不同株型观赏桃的生长及光合特性比较研究
2014-01-22刘卫东冯斌义梁文斌
谢 玲 ,刘卫东 ,冯斌义 ,梁文斌 ,马 英 ,袁 霞
(1.中南林业科技大学 林学院,湖南 长沙 410004;
2.湖南省绿林市政景观工程有限公司海口分公司,海南 海口 570100)
桃Prunus persica为蔷薇科李属桃亚属落叶小乔木,原产于我国,分布很广,主要分布在江苏、浙江、山东、河北等省,且各地的桃品种均有不同[1]。湖南常德市现有观赏桃花品种44个,其中本地品种有26个,从北京、郑州等地引种观赏桃品种18个[2]。张秀英于2000年完善了观赏桃的品种分类标准,并依据株型将其分为5大类:直枝桃、寿星桃、垂枝桃、杂种山桃、帚桃[3-4]。株型是观赏桃优质的基础,不同的株型对观赏桃的生理指标影响较大。长期以来,关于桃树株型的研究报道很少,而有关观赏桃株型的报道更是屈指可数。株型与光合作用密切相关,其中净光合速率是表征果树光合作用大小的直接指标[5]。本研究通过测定观赏桃5种不同株型叶片的生长量、光合参数、比叶重SLW、叶片叶绿素Chl.含量及其它因素与净光合速率Pn的关系,阐明不同株型观赏桃叶片的光合特性,旨在筛选出光合作用较强的观赏桃株型应用于生产,提高和扩大观赏桃的有效光合作用期,为完善观赏桃的栽培措施、加强科学管理、提高产品品质提供理论基础,同时也为观赏桃的推广应用提供参考依据。
1 材料与方法
1.1 试验地概况
试验地位于湖南省常德市双桥坪镇农夫堰村观赏桃花培育基地,地处中亚热带北部,地理坐标为29°29′N,111°80′E,属温暖湿润季风气候型,亚热带北缘湿润滨湖气候区;历年平均气温16.5℃,平均最高气温为20.8℃,平均最低气温为13.0℃,1月最冷,月均温度为4.0℃,7月最热,月均气温为28.7℃,≥10℃的活动积温为5 291.8℃。年降水1 240~1 700mm,主要集中在夏季;年日照时数1 466~1 700h,冬夏风向有明显变化。总的气候特点是:四季分明,光照充足,气候温和,雨量充沛,严寒期短,无霜期长。母岩主要为第四纪红色粘土,土壤为红壤,pH值为4.0~5.5。
1.2 试验材料
供试的观赏桃均为引种于试验地的2 a生正常植株,按株型分为5种:①直枝型:菊花桃、满天红、绛桃;②垂直型:鸳鸯垂枝、朱粉垂枝、黛玉垂枝;③寿星型:寿白、寿红、寿粉;④帚型:照手白、照手桃、照手姬;⑤杂种山桃:粉花山碧、粉红山碧。
1.2 试验方法
1.2.1 不同株型观赏桃生长指标测定
采用卷尺、直尺和游标卡尺测定植株的高度、地径、冠幅、叶片长度和宽度等。每个品种随机抽样15株进行测定。
1.2.2 不同株型观赏桃光响应曲线测定及光合参数拟合
2014年5月下旬至6月上旬,选择在晴天的上午8:00~11:30测定不同株型的不同品种观赏桃叶片的净光合速率Pn对光强的响应曲线[6]。每个品种随机选择1~3株植株,用美国Li-cor公司生产的Li-6400便携式光合仪测定。用光合测定仪测定朝南方向树冠外围上部生长良好的顶端第4~6片成熟叶片的净光合速率,每株测量3枚叶。测定时光强由强到弱,依次设定光合有效辐射(PAR)分别为2 000、1 800、1 500、1 200、900、600、300、200、150、100、75、50、25、0µmol·m-2s-1。对测得的数据拟合成回归方程,再根据回归方程计算各品种光合作用的表观光量子效率AQY、光补偿点LCP、光饱和点LSP、最大净光合速率P[7]。
光响应曲线拟合采用非直角双曲线模型(Cannell and Thornley,1998),其公式为:
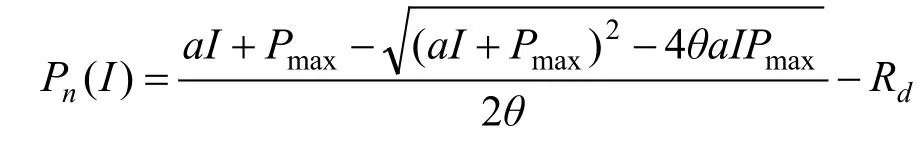
式中,Pn为净光合速率,I为光合有效辐射,a为植物光合作用对光响应曲线在I=0时的斜率(即光响应曲线的初始斜率,也称为初始量子效率),Pmax为最大净光合速率,θ为反映光响应曲线弯曲程度的曲角参数,取值0≤θ≤1,Rd为暗呼吸速率。根据模型拟合求出表观量子效率AQY、暗呼吸速率Rd和最大净光合速率Pmax。
1.2.3 叶绿素含量和比叶重的测定
于2014年6月上旬采集样叶,对5种不同株型的观赏桃,每个株型各选代表性的2~3个品种,其中每个品种各选3株,采用完全随机采样方式,每株选树体向南、生长一致的3个新梢的第5~6片树叶,按丙酮提取法[8]测定叶绿素含量。取0.2g新鲜样品用25ml 95%乙醇定容,分别用吸光度665、649nm测定吸光度,并计算总叶绿体素含量Tchl、叶绿素aChla和叶绿素bChlb的含量及叶绿素a与叶绿素b的比值a/b。
用叶面积仪测定叶片6~7片叶的面积,然后将叶片放在烘箱内烘干至衡质量,最后称取质量。比叶重=总叶质量(mg)/叶面积(dm2)。
1.3 数据统计与分析
采用Excel 2003进行数据处理,采用IBM SPSS Statistics 20进行光响应曲线拟合、显著性检验(LSD法)和绘图。
2 结果与分析
2.1 不同株型观赏桃生长的差异
测定的观赏桃均定植于2012年,在自然条件下生长,管理水平中等,开花结果正常,无明显病虫害发生。不同株型观赏桃的生长指标如表1所示,观赏桃不同株型之间的生长指标存在差异。植株高度方面,杂种山桃的植株生长值最高,极显著大于寿星型,超出其1.09倍,其与直枝型、杂种山桃、帚型之间无显著差异;植株冠幅大小以直枝型最大,与帚型、寿星型有极显著差异,分别大其1.25倍和1.86倍,直枝型、杂种山桃、垂枝型3种类型极显著大于帚型和寿星型,从冠幅大小可以直接分辨植株株型的紧凑和疏松,前者为疏松型树种,后者为紧凑型树种;地径大小以直枝型最大,直枝型地径大于寿星型1.69倍,直枝型与其他4类有极显著差异;叶片大小以寿星型最大,帚型的叶片最小,但各类型间均无显著差异。

表1 不同类型观赏桃树的生长指标†Table 1 Growth indexes of ornamental peach trees with different plant types cm
2.2 不同株型观赏桃叶绿素含量的差异
经过提取、分析和计算,测得了各类型叶片的叶绿素含量,并对各类型的叶绿素含量进行了方差分析。不同株型观赏桃的叶绿素含量如表2所示,5种类型叶片叶绿素总含量以帚型最高,平均值为2.64mg/g,叶绿素含量最低的为寿星型,平均值为1.97mg/g,但各类型观赏桃间无显著差异;叶绿素aChla含量以帚型桃为最高,平均值为1.60mg/g,含量最低的为寿星型,平均值为1.18mg/g,各类型间无显著差异;叶绿素bChlb含量为帚型最高,平均值1.04mg/g,含量最低的为寿星型,平均值为0.79mg/g,各类型间无显著差异;叶绿素a/b含量为杂种山桃最高,平均值为1.62mg/g,含量最低的为寿星桃型,平均值为1.50mg/g,各品种间无显著差异。

表2 不同株型观赏桃的叶绿素含量Table 2 Chlorophyll contents of ornamental peach trees with different plant types mg·g-1
2.3 不同株型观赏桃比叶重的差异
分别选取5种不同株型的观赏桃品种各3株,采用完全随机采取,每株选树体向南、生长一致的3个新梢第5~6片树叶,随机取4片树叶剪取一定面积进行烘干,直至恒重,测量每品种叶片的平均值,绘制图表。不同株型观赏桃的比叶重指标如图1所示,各株型的比叶重SLW含量不一,垂枝型>直枝型>杂种山桃>帚型>寿星型,其值在596±72~786±22mg/dm2范围内,用LSD方法进行显著性分析,各株型间的比叶重含量无显著差异,但株型较为疏松树种的比叶重值大于株型紧凑的树种。

图1 不同株型观赏桃的比叶重均值Fig.1 Mean speci fi c leaf weight of ornamental peach trees with different plant types
2.4 不同株型观赏桃光合参数的差异
有大量研究表明,枝条上叶片的叶位不同,光合能力也不相同,一般是测定第5至第7叶位叶片的光合能力[7]。在同一时间段内,测定了各品种的光合参数并进行方差分析,不同株型观赏桃的光合参数如表3所示。不同株型观赏桃之间的光合作用存在极显著差异,光合能力排序为:直枝型>杂种山桃>帚型>垂枝型>寿星型,直枝型观赏桃的最大净光合速率值Pmax极显著差异于寿星型并大于寿星型2.03倍,其他3个品种的平均值为11.29μmol/m2·s,直枝型与其它3个品种无显著差异。

表3 各株型观赏桃的光合参数Table 3 Photosynthetic parameters of ornamental peach trees with different plant types
根据Pn-PAR响应的回归方程(方程略),分别计算出了不同品种和不同株型桃的表观光量子效率AQY、最大净光合速率、光补偿点LCP及光饱和点LSP,结果如表3所示。杂种山桃和垂枝型桃的表观光量子效率较低与帚型桃有显著差异,直枝型和寿星型桃与其他3类型都无显著差异;直枝型和垂枝型桃的光补偿点和光饱和点较高与杂种山桃和帚型有极显著差异,说明直枝型和垂枝型桃对强光的利用效率较高,而杂种山桃和帚型桃具有较强的弱光利用能力,且较其它品种更容易在较低光强下达到最大净光合速率。
3 结论与讨论
3.1 不同株型对观赏桃生长指标的影响
杂种山桃、帚型与直枝型观赏桃株型较为高大,而垂枝型和寿星型较为矮小。其中垂枝型、直枝型和杂种山桃株型较为疏松,而帚型和寿星型较为紧凑。地径方面,直枝型极显著差异于其它4种株型植株,直枝型的生长速度较其它品种快,而帚型、寿星型的生长速度较慢。这与兴元、杨渺的研究结果相同[9-10]。叶片大小与株型的大小成反比(叶片越大,株型的体形越小),最矮小的寿星型叶片大于其它株型,而杂种山桃的叶片较小。周建涛研究表明观赏桃生长差异的出现与遗传因素有关[11]。文中试验结果表明,在相同环境下,株型较高大的杂种山桃和直枝型观赏桃的生命力较强。
3.2 不同株型对观赏桃叶绿素含量的影响
在光合作用的反应中吸收光能的色素成为光合色素,它很大程度上反映了植物生理状况和叶片的光合能力[12-13]。5种株型观赏桃的叶绿素含量无显著差异,相对而言,帚型和杂种山桃的叶绿素含量较高,寿星型的叶绿素含量较低,这与株型的高度成正比,其中帚型、杂种山桃和寿星型的叶绿素含量分别为2.64、2.53、1.97mg/g。说明株型的高矮与叶绿素含量有一定的联系,较高大的植物吸收光的能力强,叶绿素含量也就越高。
叶绿素a在红光区的吸收峰比叶绿素b的高,而蓝紫光区的吸收峰则比叶绿素b的低,也就是说,叶绿素b吸收短波长蓝紫光的能力比叶绿素a强,利用漫射光的能力强,吸收弱光的能力好。本研究中,寿星型叶绿素a/b比值最低,b的相对含量最高,在5种株型中耐阴性较强。说明株型的疏紧、叶片大小与植物吸收散射光的能力有一定的联系,株型越紧凑和叶面积越大,吸收短波长蓝紫光的能力越强,捕获弱光的能力越强,这与许慧风[14]等人的研究相同。虽然帚型观赏桃比垂枝型观赏桃高大,但前者的叶绿素a/b含量比后者低,说明在接受同样强度的光下,植株叶片中叶绿素a/b含量与植株高矮无关,与株型有关。株型越疏松越有利于叶片中叶绿素a的增加,相反,株型越紧凑越有利于吸收漫射光,从而增加叶片中叶绿素b的含量。
3.3 不同株型对观赏桃比叶重的影响
比叶重作为光合作用的一个生理指标,同时也是衡量叶片素质的一个稳定指标[15]。有研究表明,比叶重与植物叶片生长的光环境有很大关系,并与感受到的光强成正比。在强光下,叶片能通过增加光合能力、降低光合色素含量、增大比叶重以减弱光在叶肉组织中的传播[16],从而减少叶片对光照的捕获[17]。因此,生长在强光下的植物具有较厚的叶片和较高的比叶重。而在弱光下,叶片会通过增加光合色素的含量来增强捕光能力,利于碳的净积累[18]。较大的叶面积有利于叶片在弱光环境下捕捉到较多的光进行光合作用,这与弱光条件下叶片的比叶重较小的原理相同。寿星型和帚型观赏桃因株型紧凑,树冠内部吸收光能力较弱,其比叶重较低。而垂枝型、直枝型和杂种山桃因株型较疏松,利于对光线的吸收,比叶重较高,比叶重值与植株高矮无关。
3.4 不同株型对观赏桃光合参数的影响
植株的最大光合速率越大,表明能够达到的光合速率越大。光合速率的大小影响着光合作用的强弱和光能转化能力的大小。5种株型中较高大直枝型和杂种山桃的最大光合速率较高,光能转化能力较强,其中直枝型、杂种山桃分别为11.29μmol/m2·s、9.9μmol/m2·s。而株型较为矮小的垂枝型、寿星型的光能转化能力相对较弱,其中垂枝型和寿星型的分别为6.3μmol/m2·s、5.58μmol/m2·s。说明在正常生长情况下,光合作用、光能转化能力与植株的高矮成正比关系。
表观光量子效率又称低光强下的量子效率,主要反映了植物光合作用的生物物理特性,是表观光合作用中光能转化效率的重要参数,尤其是对弱光的转化效率。表观光量子效率大,表明植物吸收与转换光能的色素蛋白复合体可能较多,利用弱光的能力强[19]。本研究表明,帚型和寿星型观赏桃比其它株型的表观光量子效率较大,并与垂枝型和杂种山桃有显著差异,说明表观光量子效率与株型的高矮无关,与株型的疏松度有关。越紧凑的观赏桃株型利用弱光的能力越强。
植株光补偿点和饱和点的高低反映了植株对弱光的利用能力,是植株耐阴性评价的重要指标。植株的光补偿点越低,越能在弱光条件下顺利进行光合作用,利于有机物质的正向增长;光饱和点低则表明植株在有限的光条件下能最大利用低光量子密度,进行最大可能的光合作用[20-21]。帚型和垂枝型在光饱和点、光补偿点中,前者为最低后者为最高,但在表观光量子效率中两者却表现出相反的变化趋势。说明帚型、寿星型等紧凑型观赏桃在低光条件下仍能利用弱光,并与植株的高矮无关。
综上所述,幼年观赏桃植株中直枝型观赏桃的生长速度快,光合作用强,其次为杂种山桃和帚型观赏桃,而垂枝型、寿星型相对较弱,这可能与后者植株矮小相关。同时帚型和寿星型观赏桃比垂枝型、杂种山桃和直枝型更耐阴,这与株型的紧凑疏松成正比。在生产实际中,可根据不同株型的生理特性进行栽培管理。
[1] 曲泽洲,王永惠.桃生物学特性观察[J].河南农业大学学报,1964,2(3):25-39.
[2] 顾振华,王 燕,步洪凤,等.常德地区桃花品种资源调查[J].湖北农业科学,2009,48(2):376-378.
[3] 张秀英.中国名花丛书:桃花[M ].上海:上海科学技术出版社,2000.
[4] 张秀英,戴思兰,史历延.桃花种质资源多样性的研究[J].中国园林,1997,13(2):17 -19.
[5] GAO Z Q,Feng S Z,Zhang X C,et al.The simulation of leaf net photosynthetic rates in different radiation in apple canopy[J].Acta Ecologica Sinica,2012,32(4):1037-1044.
[6] 孟德悦,吴海勇,刘光斌,等.不同种源无患子光合与蒸腾速率差异分析[J].经济林研究,2013,31(2):48-53.
[7] 杨雨华,宗建伟,杨凤岭,等.3个核桃品种的光合-光响应特性[J].经济林研究,2014,2:19-23.
[8] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006.
[9] 兴 元,曹福亮,陈志明,等.遮荫对几种暖地型草坪成坪速度及其景观效果的影响[J].草原与草坪,2003,2:26-29.
[10] 杨 渺,毛 凯,马金星.遮阴生境下假俭草的形态变化与能量分配研究[J].中国草地,2004,26 (2):44-48.
[11] 周建涛,李惠芬.观赏桃主要性状遗传规律与国外育种实践[J].北京林业大学学报,1998,20(2):100-103.
[12] 汪景彦,杨有龙.苹果短枝型研究[M].北京:中国农业科技出版社,1987.
[13] 赵 坤,吴际友,陈 瑞,等.毛红椿光合及水分生理生态特性[J].中南林业科技大学学报,2011,5:87-91.
[14] 徐惠风,刘兴土,金研铭,等.向日葵叶片叶绿素和比叶重及其产量研究[J].农业系统科学与综合研究,2003,2:99-100.
[15] 陈雪芬.茶树病虫害防治[M].北京:金盾出版社,1996.
[16] Green D,Skkpuger E L.Light mediated constraints on leaf function correlate with leaf structure among deciduous and evergreen tree species[J].Tree Physiology,200121,1341-1346.
[17] Li Wei,Cao Kunfang.Effect of drought stress on photosynthetic characteristics and chlorophyll fluorescence parameters in seedlings ofTerminthia paniculatagrown under different light level[J].Acta Botanica Boreali-Occidentalia Sinica,2006,26(2):266-275.
[18] Kitao M,Lei TT,Koike T,et al.Susceptibility to photo inhibition of three deciduous broad leaf tree species with different successional traits raised under various light regimes[J].Plant,Cell and Enviornment,2000,23,81-89.
[19] LARCHER W.植物生理生态学[M].李 博,张陆德,岳邵先,等.北京:科学出版社,1982.
[20] 彭晓邦,蔡 靖,姜在民,等.渭北黄土区农林复合系统中大豆辣椒的光合生理特性[J].生态学报,2009,6:3173-3180.
[21] 李志辉,欧日明,王佩兰,等.6个仿栗家系光合特性的比较[J].中南林业科技大学学报,2014,9 (34):1-5.
