重组大肠杆菌表达水稻白叶枯病菌FtsZ蛋白
2014-03-21陈洋黄运红李素珍龙中儿
陈洋 黄运红 李素珍 龙中儿
(江西师范大学生命科学学院,南昌 330022)
重组大肠杆菌表达水稻白叶枯病菌FtsZ蛋白
陈洋 黄运红 李素珍 龙中儿
(江西师范大学生命科学学院,南昌 330022)
旨在通过现代分子生物学技术制备水稻白叶枯病菌FtsZ蛋白。以水稻白叶枯病菌总DNA为模板,采用巢式PCR方法扩增获得水稻白叶枯病菌ftsZ基因,构建ftsZ基因的表达载体pET-22b-ftsZ,转化表达宿主E. coli BL21后,经PCR、Nde I/ Xho I双酶切及测序鉴定、阳性克隆子经IPTG诱导表达,融合蛋白经镍柱纯化后,通过SDS-PAGE和Western blotting分析鉴定。结果显示,水稻白叶枯病菌ftsZ基因的重组表达载体构建成功,且阳性克隆子在IPTG的诱导下表达了FtsZ-6×His融合蛋白,并通过镍柱纯化获得了电泳纯的FtsZ-6×His融合蛋白。
FtsZ蛋白 水稻白叶枯病菌 大肠杆菌 基因重组
FtsZ蛋白是真核细胞微管蛋白的原核同系物,在细菌体内高度保守,具有GTPase活性,并能受GTP的诱导发生聚合,是原核生物细胞分裂的关键蛋白之一[1]。由于在细胞分裂中的重要作用及其高度的保守性,FtsZ是抗菌药物开发的理想靶标之一[2]。水稻白叶枯病是水稻白叶枯病菌(Xanthomonas oryzae)引起的水稻生产上的严重病害之一,导致水稻叶片枯萎,光合作用受到妨碍,稻穗充实率降低,不实粒增加,同时可造成米质疏松脆裂,食味降低[3,4]。目前,常用手段是使用噻枯唑作为防治水稻白叶枯病的主要化学药剂,但水稻白叶枯病菌对噻枯唑的敏感性逐渐下降,抗药突变体已出现[5,6]。因此,研发经济、高效、无毒的抗水稻白叶枯病菌的农用抗生素具有重要的意义。
本研究构建pET-22b-ftsZ重组表达载体,进行水稻白叶枯病菌FtsZ的体外表达,旨在为进一步构建靶向抑制FtsZ的抗生素筛选模型奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌种与质粒 水稻白叶枯病菌(X. oryzae),购自武汉大学中国典型培养物保藏中心(菌株保藏编号:CCTCC AB 91123),Escherichia coli DH5α感受态细胞和pMD19-T Vector克隆载体购自大连宝生物公司,E. coli BL21感受态细胞由南昌大学惠赠,
pET-22b(+)载体由本实验室于-20℃冰箱保存。
1.1.2 培养基 NB液体培养基,LB固体培养基,LB液体培养基,SOC培养基。
1.1.3 试剂 DNA分子量标准、蛋白质分子量标准、限制性内切酶Nde I、Xho I为大连宝生物公司产品,GoldView、咪唑为Solarbio公司产品,胰蛋白胨和酵母提取物为Oxoid公司产品,Ezup柱式细菌基因组提取试剂盒、SanPrep柱式DNA凝胶回收试剂盒和SanPrep柱式质粒DNA小量提取试剂盒、SDS、Tris、IPTG、氨苄青霉素为生工生物工程(上海)有限公司产品,Protein IsoTMNi-NTA Resin和预染蛋白Marker为北京全式金产品,抗His标签鼠单克隆抗体(一抗)和HRP标记的羊抗鼠多克隆抗体(二抗)为Sigma公司产品,其他试剂均为国产分析纯。
1.2 方法
1.2.1 水稻白叶枯病菌总DNA的提取 采用上海生工生物工程Ezup柱式细菌基因组提取试剂盒提取,具体操作按说明书进行。
1.2.2 ftsZ基因的扩增 根据NCBI上X. oryzae pv. Oryzae KACC10331全基因组序列(ID:58155)及其ftsZ基因(ID:3264933),分别设计扩增ftsZ的引物(forward1:5'-GGCGAGCAGGTAGGCGGTAAAATT-3',backward1:5'-ACAATGCAGACAGACGCTGAATCGA-3')和特异性引物(forward2:5'-GGAATTCCATATGGCACATTTCGAACTGATTG-3',backward2:5'-CCGCTCGAGGTCGGCCTGGCGGCGCAGG-3')。50 μL PCR反应体系含TaKaRa LA Taq(5 U/μL)0.5 μL,2× GC PCR Buffer I 25 μL,dNTP Mixture(各2.5 mmol/L)8 μL,基因组DNA 1 μL,forward1(25 μmol/L)1 μL,backward1(25 μmol/L)1 μL,灭菌超纯水13.5 μL。反应条件为:94℃预变性1 min;94℃变性1 min,57℃退火30 s,72℃延伸90 s,30个循环;72℃延伸 5 min。第二次PCR除更换引物,以第一次PCR产物为模板外,其余条件不变。扩增后,琼脂糖凝胶电泳检测扩增产物。
1.2.3 pET-22b-ftsZ重组质粒的构建 用上海生工SanPrep柱式DNA胶回收试剂盒回收目的条带,按pMD19-T Vector说明书配置连接体系,并转化入感受态细胞E. coli DH5α后,涂布于含50 μg/mL氨苄西林钠的LB固体培养基,37℃培养12 h后提取质粒,通过质粒PCR及Nde I 和Xho I 限制性内切酶双酶切鉴定出阳性克隆子后送上海生工测序[7]。
用 Nde I和Xho I限制性内切酶分别处理pET-22b(+)载体和目的基因片段,将处理后的质粒与目的基因连接,连接条件参考pMD19-T Vector说明书。重组质粒以同样的方式转入感受态E. coli BL21,以载体的氨苄抗性标记初筛转化子,再提取质粒,以Nde I和Xho I限制性内切酶双酶切及质粒PCR鉴定,将阳性克隆子送至上海生工测序[8]。
1.2.4 目的基因的诱导表达 取1 mL培养过夜的E.coli BL21阳性转化子菌悬液接种于装有50 mL LB液体培养基的锥形瓶中,200 r/min、37℃培养3 h[9],再加IPTG母液使其终浓度为0.6 mmol/L,继续培养3 h。取诱导菌悬液1 mL于12 000 r/min 离心30 s,弃上清,加100 μL 1% SDS重悬菌体,沸水浴5 min后,SDS-PAGE电泳分析表达结果[10]。
1.2.5 目的蛋白的纯化 取经IPTG诱导表达的菌悬液以8 000 r/min 离心20 min,弃上清,加10倍菌体质量的裂解液,超声破碎(3 s工作,5 s间歇,99个循环,400 W功率)后,再12 000 r/min离心30 min,取上清上镍柱纯化,最后以1倍柱体积的500 mmol/L 咪唑平衡液洗脱目的蛋白。
1.2.6 Western blotting分析 按文献[10]的方法进行。具体操作如下:将咪唑洗脱下的蛋白溶液进行SDS-PAGE电泳;剪一块与胶大小相当的PVDF膜,以甲醇浸泡60 s活化,并放入转膜液中,取3张经转膜液浸泡过的大滤纸放在转膜仪上,再依次放上PVDF膜、SDS-PAGE胶和3张经过转膜液浸泡过的略小于胶的滤纸(注意避免产生气泡),盖上转膜仪,连接电泳仪后,67 mA恒流60 min,将目的蛋白从聚丙烯酰胺凝胶上转至PVDF膜上,而后将PVDF膜放入装有5%脱脂牛奶的培养皿中封闭60 min,10 mmol/L的PBS洗涤4次。将PVDF膜放入含10 mL(1∶5 000,V/V)His一抗的培养皿中,室温条件下中速水平摇床上孵育1 h,充分反应后,用0.01 mol/L PBS 洗涤4次,以除去未结合His一抗,后将与一抗反应的PVDF膜放入含10 mL(1∶5 000,V/V)羊抗鼠二抗的培养皿中,并在室温条件下中速的水平摇床上反应30 min,再用0.01 mol/LPBS洗涤4次,
以除去未结合羊抗鼠二抗体;在避光条件下,将与二抗反应后的PVDF膜放入DAB显色液中,反应10 min,0.01 mol/L PBS洗涤3次,晾干,照相保存。
2 结果
2.1 总DNA的提取
提取的水稻白叶枯病菌总DNA经1%的琼脂糖凝胶电泳,结果(图1)显示,DNA条带明显,通过核酸分析检测仪测得DNA浓度为81.8 μg/μL,丰度适宜,可用作PCR的模板。
图1 水稻白叶枯病菌总DNA
2.2 目的基因的获得
分别采用巢式PCR及普通PCR技术扩增目的基因,产物经1%琼脂糖凝胶电泳检测,结果(图2)显示,巢式PCR产物中的目的基因丰度和纯度更高,符合预期目的。
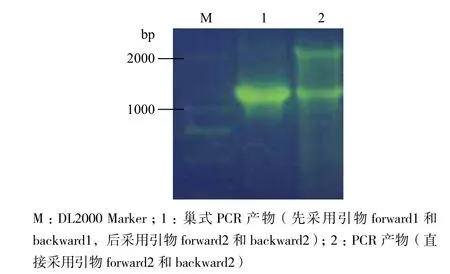
图2 ftsZ的PCR产物
2.3 pMD19-ftsZ重组质粒鉴定
2.3.1 质粒PCR鉴定 质粒PCR结果(图3)显示,以基因组DNA及以pMD19-ftsZ重组质粒为模板都扩增出含有目的基因大小的条带,说明pMD19-ftsZ重组质粒构建成功。
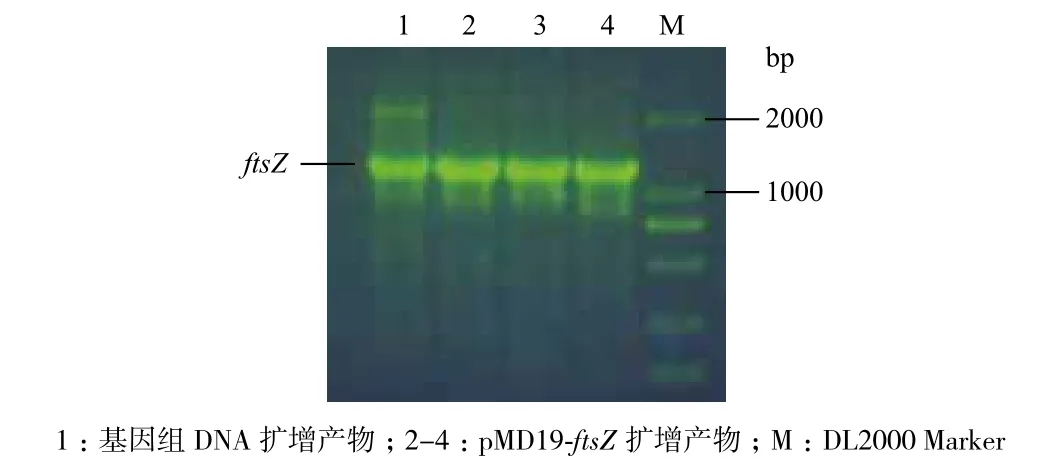
图3 pMD19-T-ftsZ重组质粒的PCR鉴定图
2.3.2 双酶切鉴定 用Nde I和Xho I对pMD19-ftsZ重组质粒进行双酶切后,结果(图4)显示,目的基因与pMD19-T Vector分离,大小符合预期。
2.4 pET-22b-ftsZ重组质粒鉴定
2.4.1 质粒PCR鉴定 pET-22b-ftsZ重组质粒的PCR结果(图5)与重组载体pMD19-ftsZ的质粒PCR结果一致,说明ftsZ基因已转入pET载体中。

图5 pET-22b-ftsZ融合质粒PCR鉴定
2.4.2 双酶切鉴定 用Nde I 和Xho I 对pET-22bftsZ重组质粒进行双酶切后,结果(图6)显示,目的基因与pET-22b(+)质粒分离,大小符合预期。
2.5 测序鉴定
将测序结果与NCBI数据库中X. oryzae pv. Ory-
zae KACC10331的ftsZ基因(ID:3264933)序列比对,两者匹配度为100%。

图6 pET-22b-ftsZ重组质粒的双酶切鉴定
2.6 FtsZ蛋白的表达与纯化
根据表达载体pET-22b(+)的特点及重组质粒pET-22b-ftsZ 的构建过程可以预测,E. coli BL21表达的FtsZ蛋白为一个C-末端带有组氨酸标签的融合蛋白(FtsZ-6×His融合蛋白),可以用镍柱纯化。FtsZ-6×His融合蛋白的表达与纯化结果(图7)显示,在细胞破碎上清液的电泳条带中,目的蛋白条带明显,证明目的蛋白以可溶的形式存在,其分子量略大于40 kD,符合预期估算的44 kD。纯化的FtsZ-6×His 融合蛋白以单条带的形式存在(图7泳道1),表明经镍柱纯化的FtsZ蛋白达到电泳纯。
同时,试验采用Western blotting分析了宿主细胞破碎后的上清液和经镍柱纯化后的目的蛋白溶液,电泳条带都在预期分子量大小的位点上显色(图8),结果证实了FtsZ蛋白表达成功,且表达的FtsZ蛋白为带有组氨酸标签的融合蛋白。
3 讨论
目前,有关水稻白叶枯病的研究,主要集中在水稻白叶枯病菌的致病性相关基因[12,13]、水稻白叶枯病菌的遗传多样性[14,15]、水稻白叶枯病菌抗性水稻的选育[16,17]以及抗水稻白叶枯病菌的药剂[18,19]等几个方面。噻枯唑是防治水稻白叶枯病的主要化学药剂,但水稻白叶枯病菌对噻枯唑的敏感性逐渐下降,抗药突变体已出现[3,4],尚未找到能够替代噻枯唑的有效化学药剂。研发抗水稻白叶枯病菌的新型抗生素对水稻白叶枯病菌的防治具有重要意义。
新抗生素的研发策略主要有三,一是通过对已知抗生素的结构改造、甚至全合成的手段获得已知抗生素的结构类似物[20,21],甚至全新结构的抗生素,以克服细菌的耐药现象;二是不断拓展抗生素来源,通过筛选新的微生物物种获得新的抗生素[22,23];三是创新抗生素筛选模型,将传统的抗生素离体筛选模型,如平板拮抗法、孢子萌发测定、杯碟法、纸片法等,发展到基于抗生素作用机理及特定靶标,尤其是全新靶标的高通量筛选模型[24,25]等。
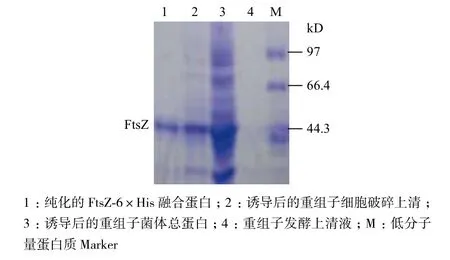
图7 FtsZ蛋白的纯化

图8 纯化FtsZ 的Western blotting分析
FtsZ蛋白是一种出现在原核细胞分裂位点上的第一个蛋白,起到细胞骨架作用,形成的FtsZ蛋白聚合体吸引与其结合的蛋白装配成原丝,螺旋成为Z环,进一步吸引膜结合蛋白,通过直接或间接作用形成隔膜,然后引导整个分裂过程。它广泛分布于原核生物细胞内,其保守性高,已成为抗细菌抗生素筛选的理想靶标[2,26]。本研究实现了水稻白叶枯病菌FtsZ蛋白的体外表达,为进一步开发靶向抑制FtsZ蛋白生物活性、进而抑制细胞分裂的抗水稻白叶枯病菌的抗生素奠定了坚实的基础。
4 结论
本研究以水稻白叶枯病菌总DNA为模板,采用巢式PCR方法扩增获得水稻白叶枯病菌ftsZ基因,构建了ftsZ基因的表达载体pET-22b-ftsZ。表达载体
转化宿主E. coli BL21后,经IPTG诱导表达了FtsZ-6×His融合蛋白,并通过镍柱纯化获得了电泳纯的FtsZ-6×His融合蛋白。
[1]Fernando MG, Estefanía S, Jesús IM, et al. Molecular dynamics simulation of GTPase activity in polymers of the cell division protein FtsZ[J]. FEBS Letters, 2012, 586:1236-1239.
[2]Kapoor S, Panda D. Targeting FtsZ for antibacterial therapy:a promising avenue[J]. Expert Opinion on Therapeutic Targets, 2009, 13(9):1037-1051.
[3]González JF, Degrassi G, Devescovi G, et al. A proteomic study of Xanthomonas oryzae pv. oryzae in rice xylem sap[J]. Journal of Proteomics, 2012, 75:5911-5919.
[4]Mew TW, Alvare AM, Leach JE. Focus on bacterial blight of rice[J]. Plant Disease, 1993, 77(1):5-12.
[5]Zhu XF, Xu Y, Peng D, et al. Detection and characterization of bismerthiazol-resistance of Xanthomonas oryzae pv. Oryzae[J]. Crop Protection, 2013, 47:24-29.
[6]孙恢鸿.我国水稻白叶枯病菌致病力分化研究[J].植物保护, 2003, 29(3):5-8.
[7]朱清禾, 贾红华, 李艳, 等. Rhodococcus erythropolis手性醇脱氢酶的克隆表达及其性质[J]. 微生物学报, 2012, 52:83-89.
[8]李承刚, 商庆龙, 谷鸿喜, 等. pET21b-HPV16E4重组质粒的构建及鉴定[J].哈尔滨医科大学学报, 2007, 4(2):112-114.
[9]王光路, 夏俊刚, 谢希贤, 等.假交替单胞菌XM2107嘌呤核苷磷酸化酶基因克隆表达、重组蛋白纯化及酶学性质[J].微生物学报, 2010, 50(2):222-227.
[10]刘箭.生物化学实验教程[M].北京:科学出版社, 2004:94-101
[11]徐静静, 蔺宇, 董立明, 等.用SSR标记和巢式PCR快速检测大豆疫霉菌[J].中国农业科学, 2009, 42(5):1624-1630.
[12]Cho JH, Jeong KS, Han JW, et al. Mutation in clpxoo4158 reduces virulence and resistance to oxidative stress in Xanthomonas oryzae pv. oryzae KACC10859[J]. Plant Pathol J, 2011, 27:89-92.
[13]Li T, Huang S, Zhao X, et al. Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukaryotes[J]. Nucleic Acids Res, 2011, 39:6315-6325.
[14]王春莲, 章琦, 周永力, 等. 我国长江以南地区水稻白叶枯病原菌遗传多样性分析[J].中国水稻科学, 2001, 15(2):131-136.
[15]Yang SQ, Liu SY, Zhao S, et al. Molecular and pathogenic characterization of new Xanthomonas oryzae pv. oryzae strains from the coastline region of Fangchenggang city in China[J]. World J Microbiol Biotechnol, 2013, 29(4):713-720.
[16]Bhasin H, Bhatia D, Raghuvanshi S, et al. New PCR-based sequence-tagged site marker for bacterial blight resistance gene Xa38 of rice[J]. Molecular Breeding, 2012, 30:607-611.
[17]Feng JX, Cao L, Li J, et al. Involvement of OsNPR1 /NH1 in rice basal resistance to blast fungus Magnaporthe oryzae[J]. European Journal of Plant Pathology, 2011, 131(2):221-235.
[18]Zhu XF, Xu Y, Peng D, et al. Detection and characterization of bismerthiazol-resistance of Xanthomonas oryzae pv. Oryzae[J]. Crop Protection, 2013, 47:24-29.
[19]Davies J. How to discover new antibiotics:harvesting the parvome[J]. Curr Opin Chem Biol, 2011, 15(1):5-10.
[20]Yan RB, Yuan M, Wu Y, et al. Rational design and synthesis of potent aminoglycoside antibiotics against resistant bacterial strains[J]. Bioorg Med Chem, 2011, 19(1):30-40.
[21]Huigens-III RW, Reyes S, Reed CS, et al. The chemical synthesis and antibiotic activity of a diverse library of 2-aminobenzimidazole small molecules against MRSA and multidrug-resistant A. baumannii[J]. Bioorg Med Chem, 2010, 18(2):663-674.
[22]Fiedler HP, Bruntner C, Bull AT, et al. Marine actinomycetes as a source of novel secondary metabolities[J]. Antonie van Leeuwenhock, 2005, 87(1):37-42.
[23]Peric-Concha N, Long PF. Mining the microbial metabolome:a new frontier for natural product lead discovery[J]. Drug Discovery Today, 2003, 8(23):1078-1084.
[24]Heath RJ, Rock CO. Fatty acid biosynthesis as a target for novel antibacterials[J]. Current Opinion in Investigational Drugs, 2004, 5(2):146-153.
[25]Jain R, Chen D, White RJ, et al. Bacterial peptide deformylase inhibitors:a new class of antibacterial agents[J]. Current Medicinal Chemistry, 2005, 12(14):1607-1621.
[26]Singh P, Panda D. FtsZ inhibition:a promising approach for antistaphylococcal therapy[J]. Drug News & Perspectives, 2010, 23(5):295-304.
(责任编辑 马鑫)
Expression of FtsZ Protein from Xanthomonas oryzae in Escherichia coli
Chen Yang Huang Yunhong Li Suzhen Long Zhonger
(College of Life Science,Jiangxi Normal University,Nanchang 330022)
It was to prepare FtsZ protein using techniques of modern molecular biology. The ftsZ gene was amplified from Xanthomonas oryzae by nested PCR, and recombinant plasmid pET-22b-ftsZ was constructed and transformed to E.coli BL21. The clony fragment was identificatified by PCR screening, Nde I/Xho I digestion and DNA sequencing, the positive clones were induced by IPTG for expression;the fusion protein was purified through Ni-NTA Resin, and identified by SDS-PAGE and Western blotting. Results showed that the recombinant plasmid pET-22b-ftsZ was constructed successfully, the FtsZ-6×His fusion protein was expressed in recombined E. coli BL21 induced by IPTG, and purified through Ni-NTA Resin by electrophoretic purity.
FtsZ protein Xanthomonas oryzae Escherichia coli Genetic recombination
2014-04-09
国家自然科学基金项目(31160029,31360018),江西省自然科学基金项目(20122BAB204008,20132BAB204007)
陈洋,男,硕士研究生,研究方向:微生物药物学;E-mail:845487250@qq.com
龙中儿,男,博士,教授,研究方向:基础和应用微生物学;E-mail:Longzhonger@163.com
