三峡库区小江库湾鱼类食物网的稳定C、N同位素分析
2013-12-21徐丹丹王志坚岳兴建张耀光
李 斌,徐丹丹,王志坚,岳兴建,张耀光,*
(1.长江上游鱼类资源保护与利用四川省重点实验室,内江师范学院生命科学学院,内江 641112;2.淡水鱼类资源与生殖发育教育部重点实验室,西南大学,重庆 400715)
小江是长江北岸重要的一个一级支流,渔业资源丰富。三峡大坝蓄水后,三峡库区从以前的河流生态系统逐渐向水库生态系统演变,同期小江成为库区淹没面积最大、消落区域面积最广的一个一级支流,其生态环境的保护直接影响到三峡库区的生态状况。三峡大坝蓄水(145—175 m)后,小江形成了长约40 km的永久静水区域,被称为小江库湾(Xiaojiang Bay)。蓄水以来水文情势的改变、捕捞强度的增加、航运加剧、环境污染、水土流失等诸多因素的影响下,小江库湾鱼类生物多样性和资源量锐减[1]。近年来,小江的研究多集中在水体的富营养化、浮游植物及鱼类资源概况方面[1-4],对水生生态系统营养结构及功能方面的研究还未见报道。因此,为了了解蓄水对三峡库区支流水生生态系统的影响,选择以小江库湾作为一个研究区域进行鱼类食物网结构的研究有着十分重要的意义。由于水生生态系统食物网自身结构的复杂性[5-8]及研究方法的有限性,传统胃含物分析法很难准确解释清楚水生生态系统食物网结构及营养关系[9]。
近20年来,稳定碳(δ13C)、氮(δ15N)同位素技术作为胃含物分析方法的补充,在描述消费者与其食物之间的营养关系方面独树一帜。消费者的δ13C值与其食源相似,平均富集度约为0‰—1‰之间,是食物来源良好的示踪指标[10-11];δ15N在营养级的传递过程中富集度约为3.4‰,通常用来指示生物所处的营养级[12]。目前,稳定碳、氮同位素技术已经广泛应用于水域生态系统食物网结构及营养关系的研究[13-23]。本文以三峡库区支流小江库湾为研究对象,应用δ13C、δ15N技术探讨鱼类食物网的主要碳源及其营养路径,以期为小江库湾渔业资源恢复和管理提供理论支持。
1 材料与方法
1.1 研究区地点
小江流域(30°49'—31°42'N,107°56'—108°54'E)总面积5172 km2,干流全长182 km,是三峡库区中段北岸流域面积最大的支流。2010年10月三峡库区蓄水到175 m后,小江回水区延伸至开县县城(汉丰湖)境内,长约75 km。但考虑回水区末端受到175—145 m水位涨落的影响,本研究选择在145 m以下的永久回水区域,即小江库湾(从双江口镇到渠口镇下游,长约40 km(图1)。为全面反映小江库湾鱼类食物网结构的情况,并综合三峡库区水位调节变化规律(175—145 m)的影响,该研究分别于枯水期(7月份)和丰水期(12月份)进行了样品采集工作(受三峡库区水位调节的影响每年的5—8月份为枯水期,9月至翌年4月为丰水期):POM(颗粒有机物)和固着藻类设置了3个采样点(图1);水生植物(仅在枯水期采集到样品)、软体动物和消落带陆生植物沿小江库湾随机采集其优势种类;鱼类样品主要在渠口到双江镇水域采集。

图1 研究地点位置和采样站点Fig.1 Location of the Xiaojaing Bay and sampling sities
1.2 样品采集及处理
徒手采集优势水生植物(水蓼Polygon hydropiper和喜旱莲子草Alternanthera philoxeroides)和沿岸优势陆生植物(苍耳Xanthium sibiricum、狗尾草Setaria faberii和狗牙根Cynodon dactylon)用清水除去表面附着物。POM(水中颗粒有机物,代表浮游植物)采集方法如下:取上层水,先通过浮游动物网(网目112 um)过滤后水样经预烧的玻璃纤维滤膜(GF/C Whatman)过滤获得含有颗粒有机物样品滤膜用锡箔纸包裹,存放于封口袋中。固着藻类从沿岸的石头、船体上刮取,挑出易区分的杂质,过滤到滤膜上(GF/C Whatman),处理方法同POM。每一样点POM和固着藻类样品至少有3个重复样混合而成。徒手采集软体动物铜锈环棱螺(Bellamya aeruginosa)和大瓶螺(Pomacea canaliculata),并将其放置于充分曝气的蒸馏水中过夜,使其肠含物排空。每个采样点铜锈环棱螺样品至少由30个个体混合而成,去除其外壳获取软组织。鱼类样品通过聘请渔民电捕获得,取其背侧肌肉。所有样品通过移动冰箱运回到实验室于-20℃低温保存备用。用于稳定碳同位素分析的滤膜先用1 mol/L的盐酸处理,然后用蒸馏水冲洗干净。所有样品在60℃烘干至恒重,用研钵研磨成均一粉末,放入干燥器保存待测。
1.3 稳定性同位素测试
所有样品的稳定C和N同位素比值均在西南大学地理科学学院地球化学与同位素实验室测定,稳定同位素质谱仪为菲尼根Flash EA112元素分析仪与菲尼根DELT plus XP稳定同位素质谱仪相连而成。稳定C、N同位素的自然丰度表示为:

式中,X是13C或15N,R是13C/12C或15N/14N;δ13C值是相对于国际PDB标准;δ15N值是相对于空气氮气的丰度。分析精度 δ13C <0.20‰,δ15N<0.30‰。
营养级计算公式:

式中,Δδ15N=3.4‰;当 baseline 是生产者时,λ=1;当 baseline初级消费者是,λ=2。
1.4 统计分析
应用SPSS 17.0软件对小江库湾枯水期和丰水期间共有的生物类群的δ13C、δ15N值进行了方差分析(ANOVA);以不同时期鱼类δ13C、δ15N值作为双重因对鱼类食性类型进行了聚类分析。
2 结果
2.1 稳定碳、氮同位素比值
表1显示了小江库湾枯水期和丰水期间初级生产者和消费者稳定碳、氮同位素比值。初级生产者中消涨区陆生C4植物狗牙根和狗尾草拥有最高的 δ13C值,分别为-12.62‰、-11.54‰;水生维管束植物 δ13C值(-29.60‰—-29.62‰)最低;固着藻类和 POM 样品 δ13C(平均值分别为(-24.85±0.38)‰、(-22.19±1.33)‰)值差异显著(P<0.05),且枯水期和丰水期间没有显著性差异(表2,P>0.05)。初级生产者固着藻类和POM样品的δ15N值变化幅度大于其δ13C值,为3.99‰—6.25‰,但不同时期差异并不显著(P>0.05);消费者大眼鳜Siniperca kneri拥有最高的δ15N值(13.65‰),其次是蒙古鲌Erythroculter mongolicus和翘嘴鲌Erythroculter ilishaeformis。甲壳类秀丽白虾Leander modestus和日本沼虾Macrobrachium mipponensisδ15N值较高,分别为 12.09‰—13.50‰,(12.16±0.44)‰;除了鲤 Cyprinus carpio、似鳊 Pseudobrama simoni和瓦氏黄颡鱼Pelteobagrus vachelli的δ15N值在枯水期和丰水期具有显著性差异外(表2,P<0.05),其它类群均无显著性差异(表 2,P > 0.05)
2.2 食物网结构及营养路径
基于δ15N 值在营养级传递过程中相对稳定的富集规律(3.4‰)计算了小江库湾鱼类的营养级(表1)。不同时期(枯水期和丰水期)小江库湾食物网均有3个营养级,其中POM、固着藻类及水生植物(初级生产
者)位于食物网的底层,鱼类位于食物网的上层,而无脊椎动物(背角无齿蚌Anodonta woodiana和螺类)介于两者之间(图2和图3)。在小江库湾鱼类食物网中蒙古鲌、翘嘴鮊及大眼鳜等位于第三营养级,其中以大眼鳜营养级最高(3.43—3.55),多数鱼类介于第二和第三营养级之间。小江库湾不同时期鱼类营养功能类群的聚类分析图显示(图4和图5)枯水期与丰水期小江库湾鱼类食性类型主要可分为3种,分别为杂食偏植食性类群(Ⅰ)、杂食偏动物食性类群(Ⅱ)和动物食性类群(Ⅲ)(图4和图5)。根据稳定C、N同位素数据和已有鱼类的胃含物分析结果[24]推测可知,小江库湾鱼类食物网主要通过4种营养途径经过2—3个环节进行碳源的传递。第一种营养路径,以POM为碳源经过浮游动物(未采集)到摄食浮游动物鱼类(接近第三营养级)如鳙Aristichthys nobilis、鲢Hypophthalmichthys molitrix,最终到顶端捕食者(接近第四营养级)。第二种和第三种营养路径,以底栖固着藻类为碳源,经过无脊椎动物(秀丽白虾、日本沼虾等)到浮游食物链中偏动物食性鱼类(第三营养级)如蒙古鮊、翘嘴鮊;或经过无脊椎动物(螺类)到底栖杂食偏动物性鱼类(接近第三营养级),最终传递到高级捕食者(接近第四营养级),如大眼鳜。第四种营养路径,以外源营养物质输入为碳源,通过以腐殖质为营养来源的水生昆虫(未采集)到底栖偏动物食性鱼类,最终进入顶端捕食者。

表1 小江库湾初级生产者和消费者稳定C、N同位素比值(平均数±标准误)Table 1 The δ13 C and δ15 N values(mean±SE)of the primary producers and consumers in the Xiaojiang Bay
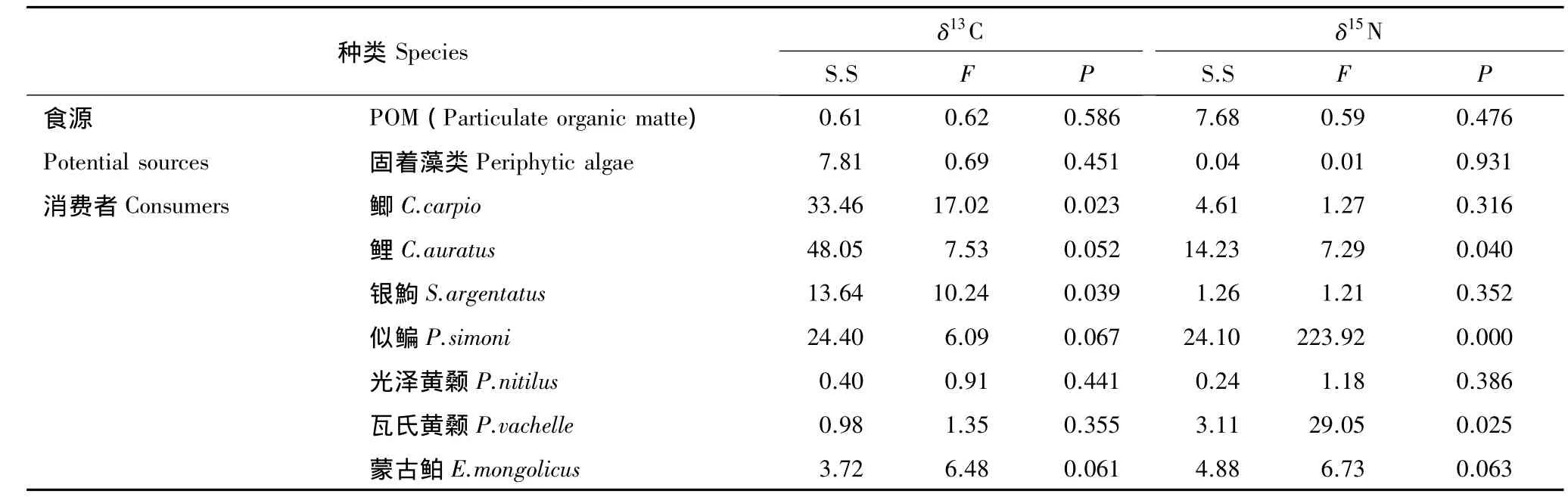
表2 枯水期与丰水期不同类群生物之间同位素比值组成的方差分析Table 2 The different organisms Variance analysis of between in dry season and wet season
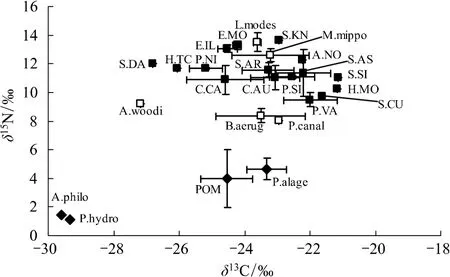
图2 枯水期小江库湾食物网结构模式图(2010-07)Fig.2 The food web structure's model of the Xiaojiang Bay in the dry season

图3 丰水期小江库湾食物网结构模式图(2010-12)Fig.3 The food web structure's model of the Xiaojiang Bay in the wet season
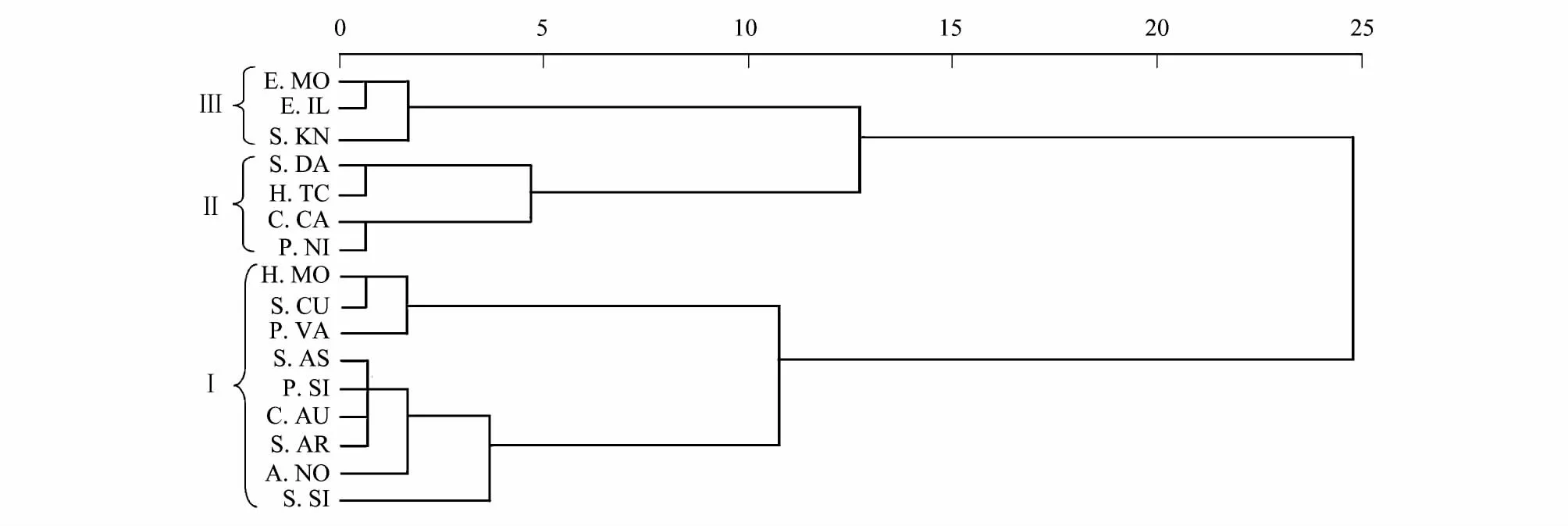
图4 枯水期间基于小江库湾16种鱼类δ13C、δ15 N值的聚类分析Fig.4 Cluster analysis ofδ13C and δ15N values for 16 fish species in dry season from the Xiaojiang Bay
3 讨论
3.1 基线值的确定

图5 枯水期间基于小江库湾13种鱼类δ13C、δ15 N值的聚类分析Fig.5 Cluster analysis of δ13C and δ15N values for 16 fish species in wet season from the Xiaojiang Bay
不同水域食物网中 δ15N 基线值差异较大,确定消费者营养级关键在于寻找合适 δ15N基线值[25]。David[26]指出生活周期较长,个体较大的螺类和双壳类δ15N值能有效的反映研究区域中初级生产者、碎屑及有机物等同位素特征。因此,它们的δ15N值作为基线值已经广泛用于评价次级消费者和高营养级生物的营养级位置[27]。该研究未能在丰水期采集到软体动物螺类或双壳类,但已有研究表明它们δ15N值组成不易受季节变化的影响[26]。因此,采用枯水期间铜锈环棱螺和背角无齿蚌δ15N值的平均值作为基线值分别计算小江库湾枯水期和丰水期间鱼类营养级。
3.2 小江库湾鱼类能量源
固着藻类、POM、水生植物和消落带陆生植物等构成了小江库湾食物网中消费者在不同时期(枯水期和丰水期)的潜在食源,其δ13C、δ15N值(陆生植物未测定δ15N值)差异显著。枯水期,绝大多数消费者δ13C值落在POM和固着藻类的δ13C值范围中,表明了POM和固着藻类是小江库湾消费者(鱼类)重要食物来源;与POM、固着藻类不同,水生植物具有特别贫乏的δ13C值,进一步证明颗粒有机物(POM)和固着藻类在支持小江库湾枯水期间食物网碳源驱动中的重要性。与枯水期相比,丰水期经历了三峡库区的蓄水过程(小江库湾水位从145 m左右上升到175 m),蓄水致使两岸陆生C4/C3植物、农田有机物及陆生昆虫等大量的陆生营养物质进入水体,为消费者提供了多元化的潜在食源[28-29],致使似鳊(P.SI)、铜鱼Coreius heterodon(C.HE)、鲫(C.AU)和鲤(C.CA)等消费者有机会摄食了δ13C值更为贫乏或更为富集的食源,同时暗示了陆生碳源输入对消费者能量贡献的重要性。
在鱼类群落中,摄食策略是影响稳定同位素数据分析鱼类复杂食物网营养关系的重要因素[30]。小江库湾银鮈、蛇鮈和瓦氏黄顙鱼等多种鱼类的δ13C值较为相近,暗示了它们的食物来源重叠度高;鲫、鲤不同时期(枯水期和丰水期)δ13C值差异显著,可能与它们摄食策略的改变相关(摄食了不同的食源)[31-33]。聚类分析显示了丰水期间似鳊(P.SI)单独聚类为一支,但胃含物分析显示其食物主要来源于藻类、植物碎屑等,综合考虑其食性类型为杂食偏植物食性更为准确。已有肠含物分析表明瓦氏黄顙鱼Pelteobagrus vachelli(P.VA)为杂食偏植物食性鱼类[34],但聚类分析显示(图5)瓦氏黄顙鱼(P.VA)和大眼鳜(S.KN)、蒙古鲌(E.MO)等动物食性鱼类聚为一支。研究结果建议,稳定碳、氮同位素在分析鱼类食物来源及营养关系时需结合肠含物分析结果才能更准确的理解同位素数值的意义。
3.3 食物网结构和营养级
在不同时期小江库湾大眼鳜(动物食性鱼类)均位于食物网的顶端。杂食性鱼类的营养级在不同时期变化不大,与Chassot等[35]得出杂食性鱼类的营养级不容易受到时间变化的影响这一结论一致。通常,湖泊生态系统中平均食物链长度为4.0个营养级[36]。与湖泊生态系统不同,小江库湾属于河流型水库生态系统,在枯水期和丰水期间均有3.0个营养级;但与枯水期相比,丰水期小江库湾食物网营养级长度略有下降,可能与蓄水过程中鱼类潜在食源(陆生输入)的种类增多有关,因为陆生营养物质拥有相对低的15N[37]。小江库湾食物链长度的相对稳定性可能暗示了该水域食物网结构的稳定性。Persson等[34]指出食物网营养级长度与生境中生产率或生境的复杂性有关;另有观点表明食物链的长度与生态系统大小呈正相关关系[39],但生态系统大小对食物链长度影响有多大,目前仍不清楚[39]。
总体而言,在三峡工程运行初期小江库湾已经形成了相对稳定的食物网结构,但随着三峡库区运行时间的增加,小江库湾鱼类食物网结构和营养关系将如何变化?仍需长期的监测。致谢:感谢西南大学彭作刚教授在论文写作过程中给予的帮助。
[1]Li B,Jiang X,Wang Z J,Jin L,Duan X B,Zhang Y G.The current situation of fishery resources in the Xiaojiang River of the Three Gorges Reservoir.Freshwater Fisheries(Chinese),2011,41(6):37-42.
[2]Li Z,Fang F,Guo J S,Chen J,Zhang C,Tian G.Spring algal bloom and nutrients characteristics in Xiaojiang River backwater area,Three Gorges Reservoir,2007.Journal of Lake Sciences,2009,21(1):36-44.
[3]Guo JS,Chen J,Li Z,Fang F,Zhang C,Tian G.Investigation of phytoplankton and assessment of Algal diversity on Backwater Area of Xiaojiang River in Three Gorges Reservoir after its initiate impounding to the water level of 156 m in Spring.Chinese Journal of Environmental Science,2008,29(10):2710-2715.
[4]Pan X J,Peng J H,Zhang Z Y,Zheng Z W.The annual characteristics of phytoplankton Communities in Pengxi River.Environmental Science&Technology,2009,32(Special):95-101.
[5]Post D M.Using stable isotopes to estimate trophic position:models,methods,and assumptions.Ecology,2002,83(3):703-718.
[6]Gray E V,Boltz JM,Kellogg K A,Stauffer JR.Food resource partitioning by nine sympatric darter species.Transactions of the American Fisheries Society,1997,126(5):822-840.
[7]Scott S,Pardo R,Vila I.Trophic niche overlap between two Chilean endemic species of Trichomycterus(Teleostei:Siluriformes).Revista Chilena De Historia Natural,2007,80(4):431-437.
[8]Norkko A,Thrush SF,Cummings V J,Gibbs M M,Andrew N L,Norkko J M,Schwarz A M.trophic structure of coastal Antarctic food webs associated with changes in sea ice and food supply.Ecology,2007,88(11):2810-2820.
[9]Jones JI,Waldron S.Combined stable isotope and gut contents analysis of food webs in plant-dominated,shallow lakes.Freshwater Biology,2003,48(8):1396-1407.
[10]Gu B,Schelske CL,Hoyer M V.Stable isotopes of carbon and nitrogen as indicators of diet and trophic structure of the fish community in a shallow hypereutrophic lake.Journal of Fish Biolgoy,1996,49(6):1233-1243.
[11]Persic A,Roche H,Ramade F.Stable carbon and nitrogen isotope quantitative structural assessment of dominant species from the Vaccares lagoon trophic web(Cammargu Biosphere Reserve,France).Estuarine,Coastal and Shelf Science,2004,60(2):261-272.
[12]DeNiro M J,Epstein S.Influence of diet on the distribution of nitrogen in animals.Geochimica et Cosmochimica Acta,1978,42(5):341-355.
[13]Hasson S,Hobbie JE,Elmgren R,Larsson U,Fry B,Johansson S.The stable nitrogen isotope ratio as a marker of foodweb interactions and fish migration.Ecology,1997,78(7):2249-2257.
[14]Kohzu A,Tayasu I,Yoshimizu C,Maruyama A,Kohmatsu Y,Hyodo F,Onoda Y,Igetaa A,Matsui K,Nakano T.Nitrogen-stable isotopic signatures of basal food items,primary consumers and omnivores in rivers with different levels of human impact.Ecological Research,2009,24(1):127-136.
[15]Karube Z,Okada N,Tayasu I.Sulfur stable isotope signature identifies the source of reduced sulfur in benthic communities in macrophyte zones of Lake Biwa,Japan,Limnology,2012,13(3):269-280.
[16]Hoekman D,Bartrons M,Gratton C.Ecosystem linkages revealed by experimental lake-derived isotope signal in heathland food webs.Oecologia,2012,170(3):735-745.
[17]Sun Z G,Mou X J,Li X H,Wang L L,Song H L,Jiang H H.Application of stable isotope techniques in studies of carbon and nitrogen biogeochemical cycles of ecosystem.Chinese Geographical Science,2011,21(2):129-148.
[18]Wang Y Y,Yu X B,Zhang L,Xu J.Food web structure of Poyang Lakeduring the dry season by stable carbon and nitrogen isotopes analysis.Acta Ecology Sinica,2009,29(3):1181-1188.
[19]Li B,Wang Z J,Jin L,Jiang X,Xu D D,Liu X L,Chen D Q,Zhang Y G.Effects of anthropogenic nutrient input on organisms from different trophic levels in Hanfeng Lake:Evidence from stable carbon and nitrogen isotope analysis.Acta Ecology Sinica,2012,32(5):1519-1526.
[20]Mao Z G,Gu X H,Zeng Q F,Zhou L H,Sun M B.Food web structure of a shallow eutrophic lake(Lake Taihu,Chine)assessed by stable isotope analysis.Hydrobiologia,2012,683(1):173-183.
[21]Quan W M.Food Web Analysis of Salt Marshes of the Yangtze River Estuary by Using Stable Isotopes.Fudan University Shanghai,2007.
[22]Wan Y,Hu JY,An L H,An W,Yang M.Determination of trophic relationships within a Bohai Bay food web using stableδ13Candδ15N analysis.Chinese Science Bulletin,2005,35(2):123-130.
[23]Cai D L,Hong X G,Mao X H,Zhang SF,Han Y B,Gao SL.Preliminary studies on trophic structure of tidal zone in the Laoshan Bay by using carbon stable isotopes.Axta Oceanologica Sinica,2001,23(4):41-47.
[24]Zhang L.A stable carbon and nitrogen isotope study of fishes from the Three Gorges riverine section fo the Yangtze River.Institute of Hydrobiology,Chinese Academy of Sciences Wuhan,2007.
[25]Vander Zanden M J,Rasmussen J B.Primary consumer δ13C and δ15N and the rophic position of aquatic consumers.Ecology,1999,80(4):1395-1404.
[26]David M,Post.Using stable isotopes to estimate trophic position:models,methods,and assumptions[J].Ecology,2002,83(3):703-718.
[27]Vander Zanden M J,Chandra S,Allen B C,Reuter J E,Goldman C R.Historical food web structure and the restoration of native aquatic communities in the Lake Tahoe(California-Nevada)basin.Ecosystems,2003,6(3):274-288.
[28]Smith B N,Epstein S.Two categories of13C/12C ratios for higher plants.Plant Physiology.1971,47(3):380-384.
[29]Kendall C,Silva SR,Kelly V J.Carbon and nitrogen isotopic compositions of particulate organic matter in four large river systems across the United States.Hydrological Processes.2001,15(7):1301-1346.
[30]Davenport SR,Bax N J.A trophic study of a marine ecosystem of Southeastern Australia using stable isotopes of carbon and nitrogen.Canadian Journal of Fisheries and Aquatic Sciences,2002,59(3):514-530.
[31]Hansona C E,Hyndesa G A,Wang SF.Differentiation of benthic marine primary producers using stable isotopes and fatty acids:implications to food webs studies.Aquatic Botany,2010,93(2):114-122.
[32]Coat S,Monti D,Bouchon C,Lepoint G.Trophic relationships in a tropical stream food web assessed by stable isotope analysis.Freshwater Biology,2009,54(5):1028-1041.
[33]Eisner L,Hillgruber N,Martinson E,Maselko J.Pelagic fish and zooplankton species assemblages in relation to water mass characteristics in the northern Bering and southeast Chukchi seas.Polar Biology,2013,36(1):87-113.
[34]Chassot E,Rouyer T,Trenkel V M,Gascuel D.Investigating trophic-levle variability in Celtic Sea fish predators.Journal of Fish Biology,2008,73(4):763-781.
[35]Vander Zanden M J,Fetzer W W.Global patterns of aquatic food chain length.Oikos,2007,116(8):1378-1388.
[36]Wangtzen K M,Machado F A,Voss M.Seasonal isotopic shifts in fish of the Pantanal wetland,Brazil.Aquatic Sciences,2002,64(3):239-251.
[37]Persson L,Didhl S,Johansson L,Andersson G,Hamrin S F.Trophic interactions in temperate lake ecosystems-a test of food-chain theory.The American Naturalist,1992,140(1):59-68.
[38]Vander Zanden M J,Shuter B J,Lester N,Rasmussen JB.Patterns of food chain length in lakes:a stable isotope study.The American Naturalist,1999,154(4):406-416.
[39]Post D M.The long and short of food-chain length.Trends in Ecology & Evolution,2002,17(6):269-277.
参考文献:
[1]李斌,江星,王志坚,金丽,段辛斌,张耀光.三峡库区蓄水后小江鱼类资源现状.淡水渔业,2011,41(6):37-42.
[2]李哲,方芳,郭劲松,陈杰,张超,田光.三峡小江回水区段2007年春季水华与营养盐特征.湖泊科学,2009,21(1):36-44.
[3]郭劲松,陈杰,李哲,方芳,张超,田光.156m蓄水后三峡水库小江回水区春季浮游植物调查及多样性评价.环境科学,2008,29(10):2710-2715.
[4]潘晓洁,彭建华,张志永,郑志伟.澎溪河浮游植物群落结构的周年变化特征.环境科学与技术,2009,21(1):36-44.
[18]王玉玉,于秀波,张亮,徐军.应用碳、氮稳定同位素研究鄱阳湖枯水末期水生食物网结构.生态学报,2009,29(3):1181-1188.
[19]李斌,王志坚,金丽,江星,徐丹丹,刘晓蕾,陈大庆,张耀光.人为营养物质输入对汉丰湖不同营养级生物的影响——稳定C、N同位素分析.生态学报,2012,32(5):1519-1526.
[18]全为民.长江口盐沼湿地食物网的初步研究:稳定同位素分析.上海:复旦大学,2007.
[30]张亮.长江三峡江段鱼类碳、氮稳定性同位素研究.武汉:中国科学院水生生物研究所,2007.
