墨宝’、‘星光灿烂’、‘檀香狮子’,抗寒性较强;第2类包括‘金锋铃’和‘早粉盘’,抗寒性中等;第3类包括‘铜雀春深’、‘云龙凤舞’、‘日出东方’、‘关东新侠’,抗寒性较弱。
3 讨 论
不同植物体内的不饱和脂肪酸组分各异,低温胁迫下其含量对不饱和度的影响也存在较大差异。在油菜籽[12]、马铃薯[13]等的研究中发现,低温可改变质膜脂肪酸的组成,诱导不饱和脂肪酸的含量和比例升高。本试验中,在低温处理条件下,9个秋菊叶片中的不饱和脂肪酸含量均增加,这主要是由亚麻酸含量增加引起;同时,饱和脂肪酸含量降低,棕榈酸含量决定着饱和脂肪酸的含量变化,表明膜脂脂肪酸组分随温度的降低进行了适应性的调整,脂肪酸去饱和化,不饱和脂肪酸含量增加,从而改善了低温环境下秋菊细胞膜的流动性。

图2 低温胁迫下9个秋菊品种树型聚类分析图
在将不饱和脂肪酸作为抗寒指标方面,尹田夫等[14]通过对大豆叶片线粒体膜进行研究,确定C16∶0和C18∶3为抗寒指示性脂肪酸;刘星辉等[15]对不同耐寒性的龙眼、荔枝品种的研究表明,其叶片中的膜脂脂肪酸组分含量存在较大差异,C18∶3/(C18∶1+C18∶2)差异显著,可作为鉴别品种间耐寒性差异的有效指标;夏明等[16]通过对低温下苜蓿叶片膜脂脂肪酸组分的研究,确定C16∶0和C18∶3为苜蓿的抗寒指标性脂肪酸。本项目组前期试验对4个秋菊品种的脂肪酸含量进行了分析,确定秋菊叶片中C18∶3/(C18∶1+C18∶2),根系中C18∶2/(C18∶1+C18∶2+C18∶3)可以作为检验菊花抗寒性的有效指标[10]。本试验中,低温处理下秋菊叶片中的C16∶0、C18∶2、C18∶3等脂肪酸组分含量变化较大,C18∶3/(C18∶1+C18∶2)变化显著,可作为鉴别秋菊品种耐寒性的有效指标,再次验证了本项目组以前的研究结果。就植物的抗寒性而言,其大小不仅取决于其不饱和脂肪酸的含量,还要考虑在低温适应过程中,植物产生的一系列包括不饱和脂肪酸合成和分解在内的综合生理变化[17]。
为了全面准确地利用各种指标对植物的抗寒性进行综合评价,克服单指标鉴定的不足,本研究通过聚类分析将秋菊品种按抗寒性强弱分为3类:第1类抗寒性较强,包括‘墨宝’、‘星光灿烂’、‘檀香狮子’;第2类抗寒性中等,包括‘金锋铃’和‘早粉盘’;第3类抗寒性较弱,包括‘铜雀春深’、‘云龙凤舞’、‘日出东方’、‘关东新侠’,这一结果与根据LT50得出的结果稍有不同。通过LT50和聚类分析发现,晚花品种‘墨宝’、‘星光灿烂’抗寒性较强,但也存在早花品种抗寒性较强、晚花品种抗寒性较弱的现象,花期与菊花的抗寒能力存在一定的关系,这可能与品种特性有关。许瑛等[6]认为,菊花植株的抗寒性与花期的相关性不显著,这种情况还有待进一步探讨。张军科等[18]采用主成分分析法发现,LT50可以代表植物生理指标的绝大部分信息,从而证实了LT50可以作为快速鉴定的单因素指标,该结果具有较好的理论参考价值。在复杂的低温响应机制中,植物的脂类物质代谢具有重要作用,而作为不饱和脂肪酸重要组分的C18∶3对低温环境更加敏感。现代生物技术和植物抗寒工程技术的有机结合,将有助于深入理解低温条件下脂肪酸代谢的作用机制,但不饱和脂肪酸合成途径中其他相关基因的调控机理还有待进一步研究。
[参考文献]
[1] Nishida I,Murata N.Chilling sensitivity in plants and cynobacteria:The crucial contribution of membrane lipids [J].Plant Mol Biol,1996,47:541-568.
[2] Murata N,Los D A.Membrane fluidity and temperature perception [J].Plant Physiol,1997,115(3):875-879.
[3] 郑 路,傅玉兰,陈树桃,等.菊花抗寒性与营养特性的研究 [J].园艺学报,1994,21(2):185-188.
Zheng L,Fu Y L,Chen S T,et al.Studies on the cold resistance and nutrition characteristic of chrysanthemum [J].Acta Horticulturae Sinica,1994,21(2):185-188.(in Chinese)
[4] Anderson N,Gesick E.Phenotypic markers for selection of wi-nter hardy garden chrysanthemum (Dendranthema×grandifloraTzvelv.) genotypes [J].Scientia Horticulturae,2004,101(1):153-167.
[5] Kim D C,Anderson N O.Comparative analysis of laboratory freezing methods to establish cold tolerance of detached rhizomes and intact crowns in garden chrysanthemums (Dendranthema×grandifloraTzvelv.) [J].Scientia Horticulturae,2006,109(4):345-352.
[6] 许 瑛,陈发棣.菊花8个品种的低温半致死温度及其抗寒适应性 [J].园艺学报,2008,35(4):559-564.
Xu Y,Chen F D.The LT50and cold tolerance adaptability of chrysanthemum during a natural drop in temperature [J].Acta Horticulturae Sinica,2008,35(4):559-564.(in Chinese)
[7] 许 瑛,陈 煜,陈发棣,等.菊花耐寒特性分析及其评价指标的确定 [J].中国农业科学,2009,42(3):974-981.
Xu Y,Chen Y,Chen F D,et al.Analysis of cold-tolerance and determination of cold-tolerance evaluation indicators in chrysanthemum [J].Scientia Agricultura Sinica,2009,42(3):974-981.(in Chinese)
[8] 孔德政,于红芳,李永华,等.干旱胁迫对不同品种菊花叶片光合生理特性的影响 [J].西北农林科技大学学报:自然科学版,2010,38(11):103-108.
Kong D Z,Yu H F,Li Y H,et al.Effect of drought stress on photosynthesis and physiologica characteristic of chrysanthemum morifolium [J].Journal of Northwest A&F University:Nat Sci Ed,2010,38(11):103-108.(in Chinese)
[9] 李永华,苏志国,李洪涛,等.水杨酸预处理对低温胁迫下菊花叶片生理活性的影响 [J].河南农业科学,2012(12):99-101.
Li Y H,Su Z G,Li H T,et al.Effects of salicylic acid pretreatment on physiological activity of chrysanthemum leaves under low temperature stress [J].Scientia Agricultura Henan,2012(12):99-101.(in Chinese)
[10] 李永华,史春会,李 永,等.低温下4种秋菊叶片和根系膜脂脂肪酸组分比较 [J].植物生理学报,2013,49(5):1-6.
Li Y H,Shi C H,Li Y,et al.Comparison of membrane fatty acids component in leaf and root of four autumn chrysanthemum cultivars under low temperature [J].Plant Physiology Journal,2013,49(5):1-6.(in Chinese)
[11] 郭海林,刘建秀,朱雪花,等.结缕草属杂交后代抗寒性评价 [J].草地学报,2006,14(1):25-27.
Guo H L,Liu J X,Zhu X H,et al.Evaluation of cold resistance of zoysia hybrids [J].Acta Agrestia Sinica,2006,14(1):25-27.(in Chinese)
[12] Tasseva G,Davy de Virville J,Cantrel C,et al.Changes in the endoplasmic reticulum lipid proprieties in response to low temperature inBrassicanapus[J].Plant Physiology and Biochemistry,2004,42(10):811-822.
[13] De Palma M,Grillo S,Massarelli I,et al.Regulation of desaturase gene expression,changes in membrane lipid composition and freezing tolerance in potato plants [J].Molecular Breeding,2008,21:15-26.
[14] 尹田夫,宋英淑,刘丽君,等.干旱对大豆线粒体膜脂的磷脂和脂肪酸组分的影响 [J].植物生理学通讯,1989(4):16-18.
Yin T F,Song Y S,Liu L J,et al.Effects of drought on the constituents of phospholipld and fatty acid in mitochondria membrane liplds of soybean leaves [J].Plant Physiology Communications,1989(4):16-18.(in Chinese)
[15] 刘星辉,佘文琴,张惠斌.龙眼、荔枝叶片膜脂肪酸与抗寒性的研究 [J].福建农业大学学报,1996,25(3):297-301.
Liu X H,She W Q,Zhang H B.Relationship between membrane fatty acid in leaves and cold resistance of longan and litch [J].Journal of Fujian Agricultural University,1996,25(3):297-301.(in Chinese)
[16] 夏 明,刘亚学,阿拉木斯,等.低温下苜蓿叶片膜脂脂肪酸组分的研究 [J].中国草地,2002(6):28-37.
Xia M,Liu Y X,A L M S,et al.Fatty acid composition of cell membrane of alfalfa leaves at chilling temperature [J].Grassland of China,2002(6):28-37.(in Chinese)
[17] 张 玮,谢锦忠,吴继林,等.低温驯化对部分丛生竹种叶片膜脂脂肪酸的影响 [J].林业科学研究,2009,22(1):139-143.
Zhang W,Xie J Z,Wu J L,et al.Changes in membrane lipid fatty acids of some sympodial bamboosin response to low temperature exposure [J].Forest Research,2009,22(1):139-143.(in Chinese)
[18] 张军科,桑春果,李嘉瑞,等.杏品种资源抗寒性主成分分析 [J].西北农业大学学报,1999,27(6):79-84.
Zhang J K,Sang C G,Li J R,et al.Principle component analysis method of almonds tolerance to cold [J].Acta Univ Agric Boreali-Occidentalis,1999,27(6):79-84.(in Chinese)
