青花菜及其近缘种亲缘关系SRAP标记分析
2014-03-17荆赞革裴徐梨唐征张小玲罗天宽刘庆朱世杨
荆赞革裴徐梨唐征张小玲罗天宽刘庆朱世杨
(1.温州科技职业学院 浙南作物育种重点实验室,温州 325006;2.南京农业大学 作物遗传与种质创新国家重点实验室,南京 210095)
青花菜及其近缘种亲缘关系SRAP标记分析
荆赞革1裴徐梨2唐征1张小玲1罗天宽1刘庆1朱世杨1
(1.温州科技职业学院 浙南作物育种重点实验室,温州 325006;2.南京农业大学 作物遗传与种质创新国家重点实验室,南京 210095)
利用SRAP分子标记技术,对青花菜与其近缘种进行遗传多样性分析。28对SRAP引物共产生302条谱带,其中多态性谱带203条,多态率为67.22%,表明种质间存在较高的多态性。相似系数分析表明,其变异范围为0.461 5-0.900 6,平均遗传相似系数为0.693 6。‘绿地’和‘矮抗青’之间的亲缘关系最远,遗传相似系数为0.461 5;‘Wzvcst-09-224’和‘Wzvcst-09-225’亲缘关系最近,遗传相似系数为0.900 6。聚类分析可将16个材料分为两大类,第Ⅰ类包括芸薹属甘蓝种蔬菜,第Ⅱ类为芸薹属白菜种。揭示了青花菜及其近缘变种间具有部分相似的遗传基础,亲缘关系较近。结果表明,同一地域或来源的材料间具有较为相近的遗传背景,亲缘关系相对较近。研究结果有助于青花菜与其近缘种间种质资源分类和优异基因利用,加速青花菜新品种选育进程。
青花菜 SRAP 遗传多样性 亲缘关系分析
青花菜(Brassica oleraceaL. var.italica)别名西兰花、绿菜花,原产于地中海东部沿岸地区,是十字花科芸薹属甘蓝种中以绿或紫色花球为产品的一个变种。目前,青花菜已逐渐成为我国重要的出口创汇蔬菜之一,但其育种基础相对落后,多数青花菜种子仍依靠进口。随着青花菜产业的发展和生产需求,对青花菜新品种进行自主研发迫在眉睫。
优质种子种苗是蔬菜生产中极其重要的生产资料,近年来许多种子单位对青花菜开展了育种工作,大量优质新品种不断涌入市场。然而由于青花菜品
种间遗传背景狭窄,产品更新速度较慢。青花菜是由甘蓝演化而来,是野生甘蓝演化为木立花椰菜过程中出现的一个亚变种[1],同其它甘蓝变种间具有良好的杂交亲和性,因此研究青花菜及其近缘种之间的遗传多样性,促进优异基因转移,是进行青花菜新品种选育的重要方式之一。
常规的田间植株形态鉴定和同工酶鉴定都存在易受环境条件、所取材料部位等影响的缺点,而近年来发展起来的DNA分子标记技术,如随机扩增多态性DNA(RAPD)、ISSR、简单重复序列(SSR)、AFLP、相关序列扩增多态性(SRAP)等,以其快速、准确、不受环境条件影响等优点,现已成功应用于辣椒[2]、莼菜[3]、苦瓜[4]、黄瓜[5]、茄子[6]、萝卜[7]、大白菜[8]、西瓜[9]等多种作物,在种质资源鉴定[10,11]、遗传材料创建[12]、遗传多样性[13,14]、品种鉴定[15,16]等方面都得到了较好的应用。目前对青花菜品种及其近缘种亲缘关系分析方面的研究报道较少。本研究运用SRAP分子标记技术对青花菜及其近缘种进行品种鉴定与亲缘关系分析,旨在为青花菜与其近缘种间的利用与种质资源分类提供技术基础和理论依据,从而有助于优异近缘种基因转移,加速青花菜新品种选育进程。
1 材料与方法
1.1 材料
供试材料从浙江各地搜集,共15份甘蓝变种类蔬菜,包括10份青花菜品种和2份花椰菜地方品种,甘蓝、苤蓝、芥蓝各1份,以不结球白菜作为对照(表1)。试验材料种植于温州科技职业学院试验田中,常规田间管理。

表1 材料名称和编号
1.2 方法
1.2.1 基因组DNA的提取 取适量幼嫩叶片,液氮研磨粉碎后,采用参照植物基因组DNA提取试剂盒操作方法提取基因组总DNA。提取试剂盒(离心柱型)由TIANGEN生物技术有限公司(北京)生产。1.2.2 SRAP-PCR扩增与产物检测 试验选用了4个正向引物和12个反向引物(表2),由上海生工生物工程有限公司合成。以基因组DNA为模板进行PCR扩增,总反应体系为16 μL[17]:其中包括15 ng基因组DNA,2.0 mmol/L Mg2+,0.2 mmol/L dNTPs,1 UTaqDNA 聚合酶(TaKaRa),0.25 μmol/L引物。
PCR扩增程序为:94℃预变性5 min;94℃变性1 min,35℃复性1 min,72℃延伸1.5 min,5个循环;94℃变性1 min,50℃复性1 min,72℃延伸1.5 min,35个循环;72℃延伸7 min,低温保存。
扩增产物加4 μL上样缓冲液。采用6%变性聚丙烯酰胺凝胶,1×TBE 缓冲液,60 V预电泳30 min后,120 V恒压电泳1-2 h,至指示剂迁移至凝胶下
部时,结束电泳。采用AgNO3染色法检测电泳结果。

表2 用于遗传多样性分析的SRAP引物序列
1.2.3 数据处理与分析 对SRAP-PCR电泳图谱进行分析,统计清晰的SRAP位点,同一位置上出现谱带的记为“1”,未出现谱带的记为“0”。应用统计分析软件NTSYS pc version2.10e计算相似性系数,按UPGMA进行聚类分析,绘制聚类分析树状图。
2 结果
2.1 SRAP引物扩增多态性分析
从48对SRAP引物中筛选出了28对引物组合对16份材料基因组DNA进行了扩增,共产生了302个位点,平均每个引物扩增出10.79个位点。多态性位点共203条,平均多态性位点7.25个,多态率为67.22%,表明甘蓝变种间多态性高,遗传多样性较丰富,遗传背景也相对复杂。
2.2 遗传相似性分析
根据28对SRAP引物扩增结果,通过NTSYS pc version2.10e软件计算出品种间遗传相似系数(GSC)的变异范围为0.461 5-0.900 6,平均遗传相似系数为0.693 6。‘绿地’(3)和‘矮抗青’(16)之间的亲缘关系最远,遗传相似系数为0.461 5;‘Wzvcst-09-224’(11)和‘Wzvcst-09-225’(12)亲缘关系最近,遗传相似系数为0.900 6(表3)。
2.3 聚类分析
采用 UPGMA 方法进行聚类分析,得到16份材料的遗传聚类图(图1)。在遗传相似系数0.54处,可将供试材料分为2类:Ⅰ类包括10份青花菜(1-10)、2份花椰菜(11、12),甘蓝(13)、苤蓝(14)、羽衣甘蓝(15)各1份,共15份材料;Ⅱ类仅包括1个材料,即不结球白菜‘矮抗青’(16)。第Ⅰ类15份材料均属于芸薹属甘蓝种蔬菜,第Ⅱ类为芸薹属白菜种,与植物学分类结果一致。二者虽同属十字花科芸薹属,却为不同的种,亲缘关系相对较远。从形态上看,甘蓝变种植株相对高大,主根发达,叶片宽大,被有蜡粉,叶表面常呈灰绿色或蓝绿色;不结球白菜植株相对矮小,浅根系,须根发达。叶
片较小,叶色淡绿至墨绿,叶柄肥厚,白色或绿色。二者间植物学性状差异较大,不结球白菜与甘蓝变种间差异较大,分别形成独特的类群。在遗传相似系数为 0.67处,第Ⅰ类又可以分成2个亚组,第一亚组包括10份青花菜品种,第二亚组包括花椰菜、甘蓝、苤蓝和羽衣甘蓝。第一亚组中青花菜No.1-7大多来源于日本,聚为一个小组,表明同一地域来源的材料间具有较为相近的遗传背景。绿珍F1(9)和美好F1(10)来源于台湾长胜种苗股份有限公司,聚类在同一小组,推测可能采用了同一亲本或亲本来源相同;本研究结果表明同一来源或同一地理区域的品种间具有相对较近的遗传基础。第二亚组中Wzvcst-09-224(11)和Wzvcst-09-225(12)搜集于温州市洞头县,为松散型花椰菜地方品种,二者植物学性状相近,相似系数高,聚类在同一小组。表明二者间基因组信息相近,具有较低的遗传多样性。
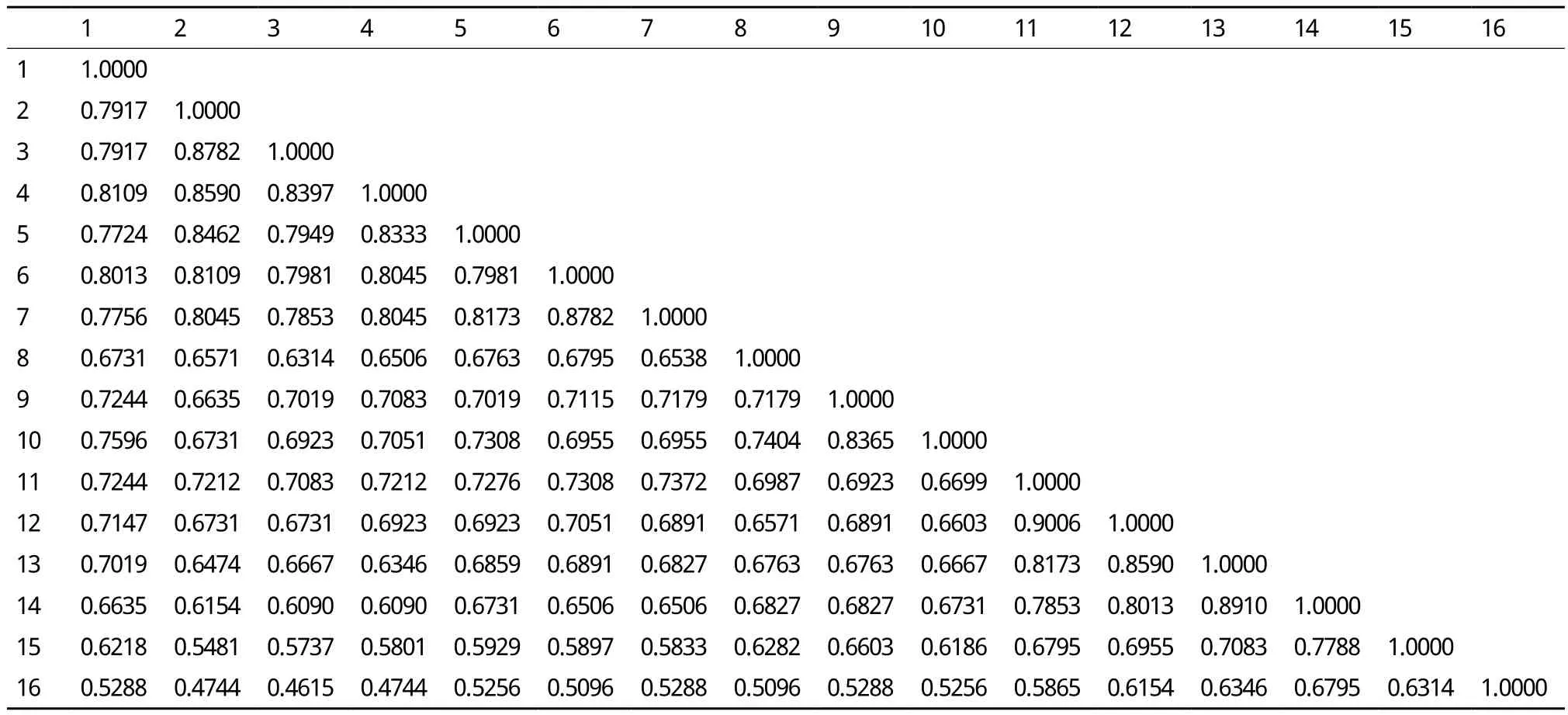
表3 16个种质间的相似系数

图1 基于SRAP标记的16份种质聚类图
3 讨论
本试验中28对多态性引物组合可检测到302个位点,其中多态性位点203个,多态率为68.26%。缪体云等[18]采用SRAP标记技术对甘蓝种质的遗传多样性和亲缘关系进行了分析,26个SRAP 引物组合可扩增出稳定清晰条带439条,其中多态性条带227条,多态性位点比率为51.7%。二者平均每个引物组合扩增条带数和多态性比例相差较大,推测由不同的引物组合和试验材料所引起。
甘蓝类蔬菜原产地中海沿岸和西北欧的海滨,先经甘蓝野生种(Brassico oleraceavar.oleracea)栽培驯化成羽衣甘蓝,而后分化出分枝细茎、髓状茎和高茎3个类型。结球甘蓝由不分枝类型的羽衣甘蓝分化而来,球茎甘蓝起源于髓状茎类型。分枝细茎类型进化成木立花椰菜,青花菜和花椰菜均为木立花椰菜的亚变种[1]。Song等[19]采用RFLP分子标记分析了芸薹属植物野生种和栽培种的遗传多样性,发现野生甘蓝独立于甘蓝栽培种聚为一类,推测甘蓝类作物可能是起源于某一种野生甘蓝。田源等[20]利用RAPD标记对甘蓝类蔬菜材料进行亲缘关系和遗传多样性分析。UPGMA分析结果可以将结球甘蓝、抱子甘蓝、羽衣甘蓝、青花菜、花椰菜、皱叶甘蓝清楚区分为6组,从分子生物学的角度分析验证了它们之间的亲缘进化关系。抱子甘蓝植物学形态特征特性与普通甘蓝相比差异明显,遗传多样性分析发现与结球甘蓝材料亲缘关系相对较远,独立聚类[8]。周禹等[21]对芥蓝、青花菜、结球甘蓝和球茎甘蓝及它们之间的24个杂交组合的主要植物学性状进行聚类分析,显示芥蓝与结球甘蓝的亲缘关系较近,推测芥蓝是甘蓝种的一个变种。
甘蓝变种中青花菜和花椰菜间的亲缘关系较近[22]。通过SSR标记可将结球甘蓝与青花菜区分开来,而青花菜与花椰菜却无十分严格的界线,表明花椰菜遗传多样性水平较低[23]。孙德岭等[24]利用AFLP标记对花椰菜、青花菜、紫花菜和黄花菜自交系的亲缘关系进行了研究,发现黄花菜、紫花菜和青花菜间的亲缘关系较近。本研究中聚类分析可将供试材料分为甘蓝种和白菜种两大类,表明甘蓝变种间具有部分相似的遗传基础,亲缘关系较近。10份青花菜与所选松散型花椰菜品种聚为不同的亚组,表明其之间具有较高的遗传多样性。
芸薹属植物的进化研究表明,其三大基本类群,白菜组最原始、芥菜组进化程度稍高、甘蓝组进化程度最高[25],本试验中通过NTSYS软件计算得出的品种间遗传相似系数中,‘绿地’(甘蓝组)和‘矮抗青’(白菜组)的遗传相似系数最小,亲缘关系较远。甘蓝组中的5种类型间遗传相似系数相对较大,表明它们存在特殊的进化关系。
遗传变异是蔬菜品种改良和选育的重要基础。蔬菜育种中,变种内变异往往不能充分满足新品种
对抗病性和抗逆性的需求,而通过杂交引入变种间或种间变异是非常重要的技术手段。大量研究表明,青花菜近缘种质资源中具有多种抗性、品质等优良基因,因此通过各种技术手段将这些优异因导入青花菜,可有效促进现有品种改良和新品种选育。
4 结论
利用SRAP分子标记技术,对青花菜与其近缘种进行了遗传多样性分析。28对SRAP引物共产生多态性谱带203条,多态率为67.22%,表明品种间存在较高的多态性。SRAP标记聚类分析表明,16份材料的相似系数变异范围为0.461 5-0.900 6,平均遗传相似系数为0.693 6。聚类分析可将试验材料分为二大类,第Ⅰ类包括芸薹属甘蓝种蔬菜,第Ⅱ类为芸薹属白菜种。表明青花菜及其近缘变种间具有部分相似的遗传基础,亲缘关系较近。聚类结果还表明同一地域或来源的材料间具有较为相近的遗传背景,亲缘关系相对较近。
[1] 刘英, 王超.简述甘蓝类植物的起源及分类[J].北方园艺, 2006(4):58-60.
[2] 陈学军, 程志芳, 陈劲枫, 等.辣椒种质遗传多样性的RAPD和ISSR及其表型数据分析[J].西北植物学报, 2007, 27(4):662-670.
[3] 刘朝贵, 李小孟, 须建, 等.三大产区莼菜遗传多样性及亲缘关系的RAPD分析[J].西南大学学报:自然科学版, 2012, 43(10):54-59.
[4] 李光光, 郑岩松, 李向阳, 等.利用SSR分子标记研究苦瓜资源的遗传多样性[J].南方农业学报, 2013, 44(1):6-11.
[5] 李锡香, 朱德蔚, 杜永臣, 等.黄瓜种质资源遗传多样性及其亲缘关系的AFLP分析[J].园艺学报, 2004, 31(3):309-314.
[6] 吴雪霞, 查丁石, 朱宗文, 许爽.茄子耐盐种质资源遗传多样性的SRAP和ISSR分析[J].植物生理学报, 2012, 48(8):789-794.
[7] 方平, 陈发波, 姚启伦, 等.肉质色不同萝卜遗传多样性的SSR分子标记分析[J].植物遗传资源学报, 2012, 13:226-232.
[8] 李停停.大白菜耐热性基因的SRAP分子标记[D].长春:吉林大学, 2011.
[9] 朱小茜, 孙治强, 李晓慧, 徐小利.西瓜全雌基因连锁的SRAP分子标记[J].河南农业科学, 2011, 40(3):105-108.
[10] 王彬, 郑伟, 范建新, 等.蔡永强早熟枇杷种质资源的RAPD分析[J].西南农业学报, 2012, 25(6):2231-2234.
[11] 张君玉.利用SSR和SRAP分子标记构建葡萄核心种质[D].洛阳:河南科技大学, 2012.
[12] 张慧, 张淑江, 李菲, 等.大白菜隐性细胞核雄性不育恢复基因BrMsf3的标记[J].中国蔬菜, 2011(4):13-16.
[13] 刘艺平, 李创, 李娜, 等.荷花遗传多样性的ISSR标记分析[J].西北农林科技大学学报:自然科学版, 2013, 41:139-146.
[14] 王玉安, 欧巧明, 陈建军, 等.甘肃地方杏品种资源的SSR遗传多样性分析[J].西北农业学报, 2013, 22(3):98-102.
[15] 王燕, 龚义勤, 赵统敏, 等.番茄SRAP-PCR体系优化与品种分子鉴定[J].南京农业大学学报, 2007, 30(1):23-29.
[16] 赵新, 王永, 兰青阔, 等.复合EST-SSR标记在大白菜品种鉴定中的应用[J].生物技术通报, 2013(1):107-110.
[17] 荆赞革, 唐征, 张小玲, 等.青花菜SRAP-PCR体系优化与品种分子鉴定[J].生物技术通报, 2010(12):122-125.
[18] 缪体云, 薄天岳, 陈锦秀, 任云英.甘蓝种质资源遗传多样性的SRAP分析[J].分子植物育种, 2010, 8(1):94-98.
[19] Song KM, Osborn TC, Williams PH.Brassicataxonomy based on nuclear restriction fragment length polymorphisms(RFLPs).3. Genome relationships inBrassicaand related genera and the origin ofB. oleraceaandB. rapa[J]. Theoretical and Applied Genetics, 1990, 79:497-506.
[20] 田源, 王超.甘蓝类蔬菜亲缘关系的RAPD初步分析[J].中国蔬菜, 2008(1):20-22.
[21] 周禹, 李燕, 孙勃, 等.芥蓝与甘蓝其他变种分类关系的研究[J].园艺学报, 2010, 37(7):1161-1168.
[22] Farnha MW. Genetic variation among and within united collard cultivars and Landraces as determined by randomly amplified Polymorphic DNA markers[J]. Journal of American Society for Horticultural Science, 1996, 121(3):374-379.
[23] Tonguç M, Griffiths PD. Genetic relationships ofBrassicavegetables determined using database derived simple sequence repeats[J]. Euphytica, 2004, 137(2):193-201.
[24] 孙德岭, 赵前程, 宋文芹, 等.花椰菜类蔬菜自交系基因组间亲缘关系的AFLP分析[J].园艺学报, 2002, 29:72-74.
[25] 王建林, 何燕, 栾运芳, 等.中国芸薹属植物的起源、演化与散布[J].中国农学通报, 2006, 22(8):489-494.
(责任编辑 马鑫)
Genetic Diversity and Relationship Analysis of Broccoli with Its Related Species by SRAP Markers
Jing Zange1Pei Xuli2Tang Zheng1Zhang Xiaoling1Luo Tiankuan1Liu Qing1Zhu Shiyang1
(1. Zhenan Key Laboratory of Crop Breeding,Wenzhou Vocational College of Science and Technology,Wenzhou 325006;2. State Key Laboratory of Crop Genetics and Germplasm Enhancement,Nanjing Agricultral University,Nanjing 210095)
In this study, we analyzed genetic diversity and relationship between broccoli and its related species by SRAP molecular markers. Using 28 SRAP primer combinations, a total of 302 amplified fragments were detected and 203 were polymorphic, with polymorphism rate 67.22%, indicating high polymorphism among these germplasm. Similarity coefficient analysis showed that the variation ranged from 0.461 5 to 0.900 6, and the average genetic similarity coefficient was 0.693 6. ‘Lü di’ and ‘Ai kang qing’ had the farthest genetic relationship with genetic similarity coefficient 0.461 5. By contrast, the relationship was closest between ‘Wzvcst-09-224’ and ‘Wzvcst-09-225’, with genetic similarity coefficient 0.900 6. Cluster analysis divided 16 germplasm into two major clusters. Class I contained Brassica oleracea, and class II only had one member Chinese no heading cabbage. The result of cluster indicated that the genetic basis had more similarity between broccoli and its related species, with a closer relationship, comparing with Brassica campestris ssp. chinensis Makino. The results also showed that the same geographic or origin could make the germplasm had relatively similar genetic background and closer genetic relationship. The research could be helpful to germplasm classification and excellent genes utilization for broccoli and its relative species, to speed up the breeding process.
Broccoli SRAP Genetic diversity Genetic relationship
2013-10-23
浙江省自然基金项目(LY12C15009),浙江省农业新品种选育重大科技专项(2012C12903-3-3),浙江省重大科技项目(2010C12004),温州科技局项目(N20090016)
荆赞革,男,博士研究生,助理研究员,研究方向:蔬菜遗传育种与生物技术;E-mail:jingzange@aliyun.com
唐征,男,副教授,研究方向:蔬菜遗传育种与生物技术;E-mail:tzeng05@163.com
