Jeju seaweeds suppress lipopolysaccharide-stimulated proinflammatory response in RAW 264.7 murine macrophages
2014-03-23EunJinYangJiYoungMoonSangSukKimKyongWolYangWookJaeLeeNamHoLeeChangGuHyun
Eun-Jin Yang, Ji-Young Moon, Sang Suk Kim, Kyong-Wol Yang, Wook Jae Lee, Nam Ho Lee, Chang-Gu Hyun,5*
1Cosmetic Science Center, Department of Chemistry, Jeju National University, Jeju 690-756, Korea
2Jeju Technopark, Ara-1-dong, Jeju-si, Jeju 690-121, Korea
3Citrus Research Station, National Institute of Horticultural & Herbal Science, RDA, Jeju 699-946, Korea
4Jeju Love Co., Ltd., 542-5 Haengwon-ri, Gujwa-eup, Jeju 695-975, Korea
5LINC Agency, Jeju National University, Ara-1-dong, Jeju 690-756, Korea
Jeju seaweeds suppress lipopolysaccharide-stimulated proinflammatory response in RAW 264.7 murine macrophages
Eun-Jin Yang1,2, Ji-Young Moon1,2, Sang Suk Kim1,3, Kyong-Wol Yang4, Wook Jae Lee2, Nam Ho Lee1, Chang-Gu Hyun1,5*
1Cosmetic Science Center, Department of Chemistry, Jeju National University, Jeju 690-756, Korea
2Jeju Technopark, Ara-1-dong, Jeju-si, Jeju 690-121, Korea
3Citrus Research Station, National Institute of Horticultural & Herbal Science, RDA, Jeju 699-946, Korea
4Jeju Love Co., Ltd., 542-5 Haengwon-ri, Gujwa-eup, Jeju 695-975, Korea
5LINC Agency, Jeju National University, Ara-1-dong, Jeju 690-756, Korea
PEER REVIEW
Peer reviewer
Assistant Prof. Seung-Young KIM, Department of Food Science and Biotechnology, University of Shizuoka, 52-1, Yada, Suruga-ku, Shizuoka, 422-8526, Japan. Tel:+81 54 264-5555, Fax: +81 54 264-5555, E-mail: sykim01@ u-shizuoka-ken.ac.jp
Co-reviewers: Rajnish Sahu, Kragujevac, MS, USA. Kil-Nam Kim, Jeju, Korea. Ji-Kwon Park, Jeju, Korea.
Comments
This is a good study in which the authors evaluated the effect of ethylacetate extracts of Jeju seaweed that are effective against LPS-induced inflammation in murine macrophage cells. The results are interesting and suggest that Jeju seaweeds may be used as alternative therapy for antiinflammatory diseases.
Details on Page 535
Objective:To investigate the anti-inflammatory effects of Jeju seaweeds on macrophage RAW 264.7 cells under lipopolysaccharide (LPS) stimulation.
Nitric oxide, Interleukin-6, Prostaglandin E2, Tumor necrosis factor-α, Seaweeds, Proinflammatory mediators
1. Introduction
The marine ecosystem and organisms supply bioactive and/or secondary metabolites with a number of biological and pharmacological activities. Of marine organisms, seaweeds are an important source of protein, iodine,vitamins, and minerals, and secondary metabolites might be promising anti-inflammatory agents[1]. Indeed, seaweed extracts and their natural products are known to possess a wide range of biological functions with diverse health benefits[2-5]. Recently, many studies have confirmed the anti-inflammatory activity of seaweeds. Yang and others reported that an anti-inflammatory effect ofPetalonia binghamiae, brown algae was mediated by the suppression of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2)[6], while Yoon and others reported thatSargassum muticum, also brown algae, inhibited the expression of proinflammatory cytokines, iNOS, and COX-2 in the RAW 264.7 cells[7]. Hwanget al.and Kazłowskaet al.reported the inhibition of lipopolysaccharide (LPS)-induced inflammatory responses of marine sulfated polysaccharide and phenolic compounds[8,9]. Also, several anti-inflammatory natural products such as apo-9’-fucoxanthinone, sargaquinoic acid, and sargachromenol have been discovered and described[10,11]. These compounds are abundant inSargassumsp.; they show anti-inflammatory activity and have various other functions[12].
Inflammation is an essential host response to external stimuli (e.g., microbial infection, endotoxin exposure, or tissue injury). It ultimately releases the complex array of inflammatory mediators, leading to the restoration of normal cell structure and function. However, inflammation is in a way a double-edged sword in that the prolonged inflammation contributes to the development of a number of chronic diseases such as atherosclerosis, rheumatoid arthritis, Alzheimer’s disease, asthma, and allergies[13-17]. In inflammatory processes, macrophages, activated immunocytes, play a crucial role in providing an immediate defense against microbial invasion and tissue injury. On activation with an LPS, a well-studied component from the outer membrane of Gram-negative bacteria, macrophages produces proinflammatory mediators and cytokines, including nitric oxide (NO), prostaglandin E2(PGE2), tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6[18-20]. Hence, a down-regulation of these mediators and cytokines in macrophages is essential in suppressing chronic inflammatory process and provides a rationale for the development of therapeutic agents that might be used in various inflammatory diseases[21,22].
As part of our continuous effort to develop value-added utilization of seaweeds, the present work aimed to evaluate the anti-inflammatory properties of five kinds of Jeju seaweeds for human benefit. In this study, we investigated whether ethyl acetate (EtOAc) fractions ofDictyopteris divaricate(D. divaricate),Dictyopteris prolifera(D. prolifera),Prionitis cornea(P. cornea),Grateloupia lanceolata(G. lanceolata), andGrateloupia filicina(G. filicina) inhibit NO and PGE2production as well as iNOS and COX-2 protein expression in the LPS-induced murine macrophage system. Our study may provide a molecular basis for the therapeutic use of Jeju seaweeds in various inflammatory diseases.
2. Materials and methods
2.1. Algal material
All seaweeds in this study were collected from the Jeju Island, Jeju Province, South Korea, and identified by Dr. Wook Jae Lee, Jeju Technopark, South Korea. The voucher specimen was deposited in the herbarium of the Jeju Biodiversity Research Institute, Jeju, Korea. The fresh seaweeds were washed in freshwater to remove sediment, epifauna, and epiphytes, and then dried in the air for 2 weeks, ground into fine powder, and stored in plastic bags at 4 °C until further experiment. The dried seaweeds (1 kg) were extracted with 80% ethanol (5 L) at room temperature for 24 h and then evaporated under vacuum. The evaporated seaweed extracts (10 g each) were suspended in water (1 L) and partitioned with EtOAc (1 L; repeated 3 times).
2.2. Cell culture and viability
Murine RAW 264.7 macrophages were obtained from the Korean Cell Line Bank (Seoul, South Korea). Cells were cultured in Dulbecco’s modified Eagle’s medium, supplemented with 10% fetal bovine serum, penicillin, and streptomycin sulfate (all from GIBCO, Grand Island, NY, USA), in an incubator at 37 °C in a humidified atmosphere of 95% air and 5% CO2. For this study the cells were mechanically passaged by dissociation every other day; they underwent fewer than 25 passages. A 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was undertaken as described previously[10]. Cells were counted with a haemocytometer, and the number of viable cells was assessed by trypan blue dye exclusion method. The RAW 264.7 macrophages were seeded in 96-well plates for 18 h and then stimulated with various concentrations (12.5, 25, 50, or 100 µg/mL) of the Jeju seaweeds (D. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicina) or LPS (1 µg/mL) for the indicated time. On the day of collection, cells were incubated with MTT solution for 4 h at 37 °C in a humidified atmosphere of 95% air and 5% CO2. The MTT-containing supernatant was removed and the formazan crystals were solubilized in dimethylsulfoxide. Absorbance of each well at 540 nm was measured using an automatic microplate reader (PowerWave X340, Bio-tech Instruments, Inc., Winooski, VT, USA).
2.3. Determination ofNOconcentration
The NO concentration inD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinaextracts was determined based on the method of Yanget al[10].RAW 264.7 cells (5×105cells/well) were cultured on 24-well dishes with 1 µg/mL of LPS and various concentrations (12.5, 25, 50, or 100 µg/mL) ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicina. After 18 h, culture supernatantswere collected and nitrite, a stable oxidized product of NO, was measured by a modified Griess method. Equal volumes (100 µL) of the Griess reagent [1% sulfanilamide and 0.1%N-(1-naphthyl)-ethylenediamine dihydrochloride in 5% phosphoric acid] andD. divaricate,D. prolifera,P. cornea,G. lanceolata, orG. filicinawere incubated together at room temperature for 10 min. Absorbance of each well at 540 nm was measured using an automatic microplate reader (PowerWave X340, Bio-tech Instruments).
2.4. Measurement ofPGE2and cytokine assays
The inhibitory effect ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinaon the production of PGE2and pro-inflammatory cytokines from the LPS-stimulated RAW 264.7 cells was determined using a previously described method with slight modifications[10]. Macrophage RAW 264.7 cells (1.8×105cells/mL) were cultured in 24-well plates for 18 h, and then treated with LPS (1 µg/ mL) in the presence of various concentrations (12.5, 25, 50, or 100 µg/mL) ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicina. After 24-hour incubation, PGE2levels in the culture medium were quantified using an enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MI, USA). The release of PGE2was measured relative to that of the control value. For cytokine immunoassays, the RAW 264.7 cells (1.8×105cells/mL) were cultured in 12-well plates, incubated with various concentrations (12.5, 25, 50, or 100 µg/mL) ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinain 1 µg/mL LPS for 24 h. Cell supernatants were then collected, and the levels of TNF-α and IL-6 were measured using the ELISA kit (R&D Systems) according to the manufacturer’s instructions.
2.5. Western blot analysis
The western blot analysis was performed to evaluate the effect ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinaon iNOS and COX-2 expression as described previously[10]. The RAW 264.7 cells (2.5×106cells/mL) were cultivated in a 6-well plate for 18 h and then treated with the various concentrations ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinaextracts in the absence or presence of LPS. After treatment for 24 h with LPS, the RAW 264.7 cells were harvested and the total protein was extracted with RIPA lysis buffer (Santa Cruz, CA, USA) supplemented with protease inhibitors, and kept on ice for 30 min. β-actin was used as control. The protein was denatured and separated on sodium dodecyl sulfonate (SDS)-polyacrylamide gels and then transferred onto a polyvinylidene fluoride membrane (Bio-Rad, HC, USA). The membranes were blocked and incubated with iNOS or COX-2 antibodies, followed by incubation with the horseradish peroxidaselinked secondary antibody (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA, USA). The signals were detected using an enhanced chemiluminescence reagent (Amersham Life Sciences, Arlington Heights, IL, USA).
2.6. Analysis of total phlorotannin
The total phlorotannin content ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinawas determined by the Folin-Ciocalteu colorimetric method according to the method of Huanget al[23], as modified by Yanget al[24]. Sample absorbances were read at 725 nm. Phloroglucinol was used as the standard for a calibration curve; the total phlorotannin content of the seaweed extracts was expressed as phloroglucinol equivalents. The analysis of each seaweed extract was conducted in quadruplicate.
2.7. Statistical analysis
All results were expressed as mean±SE. Each experiment was repeated at least 3 times. Statistical significances were compared between each treated group and analyzed by the Student’st-test. AP-value of less than 0.05 was considered statistically significant.
3. Results
3.1. Effect of Jeju seaweed fractions on the viability of theRAW264.7 cells
Examination of cytotoxicity on the RAW 264.7 cells by an MTT assay indicated that none of the Jeju seaweed extracts at a concentration of up to 100 µg/mL affected the viability of the cells after 48-hour incubation (data not shown). Similarly, none of the Jeju seaweed extracts had a significant effect on the viability of the RAW 264.7 cells at various concentrations (up to 100 µg/mL) after 24-hour incubation (Figure 1).
3.2. Inhibition ofLPS-inducedNOproduction by Jeju seaweed fractions
In this study, to identify whether Jeju seaweeds have antiinflammatory activity, we prepared 80% ethanol extracts fromD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinafrom the Jeju Island, South Korea. All five types of evaporated 80% ethanol extracts were partitioned with EtOAc. All EtOAc fractions of the seaweeds were dissolved in 80% ethanol and diluted with sterile water to normalize the concentration of the experimental samples. After treatment with LPS (1 µg/mL) for 24 h, nitrite concentrations in the medium increased remarkably by about six-fold. When the RAW 264.7 cells were treated with 1 µg/mL of LPS and various concentrations (12.5, 25, 50, or 100 µg/mL) of the EtOAc fractions of the Jeju seaweeds for 18 h, a significant (P<0.05) concentration-dependent inhibition of nitrite production was detected in the medium. All EtOAc fractions of the Jeju seaweeds showed more than 50% inhibition of NO production at a concentration of 40 µg/mL in the culture medium. IC50values ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinawere 18.0, 38.36, 38.43, 32.81, and 37.14 µg/mL, respectively.
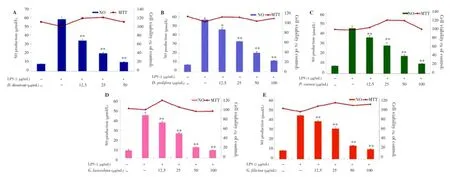
Figure 1. Effect of Jeju seaweed extracts on NO production in LPS-stimulated RAW 264.7 cells.The cells were stimulated with 1 µg/mL of LPS alone or with a combination of LPS and various concentrations (12.5, 25, 50, or 100 µg/mL) of seaweed extracts [D. divaricate (A), D. prolifera (B), P. cornea (C), G. lanceolata (D), and G. filicina (E)] for 24 h. NO production was determined by the Griess reagent method. Cell viability was determined using the 24-hour culture of cells stimulated with LPS (1 µg/mL) in the presence of each sample. The data represent the mean±SD of triplicate experiments. *P<0.05; **P<0.01 vs. LPS alone.
3.3. Inhibition ofLPS-inducedPGE2produced by Jeju seaweed fractions
A large body of data indicated that COX-2 played a key role in several biological processes such as chronic inflammation; therefore, we next examined the effects ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinaextracts on PGE2production in LPS-stimulated RAW 264.7 macrophages. When the macrophages were stimulated with LPS (1 µg/mL) for 24 h, the levels of PGE2increased in the culture medium. As shown in Figure 2, except forG. lanceolata, other samples (D. divaricate,D. prolifera,P. cornea,G. filicina) suppressed the LPS-induced PGE2production in a dose-dependent manner. Among these 4 extracts, 3 extracts--i.e., those fromD. divaricate,D. prolifera, andP. cornea--showed the most potent inhibition with IC50values below of 12.5 µg/mL.
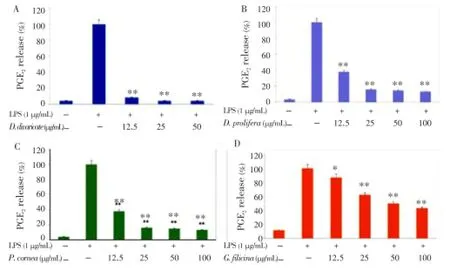
Figure 2. Effect of Jeju seaweed extracts on PGE2production in LPS-stimulated RAW 264.7 cells.The cells were stimulated with 1 µg/mL of LPS alone or with a combination of LPS and various concentrations (12.5, 25, 50, or 100 µg/mL) of D. divaricate (A), D. prolifera (B), P. cornea (C), and G. filicina (D) for 24 h. PGE2produced and released into the culture medium was assayed by the ELISA method. The data represent the mean±SD of triplicate experiments. *P<0.05; **P<0.01 vs. LPS alone.
3.4. Inhibitory effects of Jeju seaweed extracts onLPS-induced expression ofCOX-2 and iNOS
The western blot analysis was performed to determine whether the inhibitory effects of the Jeju seaweed extracts on the proinflammatory mediators, NO and PGE2, were related to the modulation of iNOS and COX-2 expression. As shown in Figures 3 and 4, we did not detect iNOS and COX-2 proteins in non-stimulated RAW 264.7 cells. On the contrary, a significant increase in the levels of COX-2 and iNOS proteins was observed in response to LPS. In addition, treatment with EtOAc fractions ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicina(12.5, 25, 50, or 100 µg/mL) resulted in a dose-dependent inhibition of the levels of LPS-stimulated COX-2 and iNOS proteins. These results were consistent with the inhibitory effects of Jeju seaweeds on PGE2and NO production in LPS-stimulated RAW 264.7 cells.
3.5. Inhibitory effects of Jeju seaweed extracts onLPS-inducedTNF-αandIL-6 production
Considering that all extracts potently inhibited the proinflammatory mediators (PGE2and NO), we continued to investigate whether they suppressed the production of proinflammatory cytokines such as TNF-α and IL-6 in LPS-stimulated RAW 264.7 cells. After 24-hour incubation with both LPS (1 µg/mL) and EtOAc fractions of the seaweeds (12.5, 25, 50, or 100 µg/mL), a remarkable inhibition of TNF-α and IL-6 release was noted. As shown in Figures 5 and 6,D. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinasignificantly inhibited the production of TNF-α and IL-6 in a dose-dependent manner with IC50values of 37.35, 80.08, 103.28, 62.53, and 84.28 µg/mL for TNF-α and 16.11, 28.21, 84.27, 45.52, and 74.75 for IL-6, respectively.
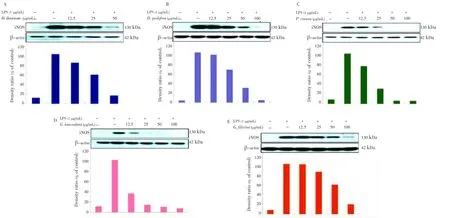
Figure 3. Effect of Jeju seaweed extracts on the activation of iNOS in LPS-stimulated RAW 264.7 cells.The RAW 264.7 cells (2.5×106cells/mL) were stimulated with LPS (1 µg/mL) in Jeju seaweed extracts (12.5, 25, 50, and 100 µg/mL) for 24 h. Whole-cell lysates (25 µg) were prepared, and the protein lysates were subjected to 10% SDS-PAGE; the expressions of iNOS and β-actin were determined by western blotting. Quantification of band intensities from three independent results was determined by densitometric analysis. The figure is representative of 3 similar experiments. The Jeju seaweed extract fractions were prepared as described in the materials and methods section.

Figure 4. Effect of Jeju seaweed extracts on the activation of COX-2 in LPS-stimulated RAW 264.7 cells.The RAW 264.7 cells (2.5×106cells/mL) were stimulated with LPS (1 µg/mL) in Jeju seaweed extracts (12.5, 25, 50, and 100 µg/mL) for 24 h. Whole-cell lysates (25 µg) were prepared, and the protein lysates were subjected to 10% SDSPAGE; the expressions of iNOS and β-actin were determined by western blotting. Quantification of band intensities from three independent results was determined by densitometric analysis. The figure is representative of 3 similar experiments. The Jeju seaweed extract fractions were prepared as described in the materials and methods section.
3.6. Phlorotannin contents of the Jeju seaweed extracts
Phlorotannins are aromatic secondary plant metabolites, which are widespread in brown seaweeds and associated with anti-inflammatory activity[25,26], hepatoprotective effects[27,28], anti-diabetic effects[29], radioprotective properties[30], and angiotensin-converting-enzyme inhibitory activity[31]. Therefore, the total phlorotannin content in all extracts was assessed, and the results were as follows: 92.0 µg/mg forD. divaricate, 151.8 µg/mg forD. prolifera, 57.2 µg/mg forP. cornea, 53.0 µg/mg forG. lanceolata, and 40.2 µg/mg forG. filicina.D. prolifera, brown algae, showed the highest phlorotannin content (>150 µg/mg).
4. Discussion
Marine algae are popular sea vegetables, which are consumed as health food by many people. Since ancient times, they have also been used in alternative medicine in such countries as Korea, China, and Japan[32]. Marine algae are a source of numerous natural products because they contain a wide variety of chemical structures and exhibit a broad range of biological and pharmacological activities (e.g., anti-inflammatory[10-11], hepatoprotective[27,28], antioxidant[33,34], anti-proliferative[35,36], antiviral[37,38], and anti-diabetic[29,39]). Recent studies have also shown that marine algae contain a wide range of highly bioactive secondary metabolites with therapeutic potential that might represent useful leads in the development of functional ingredients for neutraceuticals, cosmeceuticals, and pharmaceuticals[32,40,41]. However, the possibility of developing these functional ingredients is limited becausethe primary activity of algal ethanol extracts could be easily missed owing to the salts of the seaweeds on the surface. For this reason, the present study investigated the effect of Jeju seaweeds on inflammation by using their EtOAc fractions, which completely eliminates the salts of the seaweeds on the surface. Indeed, the activity of the ethanol extracts might be significantly different from that of the EtOAc fractions.
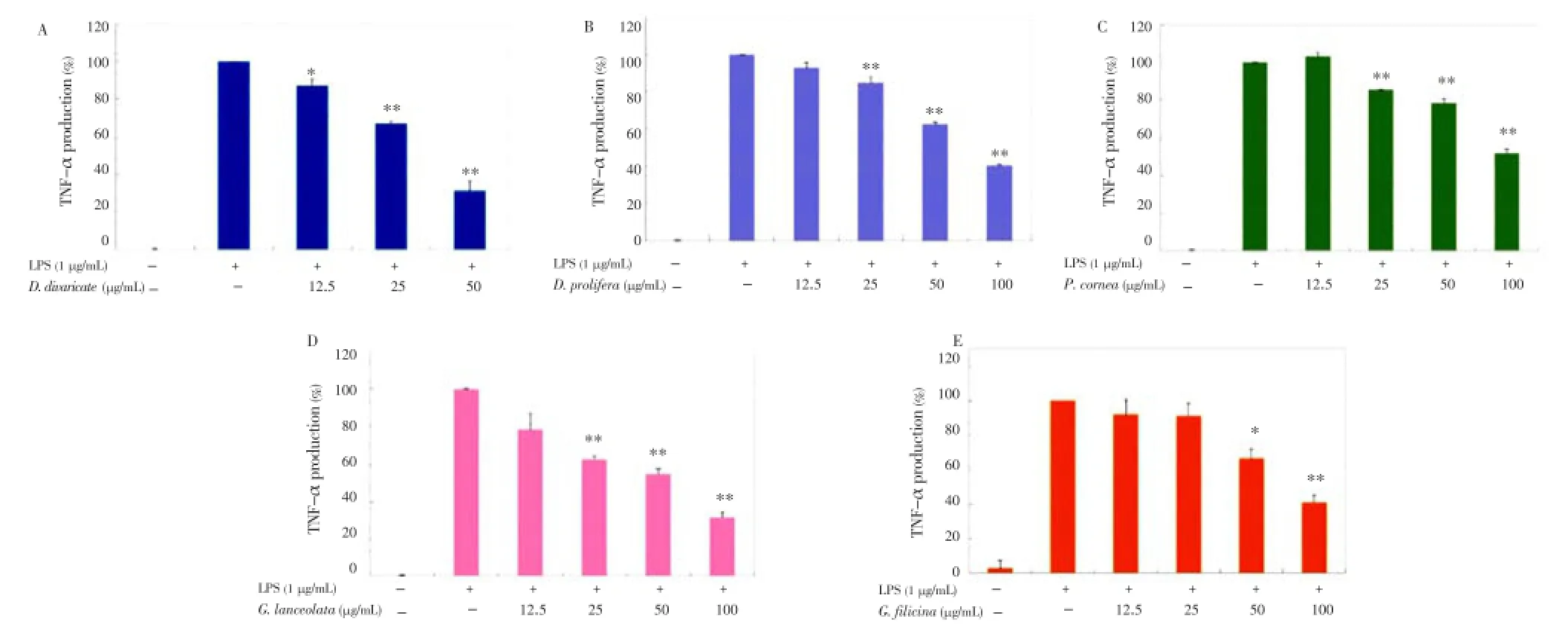
Figure 5. Effect of Jeju seaweed extracts on TNF-α production in LPS-stimulated RAW 264.7 cells.The cells were stimulated with 1 µg/mL of LPS alone or with a combination of LPS and various concentrations (12.5, 25, 50, or 100 µg/mL) of D. divaricate (A), D. prolifera (B), P. cornea (C), G. lanceolata (D), and G. filicina (E) for 24 h. TNF-α produced and released into the culture medium was assayed by the ELISA method. The data represent the mean±SD of triplicate experiments. *P<0.05; **P<0.01 vs. LPS alone.
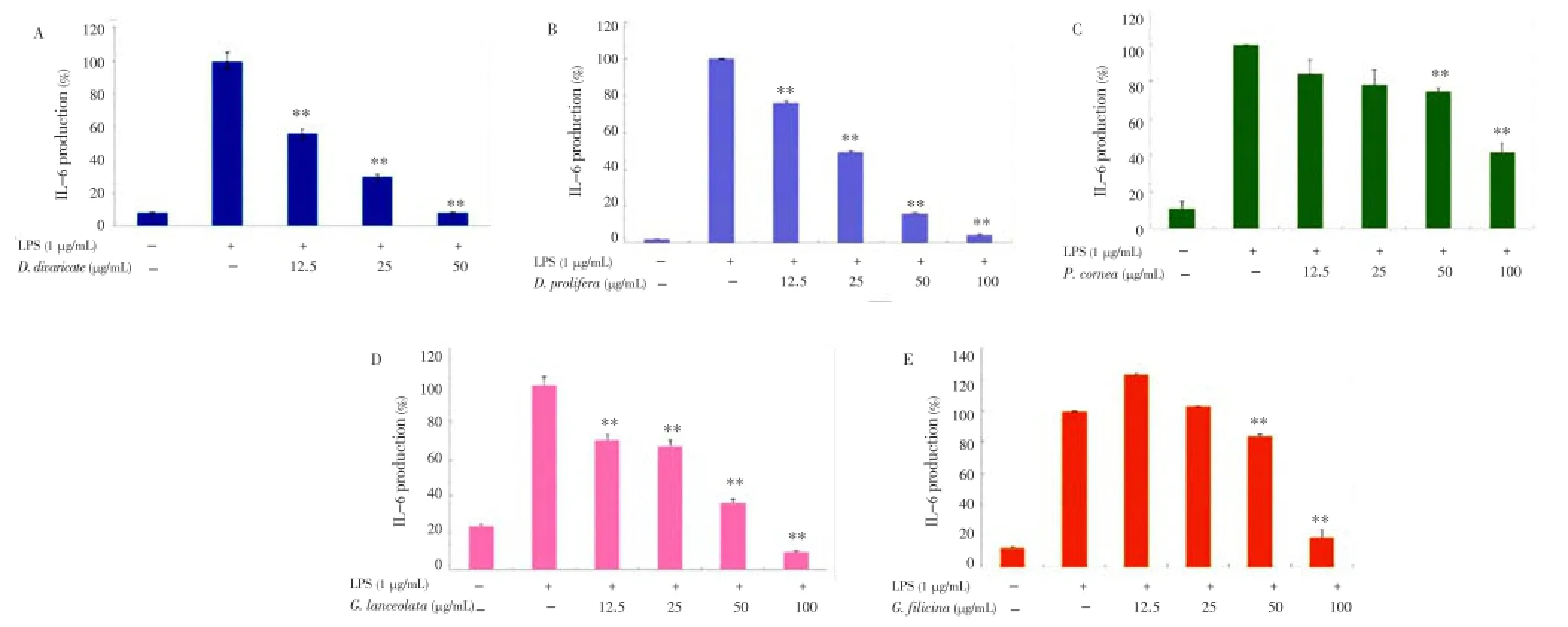
Figure 6. Effect of Jeju seaweed extracts on IL-6 production in LPS-stimulated RAW 264.7 cells.The cells were stimulated with 1 µg/mL of LPS alone or with a combination of LPS and various concentrations (12.5, 25, 50, or 100 µg/mL) of D. divaricate (A), D. prolifera (B), P. cornea (C), G. lanceolata (D), and G. filicina (E) for 24 h. IL-6 produced and released into the culture medium was assayed by the ELISA method. The data represent the mean±SD of triplicate experiments. *P<0.05; **P<0.01 vs. LPS alone.
The proinflammatory mediators, PGE2and NO, which are produced by activated macrophages, play an important role in the pathogenesis of various chronic inflammatory diseases such as hay fever, periodontitis, atherosclerosis, rheumatoid arthritis, and even cancer. Hence, the blocking effects of these proinflammatory mediators or iNOS and COX-2 expression in inflammatory cells provide an important therapeutic strategy for the treatment of chronic inflammation. Indeed, a number of natural products that inhibit the production of these inflammatory mediators have been previously considered as potential anti-inflammatory candidates[42-45].
Murine macrophage RAW 264.7 cells are an example of inflammatory cells that play a crucial role in specific and nonspecific immune responses during the inflammatory process. After macrophages are activated by stimuli, large amounts of the proinflammatory mediators (PGE2and NO) and cytokines (TNF-α and IL-6) are released. Interestingly, the production of these inflammatory molecules by macrophages can be easily induced in response to LPS stimulation[46-49]. Therefore, LPS-stimulated macrophages have been typically used for assessing the anti-inflammatory potential of natural products.
Because excessive NO production has been shown to cause acute and chronic inflammation, pharmacologicalinterference of NO production has been speculated to offer promising strategies for alleviating inflammatory disorders. To validate the use of Jeju seaweed extracts (D. divaricate,D. prolifera,P. cornea,G. lanceolata, orG. filicina) as antiinflammatory ingredients, we first investigated the effect of the extracts on NO synthesis in LPS-stimulated macrophages RAW 264.7 cells, and the results indicated that all extracts were effective inhibitors of LPS-induced NO production in these cells. The study showed that those inhibitory effects were accompanied by a decrease in the levels of iNOS expression as evidenced by dose-dependent reductions, mediated byD. divaricate,D. prolifera,P. cornea,G. lanceolata, orG. filicina. These results suggest that Jeju seaweeds have potent inhibitory activity on proinflammatory mediator production.
PGE2is also an important mediator of the inflammatory response produced by COX-2 expression, which is induced by cytokines, LPS, growth factors, mitogens, cytokines, and tumor promoters during inflammation, resulting in the production of large amounts of PGE2at the site of inflammation[50-52]. In this study, we showed that in LPS-stimulated macrophages,D. divaricate,D. prolifera,P. cornea, orG. filicinainhibited PGE2production in a dosedependent manner. COX-2 was also downregulated byD. divaricate,D. prolifera,P. cornea, andG. filicina, suggesting that this was the mechanism underlying the observed suppression of PGE2production.
It has been reported that proinflammatory cytokines such as TNF-α and IL-6 are involved in a variety of immune reactions and interactions with a variety of target cells bothin vitroandin vivo[53-55]. They are considered to be important initiators of the inflammatory response and mediators of the development of various inflammatory diseases. Therefore, we chose these parameters to investigate additional anti-inflammatory effects of the Jeju seaweed extracts. Their inhibitory effects on LPS-stimulated TNF-α and IL-6 were assessed using the EIA kit, and the data showed that treatment with the extracts significantly inhibited the secretion of LPS-induced TNF-α and IL-6 in a concentration-dependent manner. These results suggest thatD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinaexert their anti-inflammatory effects via inhibition of TNF-α and IL-6 production, which has important implications for the development of therapeutic strategies in chronic inflammation.
In summary, as part of our research on the antiinflammatory potential of the Jeju seaweeds, we previously reported their numerous inhibitory effects on the release of various inflammatory mediators induced by LPS. The results of the present study show that the anti-inflammatory effects ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicinaare achieved via the modulation of macrophage-mediated inflammatory markers such as NO, PGE2, IL-1β, and IL-6, as well as protein levels of iNOS and COX-2. Thus, considering their potent effects, these extracts--whether used separately or in combination--may become extremely useful in the prevention and treatment of various inflammatory diseases. Our study provides a better understanding of the health-promoting effects of a phytochemical that is widely consumed worldwide. In future studies, we will investigate the molecular mechanisms underlying the anti-inflammatory effect ofD. divaricate,D. prolifera,P. cornea,G. lanceolata, andG. filicina, such as NF-κB and/or MAPKs signaling pathways, and the ability of the seaweed extracts to stimulate the immune system in anin vivomodel.
Conflict of interest statement
We declare that we have no conflict of interest.
Acknowledgements
This research was financially supported by the fostering program of regionally specialized industries through the Korea Institute for Advancement of Technology (KIAT) and Jeju Institute for Regional Program Evaluation funded by the Ministry of Trade, Industry and Energy (2013-R0002288).
Comments
Background
Many attempts have been made to derive new antiinflammatory agents from natural sources of phytochemicals that have been considered safe, less toxic, and readily available, though their modes of action mostly remain unclear. Thus, elucidating the molecular mechanisms underlying the anti-inflammatory actions of naturally occurring phytochemicals might be a good strategy for identifying new therapeutic agents.
Research frontiers
The present study depicts the anti-inflammatory effects ofD. divaricata, D. prolifera, P. cornea, G. lanceolata, and G. filicinaon LPS-induced inflammation in murine RAW 264.7 macrophage cells, and the underlying mechanisms.
Related reports
A series of studies have been done for centuries to discover the anti-inflammatory potential of certain seaweeds. The extracts ofD. divaricata,D. prolifera,P. cornea,G. lanceolata, andG. filicinahave not been tested for any anti-inflammatory assay and this study is considered novel in their specific field.
Innovations and breakthroughs
The study have showed the anti-inflammatory potentialsof Jeju seaweed extract (D. divaricata, D. prolifera, P.cornea, G. lanceolata, and G. filicina). The study is highly significant in the field of pharmacology in which the developing novel drugs pose anti-inflammatory agents in near future.
Applications
This study provides basis for the traditional use of Jeju seaweeds in the treatment of inflammatory pathologies. Other pharmacological effects such as anti-aging and antimelanogenesis could be explored.
Peer review
This is a good study in which the authors evaluated the effect of ethylacetate extracts of Jeju seaweed that are effective against LPS-induced inflammation in murine macrophage cells. The results are interesting and suggest that Jeju seaweeds may be used as alternative therapy for anti-inflammatory diseases.
[1] Dellai A, Laajili S, Le Morvan V, Robert J, Bouraoui A. Antiproliferative activity and phenolics of the Mediterranean seaweed Laurencia obusta. Ind Crops Prod 2013; 47: 252-255.
[2] Balboa EM, Conde E, Moure A, Falqué E, Domínguez H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem 2013; 138(2-3): 1764-1785.
[3] Wang W, Wang SX, Guan HS. The antiviral activities and mechanisms of marine polysaccharides: an overview. Mar Drugs 2012; 10(12): 2795-2816.
[4] Kharkwal H, Joshi DD, Panthari P, Pant MK, Kharkwal AC. Algae as future drugs. Asian J Pharm Clin Res 2012; 5: 1-4.
[5] Wijesinghe WA, Jeon YJ. Exploiting biological activities of brown seaweed Ecklonia cava for potential industrial applications: a review. Int J Food Sci Nutr 2012; 63(2): 225-235.
[6] Yang EJ, Moon JY, Kim MJ, Kim DS, Lee WJ, Lee NH, et al. Antiinflammatory effect of Petalonia binghamiae in LPS-induced macrophages is mediated by suppression of iNOS and COX-2. Int J Agri Biol 2010; 12(5): 754-758.
[7] Yoon WJ, Ham YM, Lee WJ, Lee NH, Hyun CG. Brown alga Sargassum muticum inhibits proinflammatory cytokines, iNOS, and COX-2 expression in macrophage RAW 264.7 cells. Turk J Biol 2010; 34(1): 25-34.
[8] Hwang PA, Chien SY, Chan YL, Lu MK, Wu CH, Kong ZL, et al. Inhibition of Lipopolysaccharide (LPS)-induced inflammatory responses by Sargassum hemiphyllum sulfated polysaccharide extract in RAW 264.7 macrophage cells. J Agric Food Chem 2011; 59(5): 2062-2068.
[9] Kazłowska K, Hsu T, Hou CC, Yang WC, Tsai GJ. Antiinflammatory properties of phenolic compounds and crude extract from Porphyra dentata. J Ethnopharmacol 2010; 128(1): 123-130.
[10] Yang EJ, Ham YM, Lee WJ, Lee NH, Hyun CG. Anti-inflammatory effects of apo-9’-fucoxanthinone from the brown alga, Sargassum muticum. Daru 2013; 21(1): 62.
[11] Ham YM, Kim KN, Lee WJ, Lee NH, Hyun CG. Chemical constituents from Sargassum micracanthum and antioxidant activity. Int J Pharmacol 2010; 6(2): 147-151.
[12] Liu L, Heinrich M, Myers S, Dworjanyn SA. Towards a better understanding of medicinal uses of the brown seaweed Sargassum in Traditional Chinese Medicine: a phytochemical and pharmacological review. J Ethnopharmacol 2012; 142(3): 591-619.
[13] Yu T, Yi YS, Yang Y, Oh J, Jeong D, Cho JY. The pivotal role of TBK1 in inflammatory responses mediated by macrophages. Mediators Inflamm 2012; doi: 10.1155/2012/979105.
[14] Ayissi Owona B, Njayou NF, Laufer S, Moundipa PF, Schluesener HJ. A fraction of stem bark extract of Entada africana suppresses lipopolysaccharide-induced inflammation in RAW 264.7 cells. J Ethnopharmacol 2013; 149(1): 162-168.
[15] Zhu J, Luo C, Wang P, He Q, Zhou J, Peng H. Saikosaponin A mediates the inflammatory response by inhibiting the MAPK and NF-κB pathways in LPS-stimulated RAW 264.7 cells. Exp Ther Med 2013; 5(5): 1345-1350.
[16] Philippou A, Maridaki M, Theos A, Koutsilieris M. Cytokines in muscle damage. Adv Clin Chem 2012; 58: 49-87.
[17] Lee IT, Yang CM. Role of NADPH oxidase/ROS in proinflammatory mediators-induced airway and pulmonary diseases. Biochem Pharmacol 2012; 84(5): 581-590.
[18] Romeo GR, Lee J, Shoelson SE. Metabolic syndrome, insulin resistance, and roles of inflammation--mechanisms and therapeutic targets. Arterioscler Thromb Vasc Biol 2012; 32(8): 1771-1776.
[19] Wang Y, Yu C, Pan Y, Li J, Zhang Y, Ye F, et al. A novel compound C12 inhibits inflammatory cytokine production and protects from inflammatory injury in vivo. PLoS One 2011; 6(9): e24377.
[20] Borges MC, Vinolo MA, Crisma AR, Fock RA, Borelli P, Tirapegui J, et al. High-fat diet blunts activation of the nuclear factor-κB signaling pathway in lipopolysaccharide-stimulated peritoneal macrophages of Wistar rats. Nutrition 2013; 29(2): 443-449.
[21] Aoki T, Narumiya S. Prostaglandins and chronic inflammation. Trends Pharmacol Sci 2012; 33(6): 304-311.
[22] Murakami A, Ohigashi H. Targeting NOX, INOS and COX-2 in inflammatory cells: chemoprevention using food phytochemicals. Int J Cancer 2007; 121(11): 2357-2363.
[23] Huang D, Ou B, Prior RL. The chemistry behind antioxidant capacity assays. J Agric Food Chem 2005; 53(6): 1841-1856.
[24] Yang EJ, Moon JY, Kim MJ, Kim DS, Kim CS, Lee WJ, et al. Inhibitory effect of Jeju endemic seaweeds on the production of pro-inflammatory mediators in mouse macrophage cell line RAW 264.7. J Zhejiang Univ Sci B 2010; 11(5): 315-322.
[25] Jung HA, Jin SE, Ahn BR, Lee CM, Choi JS. Anti-inflammatory activity of edible brown alga Eisenia bicyclis and its constituents fucosterol and phlorotannins in LPS-stimulated RAW264.7 macrophages. Food Chem Toxicol 2013; 59: 199-206.
[26] Dutot M, Fagon R, Hemon M, Rat P. Antioxidant, antiinflammatory, and anti-senescence activities of a phlorotanninrich natural extract from brown seaweed Ascophyllum nodosum. Appl Biochem Biotechnol 2012; 167(8): 2234-2240.
[27] Kang MC, Ahn G, Yang X, Kim KN, Kang SM, Lee SH, et al. Hepatoprotective effects of dieckol-rich phlorotannins fromEcklonia cava, a brown seaweed, against ethanol induced liver damage in BALB/c mice. Food Chem Toxicol 2012; 50(6): 1986-1991.
[28] Kim SM, Kang K, Jeon JS, Jho EH, Kim CY, Nho CW, et al. Isolation of phlorotannins from Eisenia bicyclis and their hepatoprotective effect against oxidative stress induced by tertbutyl hyperoxide. Appl Biochem Biotechnol 2011; 165(5-6): 1296-1307.
[29] Kang MC, Wijesinghe WA, Lee SH, Kang SM, Ko SC, Yang X, et al. Dieckol isolated from brown seaweed Ecklonia cava attenuates type ІІ diabetes in db/db mouse model. Food Chem Toxicol 2013; 53: 294-298.
[30] Moon C, Kim SH, Kim JC, Hyun JW, Lee NH, Park JW, et al. Protective effect of phlorotannin components phloroglucinol and eckol on radiation-induced intestinal injury in mice. Phytother Res 2008; 22(2): 238-242.
[31] Wijesekara I, Kim SK. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: prospects in the pharmaceutical industry. Mar Drugs 2010; 8(4): 1080-1093.
[32] Thomas NV, Kim SK. Beneficial effects of marine algal compounds in cosmeceuticals. Mar Drugs 2013; 11(1): 146-164.
[33] Fung A, Hamid N, Lu J. Fucoxanthin content and antioxidant properties of Undaria pinnatifida. Food Chem 2013; 136(2): 1055-1062.
[34] Zhang CY, Wu WH, Wang J, Lan MB. Antioxidant properties of polysaccharide from the brown seaweed Sargassum graminifolium (Turn.), and its effects on calcium oxalate crystallization. Mar Drugs 2012; 10(1): 119-130.
[35] Kang KH, Kim SK. Beneficial effect of peptides from microalgae on anticancer. Curr Protein Pept Sci 2013; 14(3): 212-217.
[36] Senthilkumar K, Manivasagan P, Venkatesan J, Kim SK. Brown seaweed fucoidan: biological activity and apoptosis, growth signaling mechanism in cancer. Int J Biol Macromol 2013; 60: 366-374.
[37] Koishi AC, Zanello PR, Bianco ÉM, Bordignon J, Nunes Duarte dos Santos C. Screening of Dengue virus antiviral activity of marine seaweeds by an in situ enzyme-linked immunosorbent assay. PLoS One 2012; 7(12): e51089.
[38] Chen KJ, Tseng CK, Chang FR, Yang JI, Yeh CC, Chen WC, et al. Aqueous extract of the edible Gracilaria tenuistipitata inhibits hepatitis C viral replication via cyclooxygenase-2 suppression and reduces virus-induced inflammation. PLoS One 2013; 8(2): e57704.
[39] Lordan S, Smyth TJ, Soler-Vila A, Stanton C, Ross RP. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem 2013; 141(3): 2170-2176.
[40] Blunt JW, Copp BR, Keyzers RA, Munro MH, Prinsep MR. Marine natural products. Nat Prod Rep 2013; 30(2): 237-323.
[41] Blunt JW, Copp BR, Keyzers RA, Munro MH, Prinsep MR. Marine natural products. Nat Prod Rep 2012; 29(2): 144-222.
[42] Shin JH, Ryu JH, Kang MJ, Hwang CR, Han J, Kang D. Shortterm heating reduces the anti-inflammatory effects of fresh raw garlic extracts on the LPS-induced production of NO and proinflammatory cytokines by downregulating allicin activity in RAW 264.7 macrophages. Food Chem Toxicol 2013; 58: 545-551.
[43] Hao H, Gufu H, Lei F, Dang L, Zhongliang Y. Baicalin suppresses expression of TLR2/4 and NF-κB in chlamydia trachomatisinfected mice. Immunopharmacol Immunotoxicol 2012; 34(1): 89-94.
[44] Park JY, Pillinger MH, Abramson SB. Prostaglandin E2 synthesis and secretion: the role of PGE2synthases. Clin Immunol 2006; 119(3): 229-240.
[45] Blobaum AL, Marnett LJ. Structural and functional basis of cyclooxygenase inhibition. J Med Chem 2007; 50(7): 1425-1441.
[46] Hald A, Rønø B, Lund LR, Egerod KL. LPS counter regulates RNA expression of extracellular proteases and their inhibitors in murine macrophages. Mediators Inflamm 2012; 2012: 1-9.
[47] Nguyen MT, Chen A, Lu WJ, Fan W, Li PP, Oh DY, et al. Regulation of chemokine and chemokine receptor expression by PPARγ in adipocytes and macrophages. PLoS One 2012; 7(4): e34976.
[48] Cui YQ, Jia YJ, Zhang T, Zhang QB, Wang XM. Fucoidan protects against lipopolysaccharide-induced rat neuronal damage and inhibits the production of proinflammatory mediators in primary microglia. CNS Neurosci Ther 2012; 18(10): 827-833.
[49] Lee JW, Bae CJ, Choi YJ, Kim SI, Kim NH, Lee HJ, et al. 3,4,5-Trihydroxycinnamic acid inhibits LPS-induced iNOS expression by suppressing NF-κB activation in BV2 microglial cells. Korean J Physiol Pharmacol 2012; 16(2): 107-112.
[50] Park HH, Kim MJ, Li Y, Park YN, Lee J, Lee YJ, et al. Britanin suppresses LPS-induced nitric oxide, PGE2and cytokine production via NF-κB and MAPK inactivation in RAW 264.7 cells. Int Immunopharmacol 2013; 15(2): 296-302.
[51] Yang G, Lee K, Lee M, Ham I, Choi HY. Inhibition of lipopolysaccharide-induced nitric oxide and prostaglandin E2production by chloroform fraction of Cudrania tricuspidata in RAW 264.7 macrophages. BMC Complement Altern Med 2012; 12: 250.
[52] Han S, Lee JH, Kim C, Nam D, Chung WS, Lee SG, et al. Capillarisin inhibits iNOS, COX-2 expression, and proinflammatory cytokines in LPS-induced RAW 264.7 macrophages via the suppression of ERK, JNK, and NF-κB activation. Immunopharmacol Immunotoxicol 2013; 35(1): 34-42.
[53] Wu Q, Sun G, Yuan X, Soromou LW, Chen N, Xiong Y, et al. Tubeimoside-1 attenuates LPS-induced inflammation in RAW 264.7 macrophages and mouse models. Immunopharmacol Immunotoxicol 2013; 35(4): 514-523.
[54] Kwon DJ, Ju SM, Youn GS, Choi SY, Park J. Suppression of iNOS and COX-2 expression by flavokawain A via blockade of NF-κB and AP-1 activation in RAW 264.7 macrophages. Food Chem Toxicol 2013; 58: 479-486.
[55] Oh YC, Cho WK, Jeong YH, Im GY, Lee KJ, Yang HJ, et al. Antiinflammatory effect of Sosihotang via inhibition of nuclear factorκB and mitogen-activated protein kinases signaling pathways in lipopolysaccharide-stimulated RAW 264.7 macrophage cells. Food Chem Toxicol 2013; 53: 343-351.
10.12980/APJTB.4.2014C1099
*Corresponding author: Chang-Gu Hyun, Cosmetic Science Center and LINC Agency, Jeju National University, Jeju 690-756, Korea.
Tel: +82 64 754 4419
Fax: +82 64 751 3127
E-mail: cghyun@jejunu.ac.kr
Foundation Project: Supported by the fostering program of regionally specialized industries through the Korea Institute for Advancement of Technology (KIAT) and Jeju Institute for Regional Program Evaluation funded by the Ministry of Trade, Industry and Energy (2013-R0002288).
Article history:
Received 3 May 2014
Received in revised form 10 May, 2nd revised form 16 May, 3rd revised form 22 May, 2014
Accepted 25 Jun 2014
Available online 28 Jul 2014
Methods:Ethyl acetate fractions were prepared from five different types of Jeju seaweeds, Dictyopteris divaricata (D. divaricata), Dictyopteris prolifera (D. prolifera), Prionitis cornea (P. cornea), Grateloupia lanceolata (G. lanceolata), and Grateloupia filicina (G. filicina). They were screened for inhibitory effects on proinflammatory mediators and cytokines such as nitric oxide (NO), prostaglandin E2, tumor necrosis factor-α (TNF-α), and interleukin-6 (IL-6).
Results:Our results revealed that D. divaricata, D. prolifera, P. cornea, G. lanceolata, and G. filicina potently inhibited LPS-stimulated NO production (IC50values were 18.0, 38.36, 38.43, 32.81 and 37.14 µg/mL, respectively). Consistent with these findings, D. divaricata, D. prolifera, P. cornea, and G. filicina also reduced the LPS-induced and prostaglandin E2production in a concentration-dependent manner. Expectedly, they suppressed the expression of inducible NO synthase and cyclooxygenase-2 at the protein level in a dose-dependent manner in the RAW 264.7 cells, as determined by western blotting. In addition, the levels of TNF-α and IL-6, released into the medium, were also reduced by D. divaricata, D. prolifera, P. cornea, G. lanceolata, and G. filicina in a dose-dependent manner (IC50values for TNF-α were 16.11, 28.21, 84.27, 45.52 and 74.75 µg/mL, respectively; IC50values for IL-6 were 37.35, 80.08, 103.28, 62.53 and 84.28 µg/mL, respectively). The total phlorotannin content was measured by the Folin-Ciocalteu method and expressed as phloroglucinol equivalents. The content was 92.0 µg/mg for D. divaricata, 151.8 µg/ mg for D. prolifera, 57.2 µg/mg for P. cornea, 53.0 µg/mg for G. lanceolata, and 40.2 µg/mg for G. filicina.
Conclusions:Thus, these findings suggest that Jeju seaweed extracts have potential therapeutic applications for inflammatory responses.
杂志排行

Asian Pacific Journal of Tropical Biomedicine的其它文章
- Antimicrobial activity against periodontopathogenic bacteria, antioxidant and cytotoxic effects of various extracts from endemic Thermopsis turcica
- Proteomics analysis of antimalarial targets of Garcinia mangostana Linn.
- The presence of eucalyptol in Artemisia australis validates its use in traditional Hawaiian medicine
- Antioxidant potential of Rumex vesicarius L.: in vitro approach
- Cytotoxicity screening of Melastoma malabathricum extracts on human breast cancer cell lines in vitro
- In vitro cytotoxicity of Indonesian stingless bee products against human cancer cell lines