传统海产调味品中微生物及其发酵作用研究进展*
2013-11-21李莹白凤翎励建荣
李莹,白凤翎,励建荣
(渤海大学化学化工与食品安全学院,辽宁省食品安全重点实验室,“食品贮藏加工及质量安全控制工程技术研究中心”辽宁省高校重大科技平台,辽宁锦州,121013)
传统发酵海产调味品通常以新鲜的鱼或虾为原料,添加高浓度的盐(一般为20% ~30%),经过1~2年的长时间发酵,利用原料自身蛋白酶、细菌蛋白酶及各种酶类分解蛋白质、脂类、碳水化合物大分子物质形成氨基酸、脂肪酸及各种风味物质,赋予产品鲜、香、咸等品质特征。此外,产品中富含有易于人体吸收的氨基酸、多肽、维生素以及钙磷铁等矿物质,具有提高免疫力,防贫血,降血压血脂等功效。
世界很多国家生产各种传统发酵海产调味品,如中国的鱼露、虾油虾酱,韩国的 Jeotkal、Sikhae,日本的Nakazuke,印度尼西亚的 Bakasang,加纳的Momoni,埃及的 Feseekh,伊朗的 Mahyaveh,泰国的 Plaasom以及Kungchao等等。在东南亚国家,传统发酵海产调味品是当地居民饮食中十分重要的佐餐食品,可作为烹饪的调味剂和菜肴的增香剂,有时甚至被作为下饭的一道小菜[1]。不同国家海产调味品在生产方法、原料配方和消费者的喜好等方面都存在一定的差异[2]。本文从发酵微生物对产品品质作用与影响的角度出发,对各种传统发酵海产调味品中微生物的构成、微生物在发酵过程中的功能特性的研究进展进行综述,旨在利用微生物的发酵作用改善产品风味,降低产品盐度,缩短发酵周期,提高经济效益。
1 传统发酵海产调味品中微生物菌相构成
不同发酵海产品在发酵过程中微生物的种类不同,同一产品在不同发酵时期微生物的种类也不同。Sanchez和Klitsaneephaiboon对传统发酵鱼酱油Patis发酵相关的微生物进行研究,结果表明发酵初期的优势菌是芽孢杆菌属的短小芽孢杆菌、凝结芽孢杆菌和枯草芽孢杆菌,发酵中期优势菌为地衣芽孢杆菌、生皱微球菌和表皮葡萄球菌,发酵终期优势菌为变异微球菌和腐生葡萄球菌[6]。
2 传统海产调味品中微生物的发酵作用
针对传统发酵海产调味品中微生物的研究主要集中在特征性嗜盐菌的筛选和分类鉴定方面,而对嗜盐菌的发酵作用研究相对偏少,研究主要包括耐盐蛋白酶的功能、香气和风味物质形成和生物胺降解等方面。
2.1 微生物源耐盐蛋白酶
传统发酵海产调味品的生物降解过程主要是将不溶的蛋白质转化为可溶的或悬浮的多肽和氨基酸,所依赖的是海产自身的和来自嗜盐菌的蛋白酶。在发酵中后期,嗜盐菌的耐盐蛋白酶的作用更为突出。
现今商用微生物源蛋白酶大多来自嗜温型微生物,一般在中性至弱碱性pH值范围、常温和低离子强度条件下显示高蛋白酶活性,而在极端条件下活性较差。如地衣芽孢杆菌蛋白酶在pH 4.0时活性较低,随NaCl浓度的增加活性逐渐降低[7]。相反,极端环境微生物产生的蛋白酶却有不同的表现,一些古细菌蛋白酶在5.8% ~30%NaCl条件下均具有活性,如亚洲嗜盐碱杆菌172P1产生的糜蛋白酶在30%NaCl下具有最佳活性[8]。
传统发酵海产调味品中的极端嗜盐菌具有形成蛋白酶的能力,如来自泰国鱼酱的4株嗜盐四联球菌M11、MS33、MRC5-5-2和 MRC10-7-8在高盐浓度下对鱼肉蛋白都具有分解活性,所产生的寡肽和低聚肽是形成产品风味物质的前体[3]。
嗜盐菌蛋白酶在较低盐度条件下活性很低,在一定范围内酶活性随着盐浓度的增高逐渐增强。来自泰国凤尾鱼鱼酱油的嗜盐菌菌株Virgibacillus sp.SK33产生耐盐丝氨酸蛋白酶,其活性在低盐度条件下较弱,随盐浓度的增加活性逐渐增强,在25%NaCl时活性最高。相比而言,这种天然蛋白酶比碱性蛋白酶和复合蛋白酶具有更好的水解活性[9]。
2.2 微生物源性香味和风味物质
微生物发酵作用对传统发酵海产调味品特征品质形成具有非常重要的呈献作用,一是依靠酶解产物或产物之间相互作用形成的醛、酮、醇、酯类等香气成分,二是来自酶解作用形成的氨基酸、寡肽类等风味物质[10]。Jirawat等将 T.halophilus MS33 和 MRC5-5-2接种至25%NaCl的鱼肉汤中,检测出形成了1-丙醇、2-甲基丙醛和苯甲醛等挥发性物质,与自然发酵鱼酱中的主要挥发性物质组成基本一致[3]。由此说明传统发酵海产调味品中的微生物对风味和香气物质的形成起着十分重要的作用。
2.2.1 香味物质
两组均行鼻内镜手术治疗。常规局麻,观察组2例,对照组1例因病变范围广泛行低血压控制性全麻。采用Messerk-linger术式,切除鼻腔内钩突及息肉,清除不可逆病变组织,开放病变鼻窦;术后采用藻酸钙敷料填塞止血,48h后取出。
传统发酵海产调味品有其特有的香味,如氨味、芝士味、肉香味和奶油香味。氨味挥发性物质包括氨、胺和含氮化合物,它们对产品的香味有增强作用。芝士味主要是由酮类物质形成的,而醛类物质呈现肉香味,小分子量的脂肪酸、乙酸和n-丁酸等呈现奶油香味。
大量研究发现,海产调味品中微生物参与香味物质的形成。当腌鱼调料含有抗生素或防腐剂时,产品就不具有典型的芳香气味,同时挥发性脂肪酸(VFA)的含量很低。VFA不是脂肪酸自然氧化的产物,而是来自于嗜盐菌的次级代谢产物。嗜盐菌代谢形成的氨基酸、寡肽进一步分解产生的醇类、酮类、醛类以及酸类成分是香味挥发性物质的来源。例如亮氨酸分解形成的代谢产物2-甲基丁醛、3-甲基丁醛和3-甲基丁醇主要是芝士风味及肉香味的挥发性成分[11]。
Cha等从传统发酵鱼酱、虾酱中检测出多达155种挥发性物质,其中占绝大多数为醛、酮、醇、酯类、芳烃、含氮和含硫化合物等挥发性物质,一些特征性含氮杂环化合物如吡嗪主要存在虾酱中[12]。泰国鱼酱中的嗜盐乳酸菌有助于醇类的积累,同时也产生少量的乙酸乙酯和不同含量的丙酮、2-丁酮、2,3-丁二酮和环己酮等挥发性成分,其中,菌株 T.halophilus MS33、MRC5-5-2、MCD10-5-15 有利于 2-甲基丙醛的形成,菌株MRC10-1-3有利于苯丙醛形成[3]。
发酵海产调味品中微生物发酵过程中在形成各种香气物质的同时,也产生不受人们欢迎的二甲基硫醚、二甲基二硫醚、二甲基三硫醚等臭味物质。一些细菌具有抑制这些物质形成的能力,Fukami等人的研究表明,在鱼酱中添加木糖葡萄球菌(Staphylococcus xylosus)后,终产品中的二甲基二硫醚、二甲基三硫醚含量显著减少[13]。
2.2.2 风味物质
传统发酵海产调味品最主要的风味是由挥发性化合物产生的咸味,其次是由氨基酸、寡肽和多肽呈现的特殊风味。海产调味品的咸味主要是由氯化钠和一些其他无机盐以及谷氨酸单钠盐、天门冬氨酸单钠盐产生的。例如,蚝油中呈鲜味成分有琥珀酸钠、谷氨酸钠等,呈甜味成分包括甘氨酸、丙氨酸、脯氨酸等[14]。Saisithi等发现在泰国腌鱼调料Nam-pla中的呈鲜物质主要含有组氨酸、酪氨酸和色氨酸等[15]。泰国鱼酱中的T.halophilus发酵鱼肉蛋白产生的氨基酸主要是组氨酸,也形成微量的酪氨酸[3]。
不同海产调味品中微生物的发酵作用与风味形成具有密切的联系,Beddows等对马来西亚Budu的研究结果也表明微生物对风味物质以及VFA的形成作用很大[16]。
2.3 微生物降解生物胺作用
传统海产调味品发酵过程会产生生物胺,过量生物胺会使发酵食品产生严重的食品安全问题[17-20]。传统发酵海产调味品中生物胺的主要成分组胺是由多种微生物的组氨酸脱羧酶诱导产生的,鱼酱油中含有超过1 000 mg/L 的组胺[21-24]。在发酵过程中,可以利用微生物控制组胺的形成,提高产品安全性[25]。研究表明,一些嗜盐菌如枝芽孢杆菌属、极端嗜盐古细菌等具有降解生物胺形成的作用。一般是通过自身组胺氧化酶氧化降解组胺为氨、醛以及过氧化氢的途径,减少在食品内的累积[26]。鱼酱油中的嗜盐微生物Brevibacillus sp.SK35可以使组胺降解率达99%[27]。Zaman 等的研究表明,Staphylococcus carnosus FS19和Bacillus amyloliquefaciens FS05分别使鱼酱油中组胺含量降低了27.7%和15.4%[28]。Dapkevicius等从鱼露中分离的78株奥默柯达酵母菌(Kodamaea ohmeri)中有胺氧化酶活性的有48株,能降解鱼酱中50%的组胺的菌有2株[29]。
3 传统海产调味品中微生物的发酵作用研究状况
世界范围内,对传统发酵海产品中微生物学研究主要包括两个方面,一是对发酵过程和产品中微生物形态学和分类学研究,二是对微生物和产品品质之间的相互作用进行研究,目前已从最初的嗜盐菌的分离鉴定及分类学研究逐步向产品的发酵作用与功能方面深入。
表1是近些年传统海产调味品中微生物的发酵作用及功能研究状况,从中可以看出微生物的作用和功能主要包括四个方面:一是研究微生物的蛋白酶作用与功能以及对产品品质的影响;二是研究微生物形成产品的风味物质作用和能力;三是研究微生物降解组胺的作用;四是以微生物作为发酵剂提高产品品质和功能方面的研究。
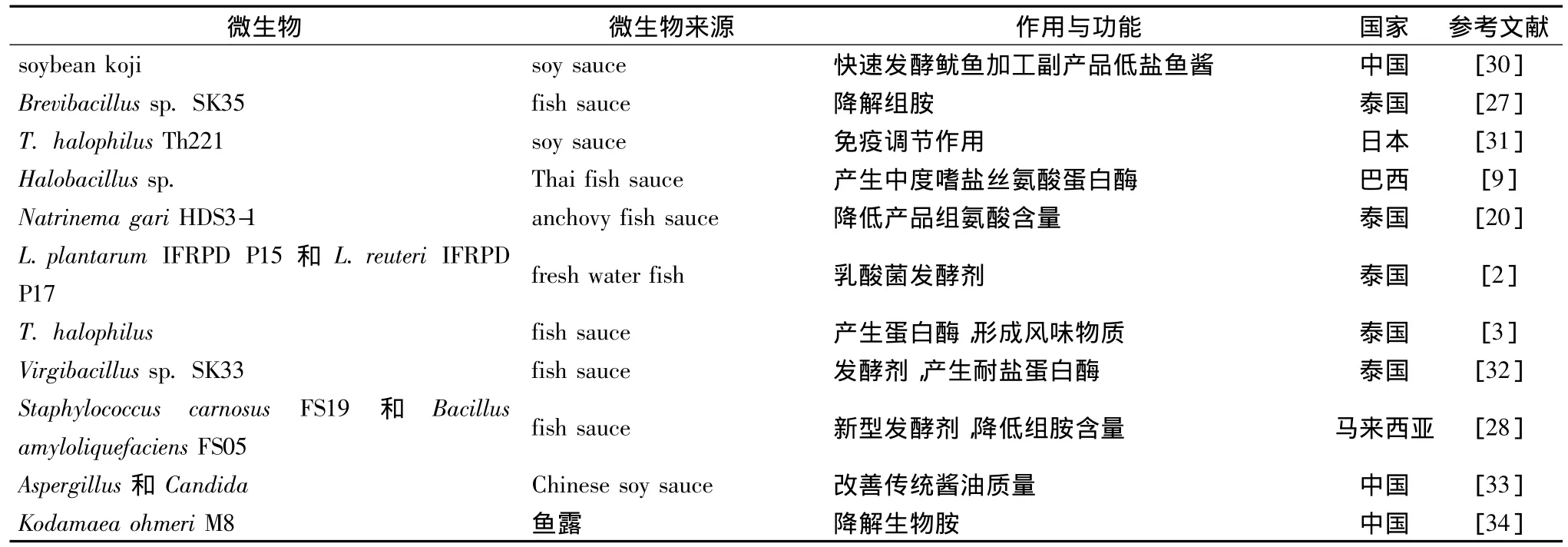
表1 传统发酵海产调味品中微生物的发酵作用及功能研究Table 1 Fermentation and function of microorganism from traditional fermented seafood products
从研究传统发酵海产调味品中微生物的区域来看,主要有泰国、马来西亚、菲律宾、印度尼西亚等东南亚国际以及中国、韩国和日本等东亚国家,也包括加纳、埃及和巴西等非亚洲国家。研究比较广泛和深入的国家首选泰国,泰国海产调味品种类众多,包括Plaa-som、Kungchao、Nam-pla 和 Pla-ra 等[35],但海产调味品中微生物的种类和发酵作用功能方面差异性很大。例如Halobacillus sp.、T.halophilus和Virgibacillus sp.SK33三种嗜盐菌均可形成耐盐蛋白酶,其中T.halophilus主要对鱼酱产品的风味形成发挥主导作用,Virgibacillus sp.SK33的主要作用是加速发酵进程,缩短发酵周期。从鱼酱油和 anchovy fish sauce产品中分离的菌株 Brevibacillus sp.SK35和Natrinema gari HDS3-1可以有效地降低组氨酸含量,从fresh water fish中分离的L.plantarum IFRPD P15和L.reuteri IFRPD P17也作为发酵剂应用于缩短发酵周期和改善海产调味品质量方面。
马来西亚对海产调味品的微生物学研究主要是利用Staphylococcus carnosus FS19和Bacillus amyloliquefaciens FS05作为Budu和Bakasang等的发酵剂应用于生产实践中,降低产品中组胺的含量。在菲律宾鱼酱Tinabal和Balbakwa的生产中,微生物发酵作用主要在于形成浓郁的风味物质[36]。
韩国和日本的研究主要是在产品和发酵过程中分离特征性的微生物。例如,从韩国传统发酵海产品中分离得到的微生物,经鉴定包括从盐腌制的北美乌鱼中分离的Halomonas jeotgali sp.nov.以及腌虾中分离的 Natronococcus jeotgali sp.nov.[37-38]。在日本soy sauce moromi中分离出具有免疫调节作用的菌株Tetragenococcus halophilus Th221。
在我国,目前应用于发酵海产调味品中的研究主要还处在初级阶段,从产品和发酵过程中分离特征性嗜盐菌。在应用方面,以Kodamaea ohmeri M8菌株在25%NaCl时添加到鱼露发酵过程中,使组胺和酪胺的降解率分别达到64.4% 和72.4%[34]。
4 结论与展望
传统发酵海产调味品是各国人民智慧的结晶,长期以来,由于对发酵过程中微生物作用的认知不足,致使产品的品质和安全性的提升徘徊不前。然而,近些年经过各国微生物工作者的潜心研究,已经获得各种传统海产调味品中一定量的微生物学信息,同时也开展了微生物对蛋白质的降解作用、产品风味与香气物质形成和生物胺降解的作用和功能,进而利用特征性微生物菌株作为发酵剂改善产品品质和提升食品安全性应用研究。如此,对传统发酵海产调味品微生物学研究进入一个新的时期。然而,人们对传统发酵海产品极端环境下微生物的菌相构成、生存机制、发酵作用与功能认知才刚刚开始,未知的还很多,有待于科研工作者不懈的努力,对高盐环境下微生物的分布状况及与产品之间的相互作用关系进行分析与研究。从机理上探知微生物对产品品质的作用和影响,从应用上获取特征性微生物应用到发酵过程中,以降低盐度、增强风味、提高安全性为指标,达到缩短发酵周期,提高产品品质和经济效益的目的。
[1]Mehdi Z,Hossein N,Mohammad H E,et al.Chemical and microbial properties of mahyaveh,a traditional Iranian fish sauce[J].Food Control,2012,23(2):511-514.
[2]Saithong P,Panthavee W,Boonyaratanakornkit M,et al.Use of a starter culture of lactic acid bacteria in plaa-som,a Thai fermented fish[J].Journal of Bioscience and Bioengineering,2010,110(5):553-557.
[3]Udomsil N,Rodtong S,Tanasupawat S,et al.Proteinaseproducing halophilic lactic acid bacteria isolated from fish sauce fermentation and their ability to produce volatile compounds[J].International Journal of Food Microbiology,2010,141(3):186-194.
[4]Satomi M,Kimura B,Mizoi M,et al.Tetragenococcus muriaticus sp.nov.,a new moderately halophilic lactic acid bacterium isolated from fermented squid liver sauce[J].International Journal of Systematic Bacteriology,1997,47(3):832-836.
[5]Antonio V,Joaquin J N,Aharon O.Biology of Moderately Halophilic Aerobic Bacteria[J].Microbiology and Molecular Biology Reviews,1998,62(2):504-544.
[6]Sanchez P C,Klitsaneephaiboon M.Traditional fish sauce(patis)fermentation in the Philippines[J].Philippine Agriculturist,1983,66(3):69-251.
[7]Gupta A,Roy I,Patel RK,et al.One-step purification and characterization of an alkaline protease from haloalkaliphilic Bacillus sp.[J].Journal of Chromatography A,2005,1075(1-2):103-108.
[8]Kamekura M,Seno Y.A halophilic extracellular protease from a Halophilic archaebacterium strain 172 P1[J].Biochemistry and Cell Biology,1990,68(1):352-359.
[9]Sinsuwan S,Rodtong S,Yongsawatdigul J.A NaCl-stable serine proteinase from Virgibacillus sp.SK33 isolated from Thai fish sauce[J].Food Chemistry,2010,119(2):573-579.
[10]Christensen J E,Dudley E G,Pederson J A,et al.Peptidases and amino acid catabolism in lactic acid bacteria[J].Antonie van Leeuwenhoek,1999,76(1-4):217-246.
[11]Masson F,Hinrichsen L,Talon R,et al.Factors influencing leucine catabolism by a strain of Staphylococcus carnosus[J].International Journal of Food Microbiology,1999,49(3):173-178.
[12]Cha Y J,Cadwallader K R.Volatile components in saltfermented fish and shrimp pastes[J].Journal of Food Science,1995,60(1):19-24.
[13]Fukami K,Funatsu Y,Kawasaki K,et al.Improvement of fish sauce odor by treatment with bacteria isolated from the fish-sauce mush(moromi)made from frigate mackerel[J].Journal of Food Science,2004,69(2):45-49.
[14]綦翠华.值得开发的海产鲜味调味品[J].中国调味品,2006(1):85-90.
[15]Saisithi P,Kasemsarn B O,Liston J,et al.Microbiology and chemistry of fermented fish[J].Journal of Food Science,1966,31(1):105-110.
[16]Beddows C G,Ardeshir A G,Daud W J B.Development and origin of the volatile fatty acids in Budu[J].Journal of Science of Food and Agriculture,1980,31(1):86-92.
[17]Lonvaud-Funel A.Biogenic amines in wines:Role of lactic acid bacteria[J].FEMS Microbiology Letters,2001,199(1):9-13.
[18]Maintz L,Novak N.Histamine and histamine intolerance[J].The American of Clinical Nutrition,2007,85(5):1 185-1 196.
[19]Til H P,Falke H E,Prinsen M K,et al.Acute and subacutetoxicity of tyramine,spermidine,spermine,putrescine and cadaverine in rats[J].Food and Chemical Toxicology,1997,35(3-4):337-348.
[20]Tapingkae W,Tanasupawat S,Parkin K L,et al.Degradation of histamine by extremely halophilic archaea isolated from high salt-fermented fishery products[J].Enzyme and Microbial Technology,2010,46(2):92-99.
[21]Santos M H.Biogenic amines:their importance in foods[J].International Journal of Food Microbiology,1996,29(2):213-231.
[22]Haas H L,Sergeeva O A,Selbach O.Histamine in the nervous system[J].Physiological Reviews,2008,88(3):1183-1241.
[23]Michihata T,Kato D,Yano T,et al.Contents of polyamines in ISHIRU(fish sauce)[J].Journal of the Japanese Society for Food Science and Technology,2006,53(6):337-343.
[24]Kuda T,Miyawaki M.Reduction of histamine in fish sauces by rice bran nuka[J].Food Control,2010,21(10):1322-1326.
[25]Canadian Food Inspection Agency(CFIA).Fish Products Standard and Methods Manual[S].Guide to Food Labelling and Advertising,2009.
[26]Garcia-Ruiz A,González-Rompinelli E M,Bartolomé B,et al.Potential of wine associated lactic acid bacteria to degrade biogenic amines[J].International Journal of Food Microbiology,2011,148(2):115-120.
[27]Sinsuwan S,Montriwong A,Rodtong S,et al.Biogenic amines degradation by moderate halophile,Brevibacillus sp.SK35[J].Journal of Biotechnology,2010,150(Supplement):316.
[28]Zaman M Z,Abu Bakar F,Jinap S,et al.Novel starter cultures to inhibit biogenic amines accumulation during fish sauce fermentation[J].International Journal of Food Microbiology,2011,145(1):84-91.
[29]Dapkevicius M L N,Nout M J,Rombouts F M,et al.Biogenic amine formation and degradation by potential fish silage starter microorganisms[J].International Journal of Food Microbiology,2000,57(1):107-114.
[30]Xu W,Yu G,Xue C,et al.Biochemical changes associated with fast fermentation of squid processing by-products for low salt fish sauce[J].Food Chemistry,2008,107(4):1597-1604.
[31]Masuda S,Yamaguchi H,Kurokawa T,et al.Immunomodulatory effect of halophilic lactic acid bacterium Tetragenococcus halophilus Th221 from soy sauce moromi grown in high-salt medium[J].International Journal of Food Microbiology,2008,121(3):245-252.
[32]Sinsuwan S,Nawong S,Rodtong S,et al.Characterization of Virgibacillus sp.SK33 cell-bound proteinases and its application as a starter culture for fish sauce fermentation[J].Abstracts/Journal of Biotechnology,2008,136(Supplement):S717-S742.
[33]Yan Y,Qian Y,Ji F,et al.Microbial composition during Chinese soy sauce koji-making based on culture dependent and independent methods[J].Food Microbiology,2013,34(1):189-195.
[34]杨利昆,付湘晋,胡叶碧等.鱼露中生物胺降解菌的筛选及其特性[J].食品科学,2012,33(11):158-162.
[35]Rhee S J,Lee J E,Lee C H.Importance of lactic acid bacteria in Asian fermented foods[J].Microbial Cell Factories,2011,10(1):S5.
[36]Van Veen A G.In:fish as food(ed.G.Borgstrom).Fermented and dried seafood products in Southeast Asia[M].New York:Academic press,1965:227-250.
[37]Kim M S,Roh S W,Bae J W.Halomonas jeotgali sp.nov.,a new moderate halophilic bacterium isolated from a traditional fermented seafood[J].The Journal of Microbiology,2010,48(3):404-410.
[38]Roh S W,Nam Y D,Chang H W,et al.Natronococcus jeotgali sp.nov.,a halophilic archaeon isolated from shrimp jeotgal,a traditional fermented seafood from Korea[J].International Journal of Systematic and Evolutionary Microbiology,2007,57(9):2 129-2 131.
