三角帆蚌四核苷酸重复微卫星的开发及特征分析
2014-03-17韩学凯李家乐2王照旗白志毅
韩学凯李家乐,2王照旗白志毅
(1.上海海洋大学 农业部淡水水产种质资源重点实验室,上海 201306;2.上海高校水产养殖学 E-研究院,上海 201306)
三角帆蚌四核苷酸重复微卫星的开发及特征分析
韩学凯1李家乐1,2王照旗1白志毅1
(1.上海海洋大学 农业部淡水水产种质资源重点实验室,上海 201306;2.上海高校水产养殖学 E-研究院,上海 201306)
用磁珠富集法和生物素标记(ATAC)5为探针构建了三角帆蚌基因微卫星富集文库。利用PrimerSelect软件对得到的微卫星序列进行引物设计并合成得到43对引物,初筛后发现有20对引物可以扩增出稳定、清晰的目的产物带。荧光标记这20对引物,然后用36只洞庭湖三角帆蚌扩增,发现17对引物呈现多态性。17个微卫星位点等位基因数的范围为6-28。期望杂合度(He)、观测杂合度(Ho)的范围分别为0.677 6-0.946 4、0.638 9-1.000 0。多态信息含量(PIC)在0.630-0.929之间。统计分析结果显示,其中有12个微卫星位点满足Hardy-Weinberg平衡条件。与本实验室之前开发的二核苷酸重复微卫星相比,该17对四核甘酸重复微卫星具有更高多态性,且更易于采用聚丙烯酰胺凝胶电泳等传统方法分析基因型,有利于节约成本,从而为三角帆蚌群体遗传分析、亲子鉴定等技术提供了准确、价格低廉的微卫星标记。
三角帆蚌 微卫星 四核苷酸重复 多态性
三角帆蚌(Hyriopsis cumingii)是我国特有的淡水珍珠蚌,其产珠质量佳,所产珍珠占全世界淡水珍珠的90%以上[1-3]。近些年来,养殖三角帆蚌中出现的产珠质量差、个体变小等一系列问题,对珍珠的产量和效益都造成较大的影响[4]。因而筛选培育抗逆性好、生长性状佳、产珠质量优的品种,促进三角帆蚌养殖业持续健康发展,是目前三角帆蚌遗传育种研究的主要方向。
由于微卫星的突变率高,可产生大量多态性丰富的位点,如今已作为一种良好的分子标记广泛应用于个体识别、种质资源鉴定、生物遗传作图、遗传多样性分析等方面的研究[5,6]。关于贝类微卫星开发与应用研究,国外已见较多报道[7-10],国内对淡水贝类如三角帆蚌[11-14]、背瘤丽蚌(Lamprotula leai)[15]、褶纹冠蚌(Cristaria plicata)[16]和背角无齿蚌(Anodonta woodiana)[17]的微卫星研究也逐渐开展起来。但目前国内淡水贝类开发的微卫星标记主要是双核苷酸重复微卫星,用传统银染法检测 PCR 产物时,扩增产物常常会出现影子带,对于PCR产物长度的正确分析干扰较大。研究人类基因组发现,四碱基重复微卫星序列具有丰富的多态性,易于 PCR 扩增,具有明显的等位基因差异,极少出现影子带[18],故分型效果较二核苷酸微卫星要好。鲁翠云等[19]研究鲤鱼(Cyprinus carpio)后发现,在黑龙江鲤野生群体中,四核苷酸重复的微卫星标记表现出的多态性水平高于双核苷酸重复。同样,谭照君等[20]在研究鲢鱼(Hypophthalmichtysmolitrix)中不同核苷酸重复微卫星特征时,发现四核甘酸重复微卫星不但多态性更高,而且遗传稳定性更好,更加适合作为种群分析的工具标记。在我国特有淡水贝类三角帆蚌中,四核苷酸重复微卫星是否同样具有高度多态性及其特征如何,还未见到有关研究报道。本研究通过生物素标记的微卫星探针获得并筛选一些三角帆蚌四核苷酸重复微卫星位点,通过对其多态性初步分析,以期为三角帆蚌群体遗传分析、亲子鉴定技术、种质资源的保护等研究提供更多高质量的微卫星分子标记。
1 材料与方法
1.1 材料
本研究使用在洞庭湖采集到的三角帆蚌野生群体作为试验样本。
1.2 方法
1.2.1 三角帆蚌基因组DNA的提取及富集文库的构建 取三角帆蚌外套膜组织作为提取材料,用苯酚-氯仿提取法提取其基因组DNA。基因组DNA的酶切和富集文库的构建主要参照罗明等[14]和Li等[21]的方法。用RsaⅠ内切酶进行三角帆蚌基因组DNA的酶切,用试剂盒回收200-1 000 bp大小的片段用于文库构建。微卫星序列的富集采用生物素标记的(ATAC)5探针,具体操作步骤与罗明等[14]所述相同。1.2.2 微卫星引物设计 先用Sequencher软件去除载体及接头后,再使用SSRhunter软件来查找其中微卫星位点,之后用GeneQuest软件查找并去除冗余序列。最后用PrimerSelect软件在微卫星序列上进行引物设计。
1.2.3 多态性微卫星引物的初筛及荧光标记引物分析 引物设计好后,首先送上海迈浦生物科技有限公司合成普通引物,然后用3个三角帆蚌的基因组DNA作为模板来初筛这些引物。经PCR扩增反应后,用2%琼脂糖凝胶电泳检测分析扩增产物,从中挑选能够稳定扩增出条带且与设计引物片段大小相符的引物,经初步筛选的引物用来合成5'上游荧光引物(FAM、HEX)。从洞庭湖中采集36只野生三角帆蚌,取每只蚌的外套膜组织进行基因组DNA提取。用荧光引物对36个模板DNA进行PCR扩增,扩增后的荧光PCR产物送上海迈浦生物科技有限公司进行STR测序分析。
1.2.4 数据分析 用GeneMapper v4.0软件对公司送来的基因组扫描结果进行分析并估算等位基因大小,利用PopGen32软件计算以下参数:等位基因数(N)、有效等位基因数(A)、观测杂合度(Ho)、期望杂合度(He),并进行Hardy-Weinberg平衡检测(HWE)。用Cervus软件来分析多态信息含量(PIC)。
2 结果
2.1 多态性引物的设计与筛选结果
用Sequencher软件去除接头及载体序列后,利用PrimerSelect设计并合成引物43对。用3个三角帆蚌的基因组DNA为模板对这43对引物进行初步筛选,PCR扩增产物经过2%琼脂糖凝胶电泳检测得到20对稳定扩增、条带清晰并且大小相符的引物。对这20对引物的5'上游荧光引物分别用 FAM和 HEX 两种荧光基团来修饰,用36只洞庭湖野生三角帆蚌个体对合成的20对荧光引物进行PCR扩增。用GeneMapper v4.0软件对公司送来的基因组扫描结果进行分析后显示,合成的20对引物中17对均具有良好的多态性(引物信息见表1),并对这17
对引物多态性进行深入评价。
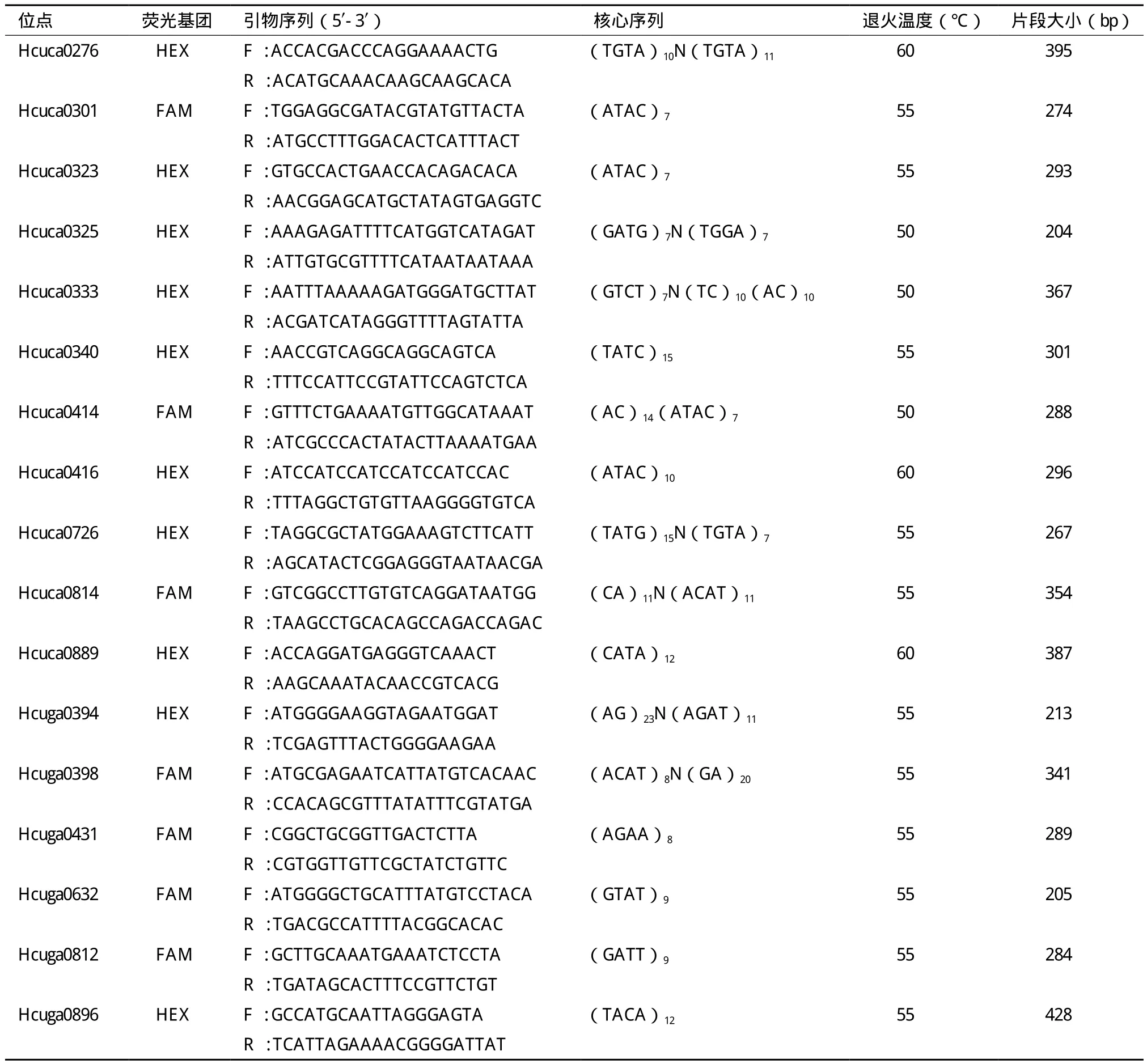
表 1 三角帆蚌 17个微卫星位点的核心序列及其引物特征
2.2 微卫星多态性引物对群体遗传评估分析
用PopGen32软件对这17对引物扩增的产物进行多态性评价分析,结果(图1)显示扩增产物大小在163-438 bp之间,17个微卫星位点等位基因数变动范围为6-28,平均每对引物获得等位基因数为17.52,其中位点Hcuca301获得等位基因数最少,仅为 6个,位点Hcuga726获得等位基因数最多,为28个,有效等位基因数在3.014 0-14.982 7之间,平均值为9.365 8,Ho在0.638 9-1.000,平均值为0.833 3;而He在0.677 6-0.946 4之间,平均值为0.877 9。用Cervus软件来分析多态信息含量,多态信息含量 PIC在0.630-0.929之间,平均值为0.853 1。经Hardy-Weinberg平衡检测显示,17个微卫星位点中有12个满足Hardy-Weinberg平衡条件(P>0.05)。17个微卫星位点多态性统计数据见表2。
3 讨论
虽然近些年微卫星标记在水产动物中得到大
量开发和应用,但是目前多数水产动物开发的微卫星标记多为双核苷酸重复。在有关人类二核苷酸重复微卫星研究中,发现PCR产物常常会出现比主带小2 bp的影子带[22,23]。本实验室利用二核苷酸重复微卫星开展三角帆蚌种质资源评价及其育种工作中也发现同样问题。研究发现,多核苷酸重复微卫星标记可以减少扩增产物中“影子带”的出现,比二核苷酸重复序列遗传稳定性更高且扩增更忠实[24],故基因分型效果较二核苷酸微卫星要好,且更易于采用聚丙烯酰胺等传统方法分析基因型。目前国内四核苷酸重复微卫星标记在水产动物中开发应用较少,仅见鲤鱼(Cyprinus carpio)[19]、鲢鱼(Hypophthalmichtysmolitrix)[20]、方正银鲫(Carassius auratus gibelioBloch)[25]、草鱼(Ctenopharyngodon idellu)[26]中有相关研究,而淡水贝类中关于四核苷酸的开发与研究却鲜有报道。而随着磁珠富集法的发展和完善,四核苷酸重复的微卫星筛选比例得到提高,并逐渐被开发应用。本研究通过构建三角帆蚌ATAC四核苷酸重复的富集文库,从中筛选和开发出17对四核甘酸重复微卫星标记用于研究。

表 2 三角帆蚌17个微卫星位点的遗传参数

图1 引物Hcuca0340在部分个体的扩增结果
多态信息含量(PIC)可以较好地反映片段的多态性和遗传信息含量。在一个群体中,多态信息含量高低与其提供的遗传信息多少成正比[27]。参照Botstein等[28]提出的衡量多态信息含量大小的指标,本研究中的17个微卫星位点的PIC在0.630-0.929之间,平均值为0.828 2,PIC值均大于0.5,为高度多态性位点,表明筛选微卫星位点多态性信息含量丰富。本实验室罗明等[14]曾用20对微卫星引物对洞庭湖野生三角帆蚌进行了遗传评估,本研究与其所用的洞庭湖样本为同一群体,并采用相同的分析方法,不同的是本研究使用的是四核甘酸重复微卫星,因此可以通过对两者结果进行对比,反映出双核苷酸重复与四核苷酸重复微卫星标记在多态性和遗传多样性评价方面差别(表3)。比较发现,本研究的四核苷酸重复微卫星多态信息含量(PIC=0.853 1)高于罗明用双核苷酸重复在同一群体中得到结果(PIC=0.823 9)。同样,与许巧晴等[13]用13个双核苷酸重复标记对洞庭湖野生三角帆蚌的
研究结果(PIC=0.519 8)相比,本研究四核苷酸重复微卫星评价的群体多态性也明显高。有效等位基因数反映了群体中等位基因的分布均匀度,而杂合度可以反映群体中遗传多样性高低。本研究17个微卫星位点在洞庭湖三角帆蚌野生群体扩增到的有效等位基因数(A=9.365 8)和期望杂合度(He=0.877 9)两项群体遗传多样性评估指标均高于罗明等用双核苷酸重复在同一群体得到的结果(A=6.582 2,He=0.823 9),表明四核甘酸重复评价的群体遗传多样性更高,遗传变异更大。与罗明和许巧晴研究结果相比,虽然本研究所用样本量稍多,但总体可以判断在三角帆蚌中四核苷酸重复微卫星多态性水平高于二核苷酸重复微卫星。此结果与谭照君等[20]通过研究鲢鱼和鲁翠云等[19]研究鲤鱼后得出结论相似。四核苷酸重复的微卫星多态性更高可能是真核生物中四核苷酸重复多存在于非编码区,在生物进化过程中承受的选择压力较小,因而可以较为“自由”突变的缘故[29]。
对17个微卫星位点进行Hardy-Weinberg平衡检测,发现其中有5个位点偏离了平衡,这可能是杂合子缺失或研究采用样本较少等缘故,这种情况在研究二核苷酸重复微卫星时也经常出现,属合理现象。选取本研究中开发的几对四核甘酸重复微卫星进行PCR扩增,通过传统银染法来检测PCR产物,发现扩增带谱清晰,“影子带”出现较少。这与Edwards等[18]研究人类中四核甘酸重复微卫星特征所得结果相一致。因此本研究开发的四核甘酸微卫星可作为理想的分子标记。

表3 不同类型核心序列的微卫星对洞庭湖三角帆蚌的评估效果比较
4 结论
本研究开发的17对四核苷酸重复的微卫星标记,不但具有高度的多态性,而且是较二核苷酸重复微卫星更为高效的分子标记,可以在不采用SSR荧光标记分析技术,节约成本情况下,用传统非荧光电泳检测方法同样取得较好效果,能够为三角帆蚌群体遗传分析、亲子鉴定技术等研究提供廉价、准确的分子标记,是很好的微卫星资源。
[1] Hua D, Gu R. Freshwater pearl culture and production in China[J]. Aquaculture Asia, 2002, 7(1):6-8.
[2] Li JL, Li YS. Aquaculture in China—Freshwater pearl culture[J]. World Aquaculture, 2009, 40(1):60-62.
[3] 刘月英, 张文珍, 王跃先, 等.中国经济动物志——淡水软体动物[M]. 北京:科学出版社, 1979:114-115.
[4] 张元培.淡水珍珠养殖技术[M].长沙:湖南科学技术出版社, 1981.
[5] Lagercrantz U, Ellegren H, Andersson L. The abundance of various polymorphic microsatellite motifs differs between plants and vertebrates[J]. Nucleic Acids Research, 1993, 21(5):1111-1115.
[6] 盛岩, 郑蔚红, 裴克全, 等.微卫星标记在种群生物学研究中的应用[J]. 植物生态学报, 2002, 26(s):119-126.
[7] Sato M, Kawamata K, Zaslavskaya N, et al. Development of microsatellite markers for Japanese scallop(Mizuhopecten yessoensis)and their application to a population genetic study[J]. Mar Biotechn, 2005, 7(6):713-728.
[8] Geist J, Geismar J, Kuehn R. Isolation and characterization of the first microsatellite markers for the endangered swan musselAnodonta cygneaL.(Bivalvia:Unionoidea)[J]. Conserv Genet, 2009, 11(3):1103-1106.
[9] Jones JW, Culver M, David V, et al. Development and characterization of microsatellite loci in the endangered oyster musselEpioblasma capsaeformis(Bivalvia:Unionidae)[J]. Mol Ecol Not, 2004, 4(4):649-652.
[10] Shaw KM, King TL, Lellis WA, et al. Isolation and characterization of microsatellite loci inAlasmidonta heterodon(Bivalvia:Unionidae)[J]. Molec Ecol Not, 2006, 6(2):365-367.
[11] Bai ZY, Yin YX, Hu SN. Identification of genes involved in immune response, microsatellite, and SNP markers from expressed sequence tags generated from hemocytes of freshwater pearl mussel (Hyriopsis
cumingii)[J]. Mar Biotechnol, 2009, 11(4):520-530.
[12] Li JL, Wang GL, Bai ZY. Genetic diversity of freshwater pearlmussel(Hyriopsis cumingii)in populations from the five largest lakes in China revealed by inter-simple sequence repeat(ISSR)[J]. Aquacult Int, 2009, 17(4):323-330.
[13] 许巧情, 谢骏, 张书环, 等.三角帆蚌微卫星富集文库的构建、鉴定及多态性分析[J].中国水产科学, 2010, 17(6):1200-1207.
[14] 罗明, 白志毅, 李应森, 等.三角帆蚌微卫星位点筛选及多态性分析[J].淡水渔业, 2012, 42(1):81-83.
[15] Xu B, Li JL, Wang GL. Development and characteriza tion of microsatellite loci inLamprotula leai, with cross-amplication inHyriopsis cumingii[J]. Conserv Genet Resour, 2011, 3(3):545-547.
[16] Jia MJ, Li JL, Wang SL. Isolation and characterization of 18 polymorphic microsatellite loci from freshwater pearl mussel(Cristaria plicata)[J]. Conserv Genet, 2010, 11(3):1131-1133.
[17] 汪桂玲, 苏翔, 李家乐, 等.背角无齿蚌基因组(GT)n微卫星DNA特征[J].生态学杂志, 2011, 30(1):1-6.
[18] Edwards A, Civitello A, Hammond HA, et al. DNA typing and genetic mapping with trimeric and tetrameric tandem repeats[J]. American Journal of Human Genetics, 1991, 49(4):746-756.
[19] 鲁翠云, 毛瑞鑫, 李欧, 等.鲤鱼三、四核苷酸重复微卫星座位的筛选及特征分析[J].农业生物技术学报, 2009, 17(6):979-987.
[20] 谭照君, 张天奇, 鲁翠云, 等.鲢三、四核苷酸重复微卫星标记的筛选及其特征分析[J].上海海洋大学学报, 2011, 20(3):329-335.
[21] Li JL, Wang GL, Bai ZY, et al. Ten polymorphic microsatellites from freshwater pearl musselHyriopsis cumingii[J]. Mol Ecol Notes, 2007, 7(6):1357-1359.
[22] Hauge XY, Litt M. A study of the origin of‘shadow bands’seen when typing dinucleotide repeat polymorphisms by the PCR[J]. Human Molecular Genetics, 1993, 2(4):411-415.
[23] Murray V, Monchawin C, England PR. The determination of the sequences present in the shadow bands of a dinucleotide repeat PCR[J]. Nucleic Acids Research, 1993, 21(10):2395-2398.
[24] Gastier JM, Pulido JC, Sunden S, et al. Survey of trinucleotide repeats in the human genome:assessment of their utility as genetic markers[J]. Human Molecular Genetics, 1995, 4(10):1829-1836.
[25] 曹柱, 鲁翠云, 邓先虎, 等.磁珠富集法筛选方正银鲫的三、四核苷酸重复微卫星 DNA[J].淡水渔业, 2011, 41(1):23-28.
[26] 李文升, 刘翠, 鲁翠云, 等.草鱼三、四核苷酸重复微卫星标记的分离与特征分析[J].中国水产科学, 2011, 18(4):742-750.
[27] 田艳苓, 毕伟伟, 杨海玲, 等.济宁百日鸡群体遗传多样性的微卫星标记分析[J].经济动物学报, 2010, 14(1):41-45.
[28] Botstein D, White RL, Skolnick M, et al. Construction of a genetic linkage map in man using restriction fragment length polymorphisms[J]. American Journal of Human Genetics, 1980, 32(3):314-331.
[29] 高焕, 孔杰.串联重复序列的物种差异及其生物功能[J].动物学研究, 2005, 26(5):555-564.
(责任编辑 马鑫)
Development and Characteristics of Tetranucleotide Repeat Microsatellite Loci in Hyriopsis cumingii
Han Xuekai1Li Jiale1,2Wang Zhaoqi1Bai Zhiyi1
(1. Key Laboratory of Freshwater Fishery Germplasm Resources, Ministry of Agriculture, Shanghai Ocean University, Shanghai 201306;2. Aquaculture Division,E-Institute of Shanghai Universities,Shanghai 201306)
A microsatellite-enriched library of the Hyriopsis cumingii was constructed using repeat-enrichment method with biotinlabeled oligo(ATAC)5and streptavidin coated magnetic beads. Forty-three pairs of primers designed by software PrimerSelect according to the sequences were synthesized for the pre-screening. Twenty pairs of primers which could amplify constant and legible expected DNA products were labeled by fluorescence. Seventeen microsatellite loci showed high levels in genetic polymorphism testing on 36 individuals sampled from Dongting Lake. The number of alleles at each locus ranged from 6 to 28. The expected and observed heterozygosities varied from 0.677 6 to 0.946 4 and from 0.638 9 to 1.000, respectively. The PIC value ranged from 0.630 to 0.929. Twelve microsatellite loci fitted to Hardy-Weinberg equilibrium. Compared with the previous dinucleotide repeat microsatellite developed in our laboratory, these 17 tetranucleotide repeat microsatellite loci showed higher polymorphism. Also, these 17 microsatellite loci can make genotyping easier by conventional methods such as polyacrylamide gel electrophoresis and save cost. Therefore, it offered accurate and inexpensive microsatellite markers for population genetic analysis and paternity test in Hyriopsis cumingii.
Hyriopsis cumingii Microsatellite Tetranucleotide repeat Polymophism
2013-12-16
上海市高校知识服务平台项目(ZF1206),国家科技支撑计划项目(2012BAD26B04),国家自然科学基金项目(31272657),上海市青年科技启明星(A类)(12QA1401400),上海工程技术中心能力提升项目(13DZ2280500)
韩学凯,男,硕士,研究方向:种质资源与遗传育种;E-mail:15618062362@163.com
白志毅,男,博士,副教授,研究方向:种质资源与遗传育种;E-mail:zybai@shou.edu.cn
