3个核桃品种的光合-光响应特性
2014-01-22杨雨华宗建伟杨风岭
杨雨华,宗建伟,2,3,杨风岭
(1.平顶山学院a.低山丘陵区生态修复重点实验室;b.资源与环境科学学院,河南 平顶山 467000;2.西北农林科技大学 林学院,陕西 杨凌 712100;3.中国林业科学研究院 林业研究所,北京 100091)
光合作用主要是由绿色植物叶绿素将太阳光能转化成化学能的复杂生理生化过程,它不仅受到植物自身因素的影响,而且受到环境生态因子的影响。光照是影响光合作用最主要的因子,植物与光环境的关系一直是科研工作者研究的热点问题[1-4]。随着光照强度的变化,植物可以通过自身形态及生理变化产生可塑性,以适应光环境[5]。植物光合作用的光响应曲线反映了光合有效辐射与植物光合速率之间的关系,研究植物光合作用过程中的光化学效率至关重要[6]。光响应曲线可以间接反映植物光合特性的相关生理参数,如最大(净)光合速率(Amax)、暗呼吸速率(Rd)、表观量子效率(AQY)、光饱和点(LCP)及光补偿点(LSP)等重要生理参数[7],这些参数能够反映植物光合作用机构是否正常运转,可用以判别植物在不同光照环境条件下的光适应能力及光合作用能力等[8]。因此,通过测定光合作用光响应曲线来确定植物光合生理生态特性,这已成为目前科研工作者研究的热点[9-12]。
核桃Juglans regiaL.属于胡桃科核桃属落叶乔木,其果实含有多种氨基酸、蛋白质、糖类物质及人体必需的多种矿质元素。此外,我国中医学者研究发现,核桃性温、味甘、无毒,具有补血养气、补肾填精、止咳平喘等功效,核桃中的精氨酸、油酸、抗氧化物质等对保护心血管、预防脑血管疾病大有裨益。因此,核桃是一种集食用、保健和药用功效于一体的坚果。过去,科研工作者对于核桃种质资源、生理生化、栽培管理等方面的研究已有一些报道[13-16],但对于核桃光合特性的研究报道却较少[17]。我国核桃品种丰富,研究和分析其生理变化特征,对于核桃的广泛种植及开发利用具有重要的现实意义。为此,文中测定了3个核桃品种的光合参数,分析其对不同光照强度的响应规律,以期为今后核桃的高产优质栽培提供一些生理学方面的科学依据。
1 材料与方法
1.1 试验地概况
试验地选在陕西杨凌西北农林科技大学林学院苗圃(108°07′E、34°12′N)内。杨凌位于陕西关中平原中部,属暖温带半湿润大陆性季风气候,海拔454.5m,年均降水量660mm,年均温12.9℃,年均积温4 811℃,年均日照时数2 163.8h,年均辐射总量114.8 kJ/m2,无霜期220d。
1.2 材料与方法
试验材料为‘鲁光’、‘扎343’和‘强特勒’这3个品种的1年生早实核桃嫁接苗。2011年3月上旬选择长势一致、无病虫害的3个品种的嫁接核桃苗,分别栽植于规格相同的塑料桶内,每桶1棵,培养基质为熟土∶营养土∶沙(3∶2∶1)的混合土。
试验选在2011年6月的晴朗天气进行,利用Li-6400便携式光合分析系统( LI-COR,Lincoln,USA)的自动光曲线程序,于上午10:30~11:30测定其光合-光响应曲线。每个品种各选取5 株作为待测植株,先经1h的光活化,然后每株选取当年生成熟叶片3片进行光响应曲线的测定。依据前期预试验的结果,光合有效辐射强度依次设定为0、20、50、100、150、200、250、300、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol·mm-2s-1。叶室选用红蓝LED光源,按照设定的光合有效辐射强度从小到大顺序控制光合有效强度,测定3个品种成熟片叶在不同光合有效辐射梯度下的净光合速率(Pn,μmol·mm-2s-1)、光合有效辐射(PAR,μmol·mm-2s-1)、胞间CO2浓度(Ci,μmol·mol-1)、气孔导度(Gs,mol·m-2s-1)、 蒸 腾 速 率(Tr,mmol·m-2s-1)、叶片水压亏缺值(Vapor pressure de fi cit at the leaf surface,即Vpdl,kPa),叶片温度(Tleaf,℃)等生理参数。光饱和点(LSP)、光补偿点(LCP)、最大净光合速率(Amax)、暗呼吸速率(Rd)和表观量子效率(AQY)等参数,这些参数均可采用Photosynthesis Work Bench 程序分析得到。
叶片瞬时水分利用效率(WUE,μmol·mmol-1)的计算公式为:Pn/Tr。
采用Excel软件处理试验中测定的所有数据。
2 结果与分析
2.1 3个核桃品种光响应曲线的特征参数
试验测定的3个核桃品种光响应曲线的特征参数如表1。由表1可知,3个不同核桃品种光响应曲线的特征参数间存在差异。在3个品种中,‘强特勒’的Amax、Rd、LSP和LCP值均达到了最高值,而‘鲁光’的AQY值最高。在一定环境条件下,叶片Amax反映了植物叶片的最大光合能力[18],而Rd与叶片的生理活性有关[19],AQY则反映了植物在弱光条件下的光合能力[20]。与‘鲁光’和‘扎343’相比,‘强特勒’对PAR的响应相对较快,其光合潜力最大,生理活性较强。

表1 3个核桃品种光响应曲线的特征参数Table 1 Characteristic parameters of light response curves of three J.regia cultivars μmol•m-2s-1
2.2 3个核桃品种光合速率随光强而变化的曲线
试验测得的3个核桃品种的光合-光响应曲线如图1 所示。由图1可知,3个不同核桃品种的Pn值均表现为先增后减的变化趋势,即起初随着PAR的增强而增大,且增幅较大,随后增幅减小,达到一定数值后,Pn值开始呈下降趋势。进一步分析3个核桃品种的Pn值随PAR值而变化的规律可以看出,在PAR值低于300 μmol·m-2s-1时,3个核桃品种的Pn值的差异不显著,均随着PAR的加强而上升,但‘鲁光’的Pn值略高于‘扎343’和‘强特勒’,这说明,在弱光条件下,‘鲁光’的光合能力要强于‘扎343’和‘强特勒’;当PAR值为300~1 000 μmol·m-2s-1时,与‘扎343’和‘强特勒’相比,‘鲁光’的Pn值最低,而‘扎343’和‘强特勒’两个品种的Pn值之间的差异较小,这说明,在中等强度的光强下,‘扎343’和‘强特勒’能够更好得利用光环境,其叶片具有较高的光合速率;当PAR值大于1 000 μmol·m-2s-1时,‘扎343’和‘鲁光’的Pn值的变化趋于平缓,而‘强特勒’的Amax值则显著高于前两者。进一步分析3个核桃品种的光响应曲线可知,‘鲁光’能够有效利用弱光环境,在弱光强下能够保持较高的光合速率,宜于发挥群体优势,可以密植;在强光环境下,‘强特勒’更容易发挥其较大的光合能力,宜于一般密度的种植;‘扎343’的整体表现介于‘鲁光’和‘强特勒’之间。

图1 3个核桃品种的光合-光响应曲线Fig.1 Light response curves of photosynthesis in three J.regia cultivars
2.3 3个核桃品种其它光合参数随光强而变化的曲线

图2 3个核桃品种的蒸腾速率和叶片水压亏缺随光强而变化的曲线Fig.2 Change curves of transpiration rate and vapor pressure de fi cit at leaf surface of three J.regia cultivars with light intensity increasing
3个核桃品种各光合参数随光强而变化的曲线分别如图2~4所示。从图2~4中可以看出,Tr和Vpdl值均随着PAR的增强而逐渐增大。其原因在于,光强的增强导致了空气湿度的降低,从而增大了叶片与大气间的水势差,致使Tr值增加[19];而叶片内水势下降,使得Vpdl值升高。Gs和Tleaf值同样随着PAR的增强而增大,Ci的变化趋势则是随着PAR的增强而迅速减小,当PAR值达到300 μmol·m-2s-1时,Ci值最小,随后略有升高,且渐趋于平缓。其原因在于,前期Gs的增幅较小,Amax值不断提高,使得Ci急剧下降,CO2的供应满足不了Amax的需求,表现出气孔限制现象;后期光强增强,对气孔的刺激也加强,导致气孔开放,Gs增幅较大,Ci表现出升高的趋势[19]。3个核桃品种其生理活性对PAR的适应性不尽相同。‘鲁光’在弱光的刺激下,可使气孔很快地打开,使Tr迅速增加,从而使Pn值快速提高;在高光强下,‘强特勒’的Gs值高于‘鲁光’和‘扎343’,其原因可能是,在高光强条件下,‘强特勒’的Pn值较高。水分利用效率取决于植物的光合速率与蒸腾速率。3个核桃品种的WUE值均随着PAR的增强而呈现出先增后降的变化趋势,‘鲁光’和‘扎343’的WUE值在光强为250 μmol·m-2s-1时达到了最大,而‘强特勒’的WUE值在光强为300 μmol·m-2s-1时达到最大。
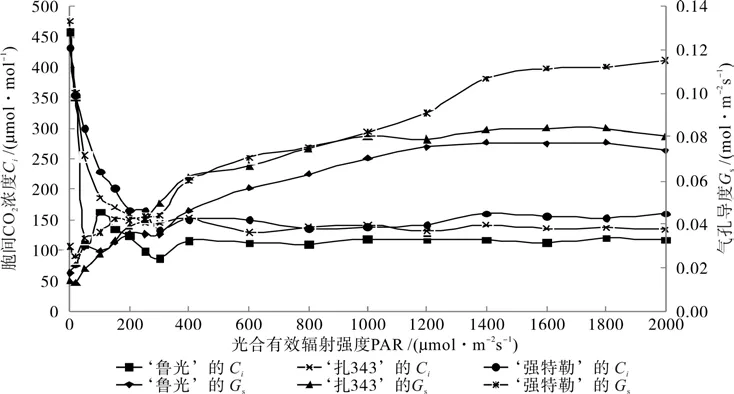
图3 3个核桃品种胞间CO2浓度和气孔导度随光强而变化的曲线Fig.3 Change curves of intercellular CO2 concentration and stomatal conductance of three J.regia cultivars with light intensity increasing

图4 3个核桃品种叶片温度和水分利用效率随光强而变化的曲线Fig.4 Change curves of leaf temperature and water use ef fi ciency of three J.regia cultivars with light intensity increasing
3 结论与讨论
LSP和LCP是植物的两个重要的光合生理指标。植物叶片的LSP与LCP可以反映植物对光照条件的要求,LSP值较低且LCP值越低,表明其对弱光的利用能力越强;而LSP值越高,则表明其对强光的利用能力越强,即LCP值较低而LSP值较高的植物对环境适应性很强;LCP值较高而LSP值较低的植物对光照的适应范围相对较窄[21-22]。文中对3个核桃品种的光合-光响应曲线的分析结果表明,不同核桃品种的光响应特征参数之间存在差异。‘强特勒’的LCP和LSP值均高于‘扎343’和‘鲁光’,这在一定程度上说明了‘强特勒’对环境的适应性要弱于‘扎343’和‘鲁光’。Amax是衡量群体光合能力的重要指标[23],在3个核桃品种中,‘强特勒’的Amax值最大,说明‘强特勒’的光能利用能力最强。从光合生产角度来看,在高光强下,‘强特勒’能够充分利用光能,宜于一般密度的栽培;‘鲁光’的AQE值最高,表明其在弱光环境下可以有效利用光能,适宜密植;‘扎343’则宜于适度密植。目前,高光效育种已经越来越受到科学研究者的重视,而在遗传特性方面寻找具有高光合效率的新品种尤为有关科研工作者关注。
[1] Awada T,Radoglou K,Fotelli M N,et al.Eco physiology of seedlings of three Mediterranean pine species in contrasting light regimes [J].Tree Physiology,2003,23: 33-41.
[2] Moreno-Sotomayor A,Weiss A,Paparozzi E T,et al.Stability of leaf anatomy and light response curves of fi eld grown maize as a function of age and nitrogen status [J].Journal of Plant Physiology,2002,159: 819-826.
[3] 张小全,徐德应.18年生杉木不同部位叶龄光响应研究[J].生态学报,2001,21(3): 409-404.
[4] 陈根云,俞冠路,陈 悦,等.光合作用对光和二氧化碳响应的观测方法探讨[J].植物生理与分子生物学报,2006,32(6):691-696.
[5] 钱莲文,张新时,杨智杰,等.几种光合作用光响应典型模型的比较研究[J].武汉植物学研究,2009,27(2):197-203.
[6] Robert E S,Mark A,John S B.Kok effect and the quantum yield of photosynthesis [J].Plant Physiology,1984,75:95-101.
[7] 叶子飘,康华靖.植物光响应修正模型中系数的生物学意义研究[J].扬州大学学报(农业与生命科学版),2012,32(2):51-57.
[8] 王荣荣,夏江宝,杨吉华,等.贝壳砂生境干旱胁迫下杠柳叶片光合光响应模型比较[J].植物生态学报,2013,37(2):111-121.
[9] 周玉梅,韩士杰,张军辉,等.不同CO2浓度下长白山3种树木幼苗的光合特性[J].应用生态学报,2002,(1):41-44.
[10] Damesin C.Respiration and photosynthesis characteristics of current-year stems of Fagus sylvatica from the seasonal pattern to an annual balance[J].New Phytologist,2003,158: 465-475.
[11] Mao Zijun,Wang Yanjun,Wang Xiuwei,et al.Effect of Doubled CO2on Morphology: Inhibition of Stomata Development in Growing Birch(Betula platyphyllaSuk)Leaves[J].Russian Journal of Plant Physiology,2005,52:198-202.
[12] 邱 权,潘 昕,李吉跃,等.速生树种尾巨桉和竹柳幼苗的光合特性和根系特征比较[J].中南林业科技大学学报,2014,34(1): 53-59.
[13] 马和平,朱雪林,刘务林,等.西藏核桃种质资源研究[J].果树学报,2011,(1):151-155.
[14] 黄少勇,张智俊,罗淑萍,等.山核桃EST-SSR引物的筛选及通用性分析[J].经济林研究,2013,31(3):10-15.
[15] 高 英,董宁光,张志宏,等.早实核桃雌花芽分化外部形态与内部结构关系的研究[J].林业科学研究,2010,(2):241-245.
[16] 张 瑞,李永荣,彭方仁.薄壳山核桃品种“马汉”的栽培适应性及其评价[J].经济林研究,2013,31(2):176-180.
[17] 宗建伟,杨雨华,杨风岭,等.四个核桃品种光合日变化的比较研究[J].草业科学,2013,30(3):397-401.
[18] 陆佩玲,罗 毅,刘建栋,等.华北地区冬小麦光合作用的光响应曲线的特征参数[J].应用气象学报,2000,11(2):236-241.
[19] 郭 江,郭新宇,王纪华,等.不同株型玉米光响应曲线的特征参数研究[J].西北植物学报,2005,25(8):1612-1617.
[20] 蹇洪英,邹寿青.地毯草的光合特性研究[J].广西植物,2003,23(2): 181-184.
[21] 杨秀芳,玉 柱,徐妙云,等.2种不同类型的尖叶胡枝子光合-光响应特性研究[J].草业科学,2009,26(7):61-65.
[22] 王旭军,潘百红,程 勇,等.红榉不同种源光合特性的比较[J].中南林业科技大学学报,2013,33(6):38-42.
[23] 谭晓红,彭祚登,贾忠奎,等.不同刺槐品种光合光响应曲线的温度效应研究[J].北京林业大学学报,2010,32(2):64-68.
