氯化锦葵色素-3-半乳糖苷对TNF-α诱导内皮细胞炎症的抑制作用
2014-01-20黄午阳李春阳郑其升刘娅梅
王 健,黄午阳,李春阳,郑其升,刘娅梅
氯化锦葵色素-3-半乳糖苷对TNF-α诱导内皮细胞炎症的抑制作用
王 健1,2,黄午阳1,*,李春阳1,郑其升3,刘娅梅3
(1.江苏省农业科学院农产品加工研究所,江苏 南京 210014;2.南京农业大学食品科技学院,江苏 南京 210095;3.江苏省农业科学院 国家兽用生物制品技术研究中心,江苏 南京 210014)
目的:研究氯化锦葵色素-3-半乳糖苷对细胞肿瘤坏死因子(tumour necrosis factor-α,TNF-α)诱导人脐静脉内皮细胞(human vascular umbilical endothelial cells,HUVEC)所产生炎症的抑制作用。方法:设立分组,空白组(二甲基亚砜)、10 μg/L TNF-α刺激组、浓度为1、10、50、100 μmol/L氯化锦葵色素-3-半乳糖苷+10 μg/L TNF-α刺激组,通过酶联免疫吸附剂测定方法检测细胞上清中可溶性细胞间黏附因子(intercellular cell adhesion molecule-1,ICAM-1)蛋白含量的变化;用实时荧光定量聚合酶链式反应检测细胞中ICAM-1 mRNA表达量变化;Western blotting检测ICAM-1及核因子κB抑制蛋白(IκBα)蛋白含量的变化;通过免疫荧光方法测定核转录因子κB(nuclear transcription factor-κB,NF-κB)核异位的变化。结果:氯化锦葵色素-3-半乳糖苷呈浓度依赖性的抑制TNF-α诱导HUVEC相应蛋白表达,同时结果显示抑制作用与NF-κB信号通路有关。结论:氯化锦葵色素-3-半乳糖苷抑制TNF-α诱导HUVEC的炎症反应。一般心血管疾病产生的原因中包括因促炎症因子引起的内皮功能障碍及产生的炎症,氯化锦葵色素-3-半乳糖苷的抗炎症作用为防治心血管疾病提供了一定的理论基础。
氯化锦葵色素-3-半乳糖苷;人脐静脉内皮细胞;肿瘤坏死因子;炎症反应;细胞间黏附因子;核转录因子
细胞炎症引起的内皮细胞损伤可以促使白细胞及单核细胞聚集,继而黏附于血管内壁引发血管功能障碍[1]。细胞间黏附因子(intercellular cell adhesion molecule-1,ICAM-1)主要表达于活化的T细胞、血管内皮细胞等表面,在正常情况下很少表达或不表达,当受到细胞因子刺激时表达明显增加[2]。促炎症细胞因子(TNF-α、IL-6、Ang-Ⅱ)可以诱导内皮细胞表达相应蛋白,能显著增强血管内皮细胞ICAM-1蛋白和mRNA的表达,促进炎症发生及发展进程[3-4]。
抗氧化物质对氧化应激引起的血管内皮系统损伤具有防治作用[5-6]。花青素是迄今为止所发现的最有效的天然水溶性抗氧化剂。蓝莓作为功能性食品,其果实中花青素含量很高而且种类丰富,具有抗炎症、增强心脏功能、改善循环、防止脑神经衰老、明目及抗癌等独特功效[7]。研究也发现抗氧化物质花青素中重要组分如锦葵色素-3-葡萄糖苷及白藜芦醇具有抗动脉粥样硬化作用[8-9]。
对于蓝莓花青素的提取分离及抗氧化能力方面研究甚多,但在分子基础研究蓝莓花青素抗炎症的很少,而野生种蓝莓中各花青素中锦葵色素的含量最高[10]。本研究以氯化锦葵色素-3-半乳糖苷为对象,研究其在内皮细胞内抑制炎症因子的作用。
1 材料与方法
1.1 材料与试剂
3~6代人脐静脉内皮细胞 江苏省农科院国药中心保存;氯化锦葵色素-3-半乳糖苷 美国Sigma公司;ICAM-1 ELISA试剂盒、兔抗人ICAM-1一抗、鼠抗人β-actin一抗、羊抗兔IgG、羊抗鼠IgG 博士德公司;兔抗人IκBα一抗、NF-κB核转运试剂盒 碧云天公司;Trizol、逆转录试剂盒、SYBR Premix Ex Taq试剂盒 日本TaKaRa公司。
1.2 仪器与设备
YDS-10B液氮罐 四川西亚低温设备有限公司;Airtech超净工作台 苏州安泰空气技术有限公司;5% CO2细胞培养箱 美国Thermo Electron公司;Axiovert-40C倒置显微镜、荧光显微镜 德国蔡司公司;TDZ4-W5小型离心机 上海启湘仪器有限公司; BG25金属浴 明基公司;Western blotting电泳仪 美国Bio-Rad公司;JY-300C转膜仪 北京君意仪器公司;酶标仪Awareness技术公司;凝胶成像系统 美国UVP公司;PCR仪 日本TaKaRa公司。
1.3 方法
1.3.1 细胞培养、传代[11]
复苏液氮冻存的3~6代人脐静脉内皮细胞,37℃融化,1 000 r/min离心3 min,吹打细胞,37℃,5% CO2培养箱培养24 h换液,继续培养待细胞长至80%~90%,进行传代。
1.3.2 样品制备
取1.891 mL二甲基亚砜(dimethyl sulfoxide,DMSO)溶解1 mg的氯化锦葵色素-3-半乳糖苷制备成1 μmol/mL的原液,根据要求分别稀释到最终浓度为1、10、50、100 μmol/L。
细胞进行6 孔板培养,设立分组:空白组 (DMSO)、TNF-α (10 μg/L)刺激组、浓度为1、10、50、100 μmol/L氯化锦葵色素-3-半乳糖苷(malvidin-3-galactoside chloride,Mv-3-gal-Cl)+10 μg/L TNF-α刺激组,预先加入不同质量浓度的氯化锦葵色素-3-半乳糖苷刺激18 h,再加入10 μg/L TNF-α刺激6 h。
1.3.3 酶联免疫吸附测定(enzyme linked immunosorbent assay,ELISA)法测定细胞上清可溶性ICAM-1蛋白含量[12]
细胞经不同分组药物刺激,取细胞上清,按一定比例稀释,根据ELISA试剂盒进行操作,在酶标仪450 nm波长处测定OD值。
1.3.4 Western blotting测定细胞中ICAM-1及IκBα蛋白含量[13]
弃6 孔板培养液,裂解细胞得到蛋白样品,12%十二烷基磺酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)进行电泳。PVDF膜进行湿转1.5 h,取膜37 ℃封闭、孵育一抗各1 h,TBST洗5 次,每次6 min, 37℃孵育二抗1 h,TBST洗涤,DAB显色,拍照分析数据。
1.3.5 实时荧光定量PCR测定细胞中ICAM-1 mRNA含量
在GenBank查询到目的基因β-actin、ICAM-1的mRNA序列,根据引物原则设计引物。β-actin:正向引物5’-CCACAGTCACCT-ATGGCAAC-3’和反向引物5’-AGTGTCTCCT GGCTCTGGTT-3’;ICAM-1:正向引物5’-CCACAGTCACCT-ATGGCAAC-3’和反向引物5’-AGTGTCTCCT GGCTCTGGTT-3’。弃6孔板培养基进行细胞总RNA提取[14-15],根据PrimeScript RT Master Mix配制反应体系,37 ℃,15 min,85 ℃,5 s;继而PCR反应,根据琼脂糖电泳检验cDNA及引物质量;根据SYBR Real Time PCR 反应试剂盒配制PCR反应液,用所有的测试样本和校准样本,采用2-’Ct计算细胞中ICAM-1 mRNA表达水平比率。
1.3.6 免疫荧光法检测NF-κB核转运变化[16]
将细胞悬浮液铺96孔细胞板,设立分组与重复,待细胞长至90%时弃培养液,根据碧云天NF-κB激活-核转运检测试剂盒进行操作,在荧光显微镜下观察NF-κB(p65)红色荧光和细胞核蓝色荧光亮度变化。
1.4 统计方法
数据以SPSS 18.0统计软件进行统计分析。所有变量以x±s表示。多组均数间采用单因素方差分析,两两之间比较采用Dunnett-t检验。
2 结果与分析
2.1 氯化锦葵色素-3-半乳糖苷对细胞表达ICAM-1蛋白的影响
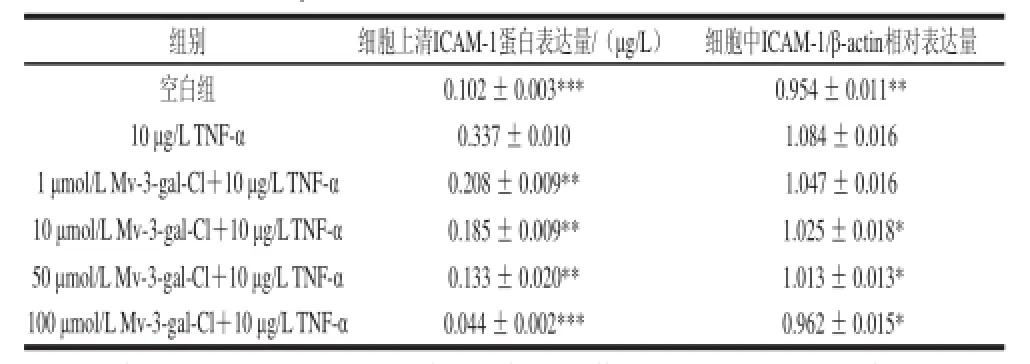
表1 不同浓度Mv-3-gal-Cl对TNF--α诱导的人脐静脉内皮细胞ICAM--1蛋白表达的影响(x±s,n=3)Table 1 Effect of Mv-3-gal-Cl on TNF-α-induced ICAM-1 protein expression in HUVECss (x±s, n=3)

图1 不同浓度Mv-3-gal-Cl对TNF--α诱导的人脐静脉内皮细胞中ICAM-1表达的影响Fig.1 Effect of Mv-3-gal-Cl at various concentrations on TNF-αinduced ICAM-1 protein expression in HUVECs
由表1和图1可知,通过ELISA及Western blotting方法检测不同浓度Mv-3-gal-Cl对细胞上清和细胞中ICAM-1蛋白水平的表达影响时发现,正常HUVEC细胞在未激活的状态下只有少量ICAM-1蛋白表达,在受到TNF-α诱导刺激下,ICAM-1表达显著增加。在细胞孵育不同浓度Mv-3-gal-Cl与TNF-α后,ICAM-1的表达明显下降,并呈浓度依赖性。浓度为1~50 μmol/L时,细胞上清中ICAM-1可溶性蛋白抑制效果明显(P<0.01);浓度为100 μmol/L时,抑制效果差异性很显著(P<0.001)。在浓度为10、50、100 μmol/L时,细胞中ICAM-1蛋白水平比较降低明显,抑制率分别为45.38%、54.61%、93.84%,具有显著性差异(P<0.05)。
2.2 氯化锦葵色素-3-半乳糖苷对细胞表达ICAM-1 mRNA的影响
为了检测Mv-3-gal-Cl如何在mRNA水平上抑制细胞表达ICAM-1,即采用实时荧光定量PCR检测细胞中ICAM-1 mRNA表达量的变化,用内参归一法确定相对表达量,由图2可知,单独TNF-α刺激细胞时ICAM-1 mRNA的表达量明显增加;同时孵育不同浓度的Mv-3-gal-Cl和TNF-α时,ICAM-1的相对mRNA的表达量随浓度的增加而降低。在浓度为1 μmol/L时,细胞中ICAM-1 mRNA表达量相对较低(P<0.05);在浓度为10、50、100 μmol/L时,细胞中ICAM-1 mRNA表达量相对单独TNF-α刺激组的抑制率为79.96%、87.94%、106.26%,具有明显显著性差异(P<0.01),从而对细胞具有一定的保护作用。

图2 不同浓度Mv-3-gal-Cl对TNF--α诱导的人脐静脉内皮细胞中ICAM-1 mRNA 表达的影响Fig.2 Effect of Mv-3-gal-Cl at various concentrations on TNF-αinduced ICAM-1 mRNA expression in HUVECs
2.3 氯化锦葵色素-3-半乳糖苷对细胞IκBα表达的影响

图3 不同浓度Mv-3-gal-Cl对TNF-α诱导的人脐静脉内皮细胞中IIκBα表达的影响Fig.3 Effect of Mv-3-gal-Cl at various concentrations on TNF-αinduced IκBα expression in HUVECs

图4 不同浓度Mv-3-gal-Cl对TNF-α诱导的人脐静脉内皮细胞中IIκBα表达的影响Fig.4 Effect of Mv-3-gal-Cl at various concentrations on TNF-αinduced IκBα expression in HUVECs
采用Western blotting检测细胞质中IκBα蛋白表达量的变化,由图3、4可知,正常细胞未受到刺激时IκBα未被酸化,含量很高。在受到TNF-α刺激时,细胞质中IκBα相对含量明显降低,IκBα被降解。当不同浓度的Mv-3-gal-Cl和TNF-α同时作用时,Mv-3-gal-Cl明显抑制TNF-α诱导的内皮细胞中IκBα的泛酸化降解,并且呈浓度依赖性。在浓度为1 μmol/L时,细胞中IκBα相对含量低(P<0.05);在浓度为10、50、100 μmol/L相比单独TNF-α刺激组具有显著性意义(P<0.01)。
2.4 氯化锦葵色素-3-半乳糖苷对细胞核NF-κB表达的影响
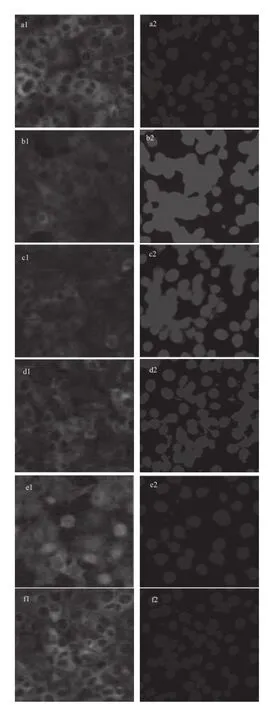
图5 不同浓度的Mv-3-gal-Cl对TNF-α诱导的人脐静脉内皮细胞中NNFF--κB核异位的变化Fig.5 Effect of Mv-3-gal-Cl at various concentration on TNF-αinduced NF-κB nuclear translocation in HUVECs
由图5可知,通过免疫荧光法检测NF-κB转入核内情况发现,细胞未受刺激时核内NF-κB的含量很低,荧光强度很弱。在单独受到TNF-α刺激时,NF-κB被激活,大量的转入细胞核内,捕捉的荧光强度变强。不同浓度Mv-3-gal-Cl和TNF-α同时作用下Mv-3-gal-Cl抑制了核转录因子NF-κB的激活,阻止其进入核内进而转录翻译各种相应蛋白,因而荧光强度渐渐变弱,抑制作用呈浓度依赖性。浓度为100 μmol/L时,荧光强度接近于正常组,具有显著意义。
3 结论与讨论
众所周知,细胞因子网络不仅参与维护正常血管的功能,而且参与涉及血管的各种病理过程。近年研究表明,心血管疾病的发生及发展是细胞通过分泌生长因子、趋化因子及其他细胞因子而相互调控的结果。ICAM-1在静息的白细胞、内皮细胞呈低水平表达,维持正常的细胞生理作用[17]。然而当细胞受到外界炎症因子TNF-α、IL-6、IFN-γ的刺激作用下,ICAM-1在纤维细胞、内皮细胞、淋巴细胞等表达迅速上调,影响细胞活力及正常功能[18-19]。大量的研究证实氧化应激亦可引起血管内皮功能障碍,而抗氧化物质对细胞具有保护作用,可抑制炎症的发生发展[20]。本研究从分子机理上研究抗氧化物蓝莓花青素氯化锦葵色素-3-半乳糖苷的抗炎症作用机理,结果显示氯化锦葵色素-3-半乳糖苷通过抑制NF-κB信号通路的激活减少TNF-α诱导的炎症因子ICAM-1蛋白和mRNA的过表达,并且抑制活性呈浓度依赖性。
TNF-α刺激内皮细胞表达ICAM-1蛋白主要是通过激活NF-κB信号传导途径实现[21];本研究通过Western blotting检测细胞质中IκBα蛋白得出,正常细胞中IκBα相对含量比较高,当受到TNF-α介导时,IκBα蛋白含量降低,验证了IκBα被磷酸化,随即被泛酸连接酶识别而被降解[22];而当孵育不同浓度氯化锦葵色素-3-半乳糖苷时,IκBα相对表达量相比刺激组呈浓度依赖性,具有显著性抑制作用。此外,通过免疫荧光检测NF-κB核转运情况也得到相应结果,单独TNF-α组捕捉的NF-κB荧光强度最强,NF-κB是与它的抑制性亚基IκB分开并进人核内激活靶基因转录[23];随着不同浓度的氯化锦葵色素-3-半乳糖苷的刺激,NF-κB荧光强度相对于对照组渐渐变弱,证实了氯化锦葵色素-3-半乳糖苷能阻止NF-κB的激活进入核内转录翻译。
总之,氯化锦葵色素-3-半乳糖苷作为花青素中的重要组成部分及营养物质,通过抑制NF-κB信号传导途径抑制黏附因子ICAM-1在蛋白及mRNA水平上的表达,从而对TNF-α诱导内皮细胞炎症具有一定抑制作用,防止内皮细胞损伤及紊乱,具有抗炎症作用。由于一般心血管疾病产生的原因中包括因为促炎症因子引起的内皮功能障碍及产生的炎症,氯化锦葵色素-3-半乳糖苷的抗炎症作用为防治心血管疾病提供了一定的理论基础。
[1] PRICE D T, LOSCALZO J. Cellular adhesion molecules and atherogenesis[J]. American Journal of the Medical Sciences, 1999, 107(1): 85-97.
[2] DUSTIN M L, ROTHLEIN R, BHAN A K, et al. Induction by IL-1 and interferon-gamma: tissue distribution, biochemistry, and function a natural adherence molecule (ICAM-1) [J]. Journal of Immunology, 1986, 137(1): 245-254.
[3] ZHOU Zhe, LIU Yong, MIAO A D, et al. Protocatechuic aldehyde suppresses TNF-α-induced ICAM-1 and VCAM-1 expression in human umbilical vein endothelial cells[J]. European Journal of Pharmacology, 2005, 513(1/2): 1-8.
[4] CHAI Hui, WANG Qiuyan, HUANG Lifeng, et al. Ginsenoside Rb1 inhibits tumor necrosis factor-alpha-induced vascular cell adhesion molecule-1 expression in human endothelial cells[J]. Biological Pharmaceutical Bulletin, 2008, 31(11): 2050-2056.
[5] AQIL F, GUPTA A, MUNAGALA R, et al. Antioxidant and antiproliferative activities of anthocyanin/ellagitannin-enriched extracts from Syzygium cumini L.(Jamun, the Indian blackberry) [J]. Nutrtion and Cancer: an International Journal, 2012, 64(3): 428-438.
[6] NIE, ZHAO Zhiping, CHEN Guoping, et al. Brassica napus possesses enhanced antioxidant capacity via heterologous expression of anthocyanin pathway gene transcription factors[J]. Journal of Plant Physiology, 2013, 60(1): 108-115.
[7] SMITH M A L, MARLEY K A, SEIGLER D, et al. Bioactive properties of wild blueberry fruits[J]. Journal of Food Science, 2000, 65(2): 352-356.
[8] PAIX‹O J, DINIS T C P, ALMEIDA L M. Malvidin-3-glucoside protects endothelial cells up-regulating endothelial NO synthase and inhibiting peroxynitrite-induced NF-κB activation[J]. Chemico-Biological Interactions, 2012, 199: 192-200.
[9] 沈阳辉. 白藜芦醇抗动脉粥样硬化作用分子机制的研究[D]. 福州:福建医科大学, 2007.
[10] HOSSEINIAN F S, BETA T. Saskatoon and wild blueberries have higher anthocyanin contents than other Manitoban berries [J]. Journal of Agricultural and Food Chemistry, 2007, 55(26): 10832-10838.
[11] FINKENZELLER G, GRANER S, KIRKPATRICK C J, et al. Impaired in vivo vasculogenic potential of endothelial progenitor cells in comparison to human umbilical vein endothelial cells in a spheroid-based implantation model[J]. Cell Proliferation, 2009, 42(4): 498-505.
[12] BELLA J, KOLATKAR P R, MARLOR C W, et al. The structure of the two amino-terminal domains of human ICAM-1 suggests how it functions as a rhinovirus receptor and as an LFA-1 integrin ligand[J]. Proceedings of the National Academy of Sciences, 1998, 95(8): 4140-4145.
[13] YUAN Mingjie, HUANG Congxing, TANG Yanhong, et al. A novel peptide ghrelin inhibits neural remodeling after myocardial infarction in rats[J]. European Journal of Pharmacology, 2009, 618(1/3): 52-57.
[14] ZAPATA E, VENTURA J K, RODRIGUEZ E, et al. Dehydroepiandrosterone inhibits the proliferation of HUVEC by enhancing the expression of p53 and p21, restricting the phosphorylation of RB, and is androgen- and estrogen-receptor independent[J]. Federation of European Biochemical Societies Journal, 2005, 272(6): 1343-1353.
[15] MIGEON C J, KELLER A R, LAWRENCE B, et al. Dehydroepiandrosterone and androsterone levels in human placenta. Effect of age and sex: dayto-day and diural variations[J]. Journal of Clinical Endocrinology & Metabolism, 1957, 17: 1051-1062.
[16] XU Zhiwei, LIN Shiqing, WU Weikang, et al. Ghrelin prevents doxorubicin-induced cardiotoxicity through TNF-alpha/NF-kappaB pathways and mitochondrial protective mechanisms[J]. Toxicology, 2008, 247(2/3): 133-138.
[17] HUANG W Y, CHAKRABART S, MAJUMDER K, et al. Eggderived peptide IRW inhibits TNF-α-induced inflammatory response and oxidative stress in endothelial cells[J]. Journal of Agricultural and Food Chemistry, 2010, 58(20): 10840-10846.
[18] HUNG C F, HUANG T F, CHEN B H, et al. Lycopene inhibits TNF-α-induced endothelial ICAM-1 expression and monocyte-endothelial adhension[J]. Journal of Pharmacology, 2008, 586(1/3): 275-282.
[19] 丁伟斌, 梁统, 周克元. 原花青素二聚体B2对大鼠滑膜细胞NF-κB核转运及炎症因子表达的影响[J]. 中国药理学通报, 2012, 28(6): 803-806.
[20] NOGUCHI N, HANYU R, NONAKA A, et al. Inhibition of THP-1 cell adhesion to endothelial cells by alphatocopherol and alphatocotrienol is dependent on intracellular concentration of the antioxidants[J]. Free Radical Biology and Medicine, 2003, 34(12): 1614-1620.
[21] ZHOU Z, CONNELL M C, MacEWAN D J. TNFR1-induced NF-κB, but not ERK, p38MAPK or JNK activation, mediates TNF-induced ICAM -1 and VCAM-1 expression on endothelial cells [J]. Cell Signal, 2007, 19(6): 1238-1248.
[22] HAYDEN M S, GHOSH S. Signaling to NF-kappaB[J]. Genes & Development, 2004, 18(18): 2195-2224.
[23] YAMOMOTO Y, GAYNOR R B. IκB kinses: key regulators of the NF-kappaB pathway[J]. Trends Biochemistry Science, 2004, 29(2): 72-79.
Inhibitory Effect of Malvidin-3-Galactoside Chloride on TNF-α-Induced Inflammation in Endothelial Cells
WANG Jian1,2, HUANG Wu-yang1,*, LI Chun-yang1, ZHENG Qi-sheng3, LIU Ya-mei3
(1. Institute of Farm Product Processing, Jiangsu Academy of Agricultural Science, Nanjing 210014, China; 2. College of Food Science and Technology, Nanjing Agricultural University, Nanjing 210095, China; 3. National Technical Research Centre of Veterinary Biological Products, Jiangsu Academy of Agricultural Science, Nanjing 210014, China)
Objective: The effect of malvidin-3-galactoside chloride (Mv-3-gal-Cl) on TNF-α-induced inflammation in human umbilical vein endothelial cells (HUVEC) was investigated in this study. Methods: HUVEC were treated with TNF-α (10 μg/L) alone and combined with Mv-3-gal-Cl (1, 10, 50 and 100 μmol/L), respectively. DMSO was used as control. The soluble ICAM-1 protein content in cell supernatant wa s detected by ELISA. The change in ICAM-1 mRNA expression level in cells was detected by RT-PCR. The expression of ICAM-1 and IкBα was assessed by Western blotting. The activity of NF-кB was evaluated by immunofluorescence. Results: Mv-3-gal-Cl appeared to specifically down-regulate the TNF-α-induced cell surface expression of ICAM-1 and ICAM-1 mRNA expression and protein release from HUVEC cells in a concentration-dependent manner. The inhibitory effect of Mv-3-gal-Cl on protein expression might be associated with the suppression of NF-κB in HUVECs. Conclusion: Mv-3-gal-Cl can inhibit TNF-α-induced inflammation in HUVECs. The anti-inflammatory effect of Mv-3-gal-Cl provides a theoretical basis for preventing and treating pro-inflammatory cytokinesinduced endothelial dysfunction and cardiovascular diseases resulting from inflammation.
malvidin-3-galactoside chloride (Mv-3-gal-Cl); human umbilical vein endothelial cell (HUVEC); tumor necrosis factor-α; inflammatory effect; intercellular cell adhesion molecule-1 (ICAM-1); NF-kappa B
R329.25;R332
A
1002-6630(2014)09-0241-05
10.7506/spkx1002-6630-201409047
2013-06-09
国家自然科学基金青年科学基金项目(NSFC31101264);江苏省自然科学基金项目(RK2011677);
江苏省农业自主创新项目(cx(12)5029)
王健(1988—),女,硕士研究生,研究方向为营养与功能食品。E-mail:wangjian454348952@126.com
*通信作者:黄午阳(1979—),女,副研究员,博士,研究方向为营养与功能食品。E-mail:wuyanghuang@hotmail.com
