独流减河野生鲫鱼的同工酶分析
2014-02-18张文术张大莉
张文术,张大莉,郝 君,杨 蔷,董 仕
(天津师范大学a.生命科学学院,b.天津市动植物抗性重点实验室,天津300387)
鲫鱼属鲤形目(Cypriniformes)、鲤科(Cyprinidac)、鲫属(Carassius).我国鲫属鱼类有黑鲫(Carassius carassius)、鲫 (Carassius auratus auratus) 以及银鲫(Carassius auratus gibelio).依据形态、染色体数和繁殖特性将各地的鲫鱼分为野鲫、红鲫、方正银鲫等.在选育种方面,也已培育出多个养殖品种或杂交种[1-3].国内外学者应用同工酶、线粒体 DNA(mtDNA)、RAPD、微卫星DNA等遗传标记对野生鲫鱼、选育品种、鲤鲫杂交子代等进行了多方面的遗传分析[4-8].
同工酶由一个或多个基因座位编码,能够催化同一生化反应,在电场中的运动有多种可分离形式[9].同工酶分析能够揭示生物种群的遗传结构,识别从形态学上不容易区别的近缘种,也可检测种群的遗传多样性,分析物种之间的亲缘关系[10].检测行有性生殖二倍体生物的同工酶的遗传结构,可以计算出多态基因座位的比例、单位基因座位的等位基因数、平均杂合度观察值、平均杂合度预期值等遗传变异指标[11-12].鲫鱼中有染色体数为100条的二倍体种群和染色体数为150条左右的三倍体种群.对于三倍体鲫鱼,由于含有3套染色体组,每一基因座位有3个等位基因,依据同工酶电泳图谱难以判别基因型,同工酶分析一般只进行电泳条带差异性、组织表达特异性、克隆鉴别等研究[4,13-16].对于二倍体鲫鱼,罗莉中等[15]、王春元等[17]、汪亚平等[18]分别对金鱼的同工酶、洪湖和洞庭湖鲫鱼的同工酶进行了检测分析.作为水产养殖中重要的养殖种类以及杂交育种中重要的育种材料,对二倍体鲫鱼遗传结构的研究尤为重要.本课题组应用同工酶检测技术研究了采自天津市独流减河的野生鲫鱼同工酶的遗传变异情况,以期为鲫鱼资源的种质鉴定、保护利用以及遗传育种等提供基础数据.
1 实验材料与方法
1.1 材料
2012年11月在独流减河用刺网捕获鲫鱼47尾,鲜活状态下测量体长为(11.93±1.10)cm、体重为(58.52±14.60)g,从臀鳍后尾柄基部采血制作血涂片.-20℃冷冻保存备用.
1.2 红细胞大小的测量以及倍性判别
用吉姆萨染液对血涂片染色,在10×100高倍镜下测量红细胞的长径.依据文献[19]的方法进行鲫鱼倍数性的判别,标准为红细胞长径小于15μm的为二倍体鲫鱼,15~19μm为三倍体鲫鱼.
1.3 同工酶电泳检测
使用水平式淀粉凝胶电泳法检测独流减河鲫鱼肌肉和肝脏组织的同工酶,电泳及染色方法参照文献[20],使用 TBE(pH8.9)、C-T(pH8.0)、C-APM(pH6.0)3种缓冲液.采用文献 [21]的方法命名同工酶的缩写名、编号、基因座位、等位基因和基因型.
1.4 数据处理与分析
根据电泳结果判断每尾鱼每种同工酶的基因型,依据基因型计算每种同工酶的基因频率、多态基因座位数、多态基因座位比例、单位基因座位的等位基因数、平均杂合度观察值、平均杂合度预期值,并对有变异的基因座位进行哈代-温伯格平衡的χ2检验[12].
2 实验结果
2.1 红细胞的大小
实验测得独流减河野生鲫鱼红细胞的长径范围为11.16~14.42μm,平均为(12.45±0.71)μm,所有个体的红细胞长径均小于15μm,因此判定实验用鱼均为二倍体鲫鱼.
2.2 同工酶电泳结果
本研究检测到独流减河野生鲫鱼中的7种同工酶和肌浆蛋白,其种类、编号、基因座位等如表1所示.
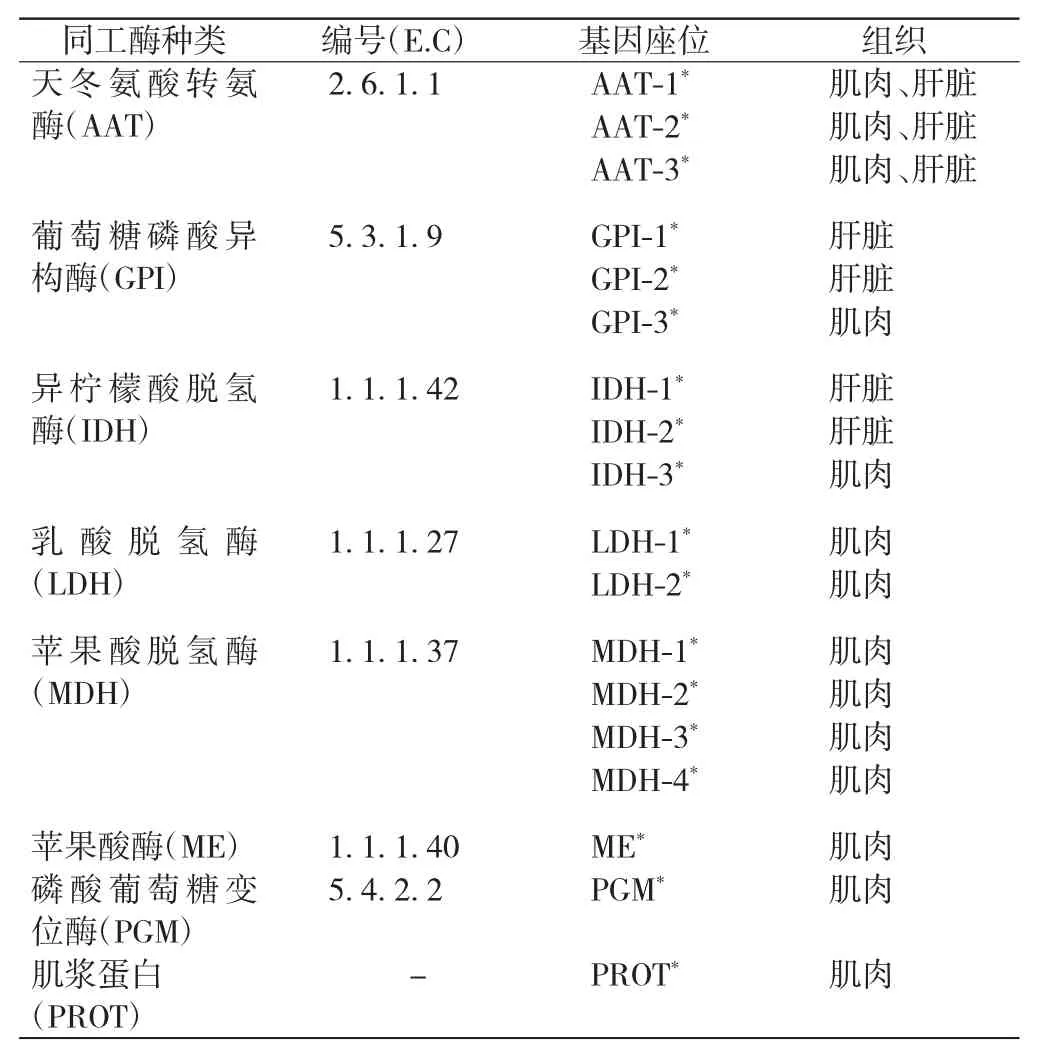
表1 检测到的同工酶种类、编号及基因座位Tab.1 Isozyme names,E C numbers and loci of crucian carp
由表1可以看出,7种同工酶和肌浆蛋白共判别出18个基因座位.其中MDH的基因座位数为4个,AAT、GPI、IDH各含3个基因座位,LDH的基因座位数为2个,其余3种同工酶的基因座位数为1个.在18个基因座位中,有变异的为8个,分别是AAT-3*、GPI-1*、GPI-2*、GPI-3*、IDH-2*、ME*、PGM*和PROT*,每个等位基因的基因频率如表2所示,部分同工酶和肌浆蛋白的电泳图谱如图1所示,每一条带下方为该条带的基因组成,由于GPI-1*和GPI-2*电泳条带较复杂,图中绘出其示意图.

表2 鲫鱼基因座位及其等位基因频率Tab.2 Loci and allele frequencies of isozymes of crucian carp
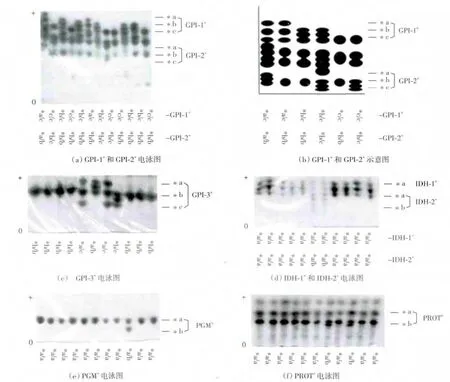
图1 鲫鱼同工酶电泳图谱Fig.1 Isozymes electrophoretogram of crucian carp
每个基因座位基因型的判别依据为:每种酶中每个条带出现的位置以及每个条带的分子构成.对于GPI而言,由于是二聚体酶,因此每条带都是两条相同或者不同的多肽链聚合而成.由表2和图1可以看出,同工酶GPI的3个基因座位均含有3个等位基因,AAT-3、IDH-2、PGM、PROT、ME含有 2个等位基因,其余的基因座位只有一个等位基因.
遗传变异结果中,群体的单位基因座位的等位基因数为1.611;如果按基因座位上最高基因频率小于0.99的为多态基因座位,则8个有变异的基因座位均为多态,多态基因座位比例为44.4%;如果按基因座位上最高基因频率小于0.95的为多态基因座位,则GPI-1*、GPI-2*、GPI-3*、ME*4个基因座位为多态,多态基因座位比例为22.2%;平均杂合度观察值(Ho)为0.0715,平均杂合度预期值(He)为0.0767,二者的比值为0.9322,接近于1.
对有变异的基因座位进行哈代-温伯格平衡的χ2检验,结果如表3所示,p值均大于0.05,符合哈代-温伯格平衡.

表3 鲫鱼群体内哈代-温伯格平衡的χ2检测Tab.3 χ2textfor departurefrom Hardy-Weinberg equilibrium in polymorphic isozyme locus of crucian carp
3 讨论与结论
同工酶作为遗传标记之一广泛应用于水产动物遗传育种学研究,如物种的分类系统学研究[22]、遗传多样性分析、同种不同群体遗传分化程度的研究[23]、河流中放流类型鱼类的混合比例分析[24]、种类的鉴别[25]、野生银鲫克隆的鉴别及亲子代的遗传特性分析[13-14]、鲤鲫杂交种的遗传分析[26]等.
鲫鱼是我国重要的淡水养殖鱼类之一,也是重要的育种材料.鲫鱼中有染色体数为100的二倍体种群和染色体数为150左右的三倍体种群.本研究通过红细胞大小判定采集自天津市独流减河的野生鲫鱼属于二倍体,再对其进行遗传多样性的检测分析.同工酶在鱼类的肌肉、肝脏、肾、心、脑、鳃等组织中经常会出现相同但电泳带迁移率不同或迁移率相同但电泳带浓度不同的现象,表现出组织特异性[23].本研究依据同工酶的电泳带数量、位置、浓度等可以看出二倍体野生鲫鱼同工酶的表达有明显的组织特异性,GPI-1*、GPI-2*、IDH-1*和 IDH-2*只在肝脏中表达,IDH-3*和GPI-3*只在肌肉中表达.
在应用同工酶检测技术分析鱼类的遗传多样性研究中,多态基因座位比例、单位基因座位的等位基因数、平均杂合度等是衡量遗传多样性的主要指标[11-12].其中平均杂合度是度量遗传变异的最简单直接、最富信息量的方法[11].一般认为贝类的平均杂合度为0.147,甲壳类的为0.043,鱼类的为0.059,水生哺乳类为0.034[27].Smith等[28]报道了89种海水鱼类的平均杂合度为0.060.汪亚平等[18]检测出洪湖和洞庭湖鲫鱼的平均杂合度分别为0.0702和0.1048.本研究分析了天津市独流减河二倍体野生鲫鱼的8种同工酶,检测出18个基因座位,其中8个有变异,平均杂合度观察值为0.0715,平均杂合度预期值为0.0767,高于以往研究中鱼类的0.059[27]以及89种海水鱼的0.060[28],这些结果表明天津市独流减河二倍体野生鲫鱼的遗传多样性较丰富.
[1] 伍献文.中国鲤科鱼类志[M].上海:上海人民出版社,1977:395-438.
[2] 俞豪祥.雌核发育与雄核发育//楼允东.鱼类育种学[M].北京:中国农业出版社,1999:153-194.
[3] 肖俊,罗永巨.我国鲫鱼研究进展[J].广西水产科技,2010(4):12-18.
[4]DONG S,TANIGUCHI N,TSUJI S.Identification of clones of Ginbuna Carassius langsdorfii by DNA fingerprinting and isozyme pattern[J].Nippon Suisan Gakkaishi,1996,62(5):747-753.
[5] 庄怡,刘必谦,唐杰.利用PCR-RFLR技术对鲫鱼6个不同品系的鉴定[J].生物技术通报,2012(11):144-149.
[6] 程磊,常玉梅,鲁翠云,等.鲫属鱼类DNA条码及种与亚种划分[J].动物学研究,2012,33(5):463-472.
[7] 程磊,刘洋,鲁翠云,等.淇河鲫雌核发育克隆系鉴定与性状分析[J].水产学杂志,2013,26(6):1-6.
[8] 郝君,杨蔷,鲍迪,等.6种mtDNA D-loop及其邻近区段的序列比较分析[J].大连海洋大学学报,2013,28(2):160-165.
[9] MARKERT C,MØLLER F.Multiple forms of enzymes:tissue,ontogenetie,and species specific patterns[J].Proceedings of the National Academy of Sciences USA,1959,45(5):753-763.
[10] 王辉.同工酶及其方法论[J].生物技术,1992,2(2):1-4.
[11]李思发,王强,陈永乐.长江、珠江、黑龙江鲢、鳙、草鱼原种种群生化遗传结构与变异//李思发,吴力钊,王强,等.长江、珠江、黑龙江鲢、鳙、草鱼种质资源研究[M].上海:上海科学技术出版社,1990:83-101.
[12]王中仁.遗传学计算//王中仁.植物等位酶分析[M].北京:科学出版社,1996:145-172.
[13]DONG S,TANIGUCHI N.Clonal nature of offsprings of Ginbuna Carassius langsdorfii by RAPD-PCR and isozyme patterns[J].Nippon Suisan Gakkaishi,1996,62(6):891-896.
[14]王茜,董仕,张学成.彭泽鲫繁殖特性的研究[J].南开大学学报:自然科学版,2001,34(4):103-106.
[15]罗莉中,毕世华,王春元.金鱼乳酸脱氢酶的同工酶的发生遗传学研究[J].遗传学报,1982,9(5):375-380.
[16]崔淼,赵俊,陈湘粦.三种鲫鱼品系同工酶比较研究[J].生态科学,2012,31(2):155-160.
[17]王春元,王长城.金鱼酯酶同工酶的研究Ⅰ:鲫鱼和红虎头金鱼各组织器官酯酶同工酶的比较[J].遗传学报,1988,15(6):442-449.
[18]汪亚平,张国华.鲫鱼种群间同工酶的比较研究[J].水生生物学报,1996,20(3):287-288.
[19]小野里坦,鳥沢雅,草間政幸.北海道に於ける倍数体フナの分布[J].魚類学雑誌,1983(30):184-190.
[20]TANIGUCHI N,OKADA Y.Genetic study on the biochemical polymorphism in red sea bream[J].Bulletin of the Japanese Society of Scientific Fisheries,1980,46(4):437-443.
[21]SHAKLEE J B,ALLENDORF F W,MORIZOT D C,et al.Gene nomenclature for protein-coding loci in fish[J].Transactions of the American Fisheries Society,1990,119:2-15.
[22]王可玲,张培军,刘兰英,等.中国近海带鱼种群生化遗传结构及其鉴别的研究[J].海洋学报,1994,16(1):93-104.
[23]张岩,高天翔,刘曼红,等.钝吻黄盖鲽同工酶组织特异性及群体遗传结构的初步研究[J].中国海洋大学学报:自然科学版,2007,37(2):235-242.
[24]TANIGUCHI N,DONG S,KONDO K,et al.Mixed ratio of the released landlocked form and distribution of the native amphidromous form of ayu,Plecoglossus altivelis in Yoshino river estimated by genetic markers[J].Susanzoshoku,2002,50(1):17-24.
[25]YOKOGAWA K,SEKI S.Morphological and genetic differences between Japanese and Chinese sea bass of the genus Lateolabrax[J].Japanese Journal of Ichthyology,1995,41(4):437-445.
[26]鲍迪,梁爱军,董莹.鲤鲫杂交子代的同工酶分析[J].水产科学,2012,31(5):283-287.
[27]藤尾芳久.水族における遺伝資源の存在様式と保全[M].日本仙台:かき研究所,1999:1-12.
[28]SMITH P J,FUJIO Y.Genetic variation in marine teleosts:high variability in habitat specialists and low variability in habitat generalists[J].Marine Biology,1982,69(1):7-20.
