Palaeobiogeographical constraints on the distribution of foraminifers and rugose corals in the Carboniferous Tindouf Basin, South Morocco
2013-09-27IanSomervillePedrozarIsmailSaidDanielVachardPaulaMedinaVareaSergioRodrguez
Ian D.Somerville , Pedro Cózar, Ismail Said, Daniel Vachard, Paula Medina‑Varea,Sergio Rodríguez,
1.UCD School of Geological Sciences, University College Dublin, Belfield, Dublin 4, Ireland
2.Dpto.de Geología Sedimentaria y Cambio Ambiental, Instituto de Geociencias (CSIC,UCM), c /José Antonio Novais 2,28040-Madrid, Spain
3.Dpto.de Paleontología, Facultad de Ciencias Geológicas, Universidad Complutense de Madrid, c /José Antonio Novais 2,28040-Madrid, Spain
4.Université de Lille 1, UMR 8217 du CNRS: Géosystèmes, Bâtiment SN 5, 59655 Villeneuve d′Ascq, France*
1 Introduction
The Tindouf Basin is situated south of the Atlas Transform Fault and the Anti-Atlas Mountains, and occupies the vast Tindouf Syncline.It constitutes one of the largest Carboniferous Saharan basins, together with the Béchar,Reggan, Taoudenni and Ahnet-Mouydir basins (Fig.1).The mostly Mississippian rocks of the Djebel Ouarkziz Formation form much of the northern limb of the syncline,which extends for about 300 km in a WSW-ENE direction between southern Morocco and Algeria, although the maximum length of the basin is about 500 km (Fig.1).
Despite the significance of the Tindouf Basin for the Carboniferous of North Africa and for the western Palaeotethys, studies in this difficult Saharan region are scarce,and most date from the 1960-1980s period.The database included in those pioneer studies is not sufficiently detailed for accurate faunal comparisons, because their focus was essentially biostratigraphic.
Owing to the spectacular landscape and excellent exposures in the Djebel Ouarkziz mountain range (Fig.2),references can be found to this region in the literature, with brief macrofaunal descriptions (a summary was included in Mametet al., 1966).Subsequently, the most important studies including data on foraminifers have been published by Mametet al.(1966), Lys (1979, 1985), Conrad (1985)and Sebbaret al.(2000).Publications dealing with rugose corals are limited to Menchikoff and Hsu (1935), Fabre(1955), Semenoff-Tian-Chansky (1974, 1985), Legrand-Blainet al.(1989)and Aretz (2011).
The stratigraphic section studied in the northern Tindouf Basin is a composite section, combining three measured sections of the Djebel Ouarkziz Formation (>450 m thick; Fig.3)located up to 27 km apart in total distance.They comprise a central section along the main Assa to Zag road, the Tinguiz Remz valley and wadi section, 17.5 km to the east, and a track in the reg situated 9.5 km to the west of the main road.In total, up to 29 carbonate horizons (labelled 1-2 and A-V; Fig.3)have been analyzed.Most of them (beds 1 to R; Fig.3)are laterally persistent between the three sections and they impart a marked cyclic character to the succession (Figs.2, 3).Each cyclic sequence is composed generally of a thick shale interval, locally with sandstone, and with carbonates (limestone and dolostone).None of the cycles is veneered by coal/palaeosol or capped by a palaeokarst.The proportion of shales in the cycles decreases toward the upper part of the sections, where locally reddish and yellowish marls occur.The entire lower member of the Djebel Ouarkziz Formation is intersected in those sections, but the marine carbonates of the middle and upper members of the Formation (beds S1-V)are only discontinuously exposed (Fig.3).A horizon of gypsum was recorded by Mametet al.(1966)in the Tinguiz Remz section, but this is no longer exposed.The top limestone bed in the Djebel Ouarkziz Formation (bed 20 in Mametet al., 1966, Fig.2)is well exposed in the western section and is represented by a thick coral biostrome (bed U), overlain by a thin sandy argillaceous limestone with brachiopods(bed V)(Fig.3).An older marine shaly and sandy interval containing moulds of brachiopods and crinoids with common trace fossils (Betaina Formation of Tournaisian-Late Viséan age), and the youngest continental conglomerates and sandstones (Betana Beds of Pennsylvanian age), are well intersected in these sections, but they are outside the main aims of this study.
The analyzed succession spans an interval with carbonates from the Late Asbian (Upper Viséan)up to the Early Bashkirian (Lower Pennsylvanian).In analyzing the published palaeobiogeographical models and palaeogeographical reconstructions, the Tindouf Basin is considered,like other Saharan basins, to be situated south of the peri-Gondwanan Mississippian basins in North and Central Morocco (Central Meseta), South Europe (SW Iberia)and Laurussia (N France, Belgium, UK and Ireland).During the Late Viséan, basins such as those in Ireland and Britain were situated 10° south of the palaeoequator, and in the Serpukhovian those basins were situated approximately at the palaeoequator (e.g., Scotese, 2002).The position of the Tindouf Basin is not so precisely known, but it could range from 25°-35° south in the Late Viséan to 20°-30°south in the Serpukhovian (Blakey, 2007).Owing to the palaeolatitudinal north-south location of the basins, the proposed faunal migration models were extremely simplified to north-south routes.As a consequence, the rich and more diverse equatorial assemblages should have migrated southwards, crossing and colonizing the intermediate peri-Gondwanan basins and platforms (in the case of benthic faunas).These kinds of models, such as in Legrand-Blainet al.(1989), can be in part correct, but also in part oversimplified for the variations observed in the Tindouf Basin.The case of the rugose corals is not as complex as that for the foraminifers, but their assemblages also contain some unusual occurrences (Rodríguezet al., 2011).
The aim of this study is to demonstrate the complexity of taxa recorded in the foraminiferal and coral assemblages of the Tindouf Basin, where, in addition to the mostly cosmopolitan foraminifers, more unusual species and genera from NW Europe and Spain are also recorded withoutoccurrences in basins located geographically in intermediate positions.The variety of taxa suggests the presence of multiple migration routes for the marine benthic faunas(foraminifers, rugose corals and brachiopods)introduced into the Tindouf Basin, even distinct from other Saharan basins.
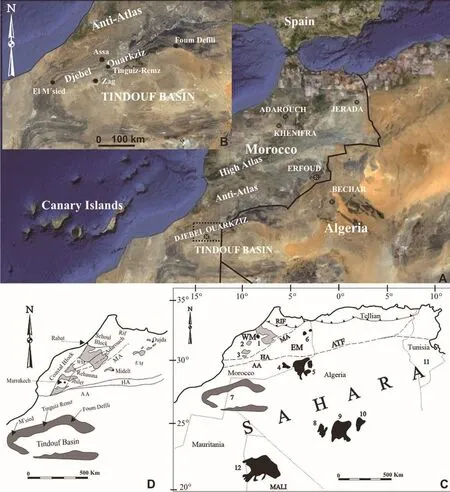
Fig.1 A-B-Aerial view of the northern flank of the Tindouf Syncline and Basin; C-Location of the main Carboniferous outcrops and basins in Morocco, Algeria, Mali and subsurface in Tunisia, 1-Central Meseta (Azrou-Khenifra), 2-Rehamna, 3-Jebilet, 4-Tafilalt,5-Béchar, 6-Jerada, 7-Tindouf, 8-Reggan, 9-Ahnet, 10-Mouydir, 11-Subsurface Carboniferous (number is situated in the map where the boreholes intersected Carboniferous rocks in Tunisia), 12-Taoudenni; AA-Anti-Atlas, ATF-Atlas Transform Fault, EM-Eastern Meseta, HA-High Atlas, MA-Middle Atlas, WM-Western Meseta; D-Detail of the Variscides and Tindouf Basin in Morocco; pale grey areas are Carboniferous outcrops, dark grey is the Tindouf Basin outcrop (modified from Cózar et al., 2011).

Fig.2 Main Assa-Zag road section.1-Field view looking west of the Djebel Ouarkziz escarpment, beds E to M; 2-View looking east of beds G to L showing the cyclicity.
2 Foraminifers with significant palaeobiogeographical constraints
2.1 Limitations of databases and physical barriers
This study is based on an examination of more than 800 thin-sections from carbonates sampled from the north Tindouf stratigraphic sections.Unfortunately, the material of other studies in North Africa was not published in detail.Thus, the first significant difference observed in Tindouf,a higher palaeodiversity compared to other Saharan basins, could be attributed to a less intense sampling programme elsewhere.The foraminiferal assemblages are extremely rich and diverse in most horizons in Tindouf,particularly those spanning the Late Brigantian (latest Viséan)to the Late Serpukhovian, which had less influence of terrigenous input in the limestone beds.The entire foraminiferal assemblages and detailed biostratigraphy will be published in forthcoming papers elsewhere.Lys (1979)described the Djebel Ouarkziz Formation succession near Foum Defili in neighbouring Algeria (Figs.1B, 1D), and we concur essentially with his biostratigraphic zonation.The only difference between his work and our study is in the precise positioning of the biozonal boundaries.
A factor that should be also taken into consideration for the reconstruction of the Tindouf platform conditions is the palaeobiogeographical dispersion of the benthic foraminifers.Modelling the interchange of fauna would simplify the controlling factors on the platform.However,those models are qualitative, and can only explain the palaeobiological relationships between the basins, and they cannot establish the precise location of geographical barriers or ocean currents.Nevertheless, these geographical models should respond to the palaeobiological constraints recognized on the platforms.Owing to the limited number of publications on Mississippian foraminifers in North Africa compared to those from Europe, this analysis cannot be focused on the lack of taxa, because this problem can be simply a function of the less intense sampling and
research in the different basins in North Africa, compared with their time equivalents in Europe.Moreover, our study can be only regarded as a preliminary interpretation, bearing in mind that the sections presented herein correspond only to c.30 km of the central part of the northern limb of the Tindouf Basin (Fig.1C).

Fig.3 Composite stratigraphic section in Tinguiz Remz and road to Zag.Location of the bed limestones labelled 2 to V.P-Pennsylvanian; B-Bashkirian; MDO-Middle Djebel Ouarkziz; UDO-Upper Djebel Ouarkziz.
Structurally, the Tindouf Basin can be closely compared with other western Saharan basins (e.g., Béchar,Ahnet-Mouydir, Reggan), all situated south of the Atlas Transform Fault and Anti-Atlas Mountains, which are generally considered as a major biogeographical barrier(Conrad, 1985).This barrier isolated the North Cratonic platforms of Gondwana (Sahara)from those platforms developed in the peri-Gondwanan terrains (e.g., SW Spain, Central Meseta, Pyrenees, Balearic Islands and Montagne Noire)located in an intermediate position,with the Laurussian platform of the extreme western part of Palaeotethys (e.g., N France, Belgium, Ireland, UK)farther north (Fig.1C).
2.2 Foraminiferal assemblages
Foraminifers in the Tindouf Basin can be grouped into four main categories: (1)the mostly cosmopolitan assemblages, or at least those which are well distributed in the western and central Palaeotethys; (2)the “missing” taxa in the Sahara, which refer to those foraminifers that are well-known in western Laurussian or peri-Gondwanan platforms, but are unknown in the Saharan basins; (3)the Saharan assemblages; and (4)the so-called “out of place”taxa, which refer to all those taxa recorded in Tindouf that are well-known in western European basins, but their migration routes to this southern margin of Palaeotethys are unknown, due to their apparent absence in intermediate basins, such as North and Central Morocco.
The well distributed foraminiferal genera of the Palaeotethys that are mostly cosmopolitan can be discarded for this analysis, since they do not contribute any significant data.A suite of those Palaeotethyan cosmopolitan taxa can be summarized as including diverse species of the following genera (listed alphabetically):Archaediscus,Asteroar‑chaediscus,Biseriella,Brenckleina,Brunsia,Calcitornella,Calcivertella,Climacammina,Consobrinella,Cribrospira,Cribrostomum,Earlandia,Endospiroplectammina,En‑dostaffella,Endotaxis,Endothyra,Endothyranopsis,Eo‑sigmoilina,Eostaffella,Eostaffellina,Koskinobigenerina,Koskinotextularia,Forschia,Forschiella,Globivalvulina,Globoendothyra,Howchinia,Janischewskina,Lituotu‑bella,Loeblichia,Neoarchaediscus,Nevillea,Nodosar‑chaediscus,Mediocris,Mikhailovella,Monotaxinoides,Omphalotis,Palaeotextularia,Planoendothyra,Plano‑spirodiscus,Plectogyranopsis,Plectostaffella,Pseudoam‑modiscus,Pseudoendothyra,Pseudoglomospira,Semino‑vella,TetrataxisandTubispirodiscus.
As mentioned above, the conclusions based on the“missing” taxa in the Sahara may be unreliable, because the sampling intensity and analyzed material for most other Saharan basins is not indicated.Moreover, despite the close geographical and palaeogeographical positions of these basins, their published foraminiferal assemblages suggest they are apparently poorer.However, it is possible that previous works may have documented only the biostratigraphically significant taxa.Hence, the foraminiferal assemblages recorded from other Saharan basins appear much less diverse than those recorded by us in the Tindouf Basin.
The other two categories, the “Saharan” and the “out of place” taxa recognized in the Tindouf Basin are considered to result from the direct influence of several other basins/regions, and a mixing of provenances.These provenances or influences can be summarized as Saharan, peri-Gondwanan (Central Meseta and SW Spain)and Laurussian (directly from the UK and surrounding areas of NW Europe).
2.2.1Saharan influence
It is obvious that there is a clear influence of other Saharan basins, from which many taxa arrived in Tindouf,which are unknown in the surrounding peri-Gondwanan terrains and Laurussian platforms.These include (Fig.4):Haplophragmina beschevensis,Archaediscus paraangu‑latus,Neoarchaediscus akchimensis,Bradyina nana,Eo‑sigmoilina explicata elongata,E.namuriensis,Eostaffella ikensis,Globivalvulina paraeogranulosa,G.minima and Howchiniasp.2 (= ?Howchinia beleutensisVdovenko).These are the most representative of a suite of foraminifers which occur in numerous sections elsewhere in the Sahara,mostly in the Béchar Basin (Lys, 1979; Sebbar and Lys,1989; Mametet al., 1994; Sebbar, 1997, 1998).In addition, other taxa (Fig.5)such as “Biseriella”scaphoidea[the occurrence of the diaphanotheca in this species is not clear enough, and thus, it should be considered also asGlo‑bivalvulina(see Vachardet al., 2006); nevertheless, specimens from Tindouf contain only a microgranular layer in the wall],Bradyina cribrostomata,Eostaffella mirifica,E.postproikensis,E.tenebrosa,Janischewskina delicata,Globivalvulina moderata,G.kamensis, species ofBrenck‑leinaandEosigmoilina, seem to be more abundant in the neighbouring Saharan basins than in peri-Gondwanan ter-rains or Laurussian platforms.

Fig.4 Influence of the Saharan basins (scale bar = 100 µm).1-Haplophragmina beschevensis (Brazhnikova), bed L, sample Pc4101,Early Serpukhovian; 2-Bradyina nana Potievskaya, bed M, sample Pc4119, Early Serpukhovian; 3-Eostaffella ikensis Vissarionova,bed L, sample Pc4110, Early Serpukhovian; 4-Archaediscus paraangulatus (Manukalova-Grebenyuk), bed J, Pc4098, Late Brigantian; 5-Eosigmoilina namuriensis (Dain), bed S5, sample Pc4160, Late Serpukhovian; 6-Neoarchaediscus akchimensis (Grozdilova and Lebedeva), bed P, sample Pc4138, Late Serpukhovian; 7-Eosigmoilina explicata elongata (Ganelina), bed S3, sample Pc4157,Late Serpukhovian; 8-Howchinia sp.2 sensu Lys (= ?Howchinia beleutensis Vdovenko), bed M, sample Pc4114, Early Serpukhovian;9-Globivalvulina paraeogranulosa (Cózar and Somerville), bed M, sample Pc4120, Early Serpukhovian; 10-Globivalvulina minima(Reitlinger), bed U, sample Pc4203, Bashkirian.
2.2.2Peri-Gondwanan influence
The peri-Gondwanan influence(s)can be focused on two terrains, the Moroccan Central Meseta and SW Spain.Taxa such asEndothyranopsis eocompressa,Endothy‑ranopsis pechorica,Kasachstanodiscus bestuvensisandVissarionovella donzelliare only recorded in this extreme western sector of the Palaeotethys in SW Spain (Cózar,2003 and references therein)and Tindouf (Fig.6).Some of them are rarely recorded also in the Central Meseta(Endothyranopsis eocompressaandEndothyranopsis pe‑chorica).Furthermore,Vissarionovella, is common in SW Spain (Cózar and Vachard, 2001)and present in the Montagne Noire (S France: Pille, 2008), but it is rare in the UK,and the Central Meseta.Representatives of this genus (e.g.,V.tujmasensis), have been never documented in the Saharan basins.The closest provenance for this genus seems to be SW Spain.These data suggest a direct interchange of fauna between SW Spain and Tindouf.Other taxa that are recorded in the Central Meseta and SW Spain, as well as Tindouf are:Euxinita pendleiensis,Praeostaffellina mac‑donaldensisandPraeplectostaffella asymmetrica, which are much more common in the UK and Ireland (Cózaret al., 2008a), but they rarely occur in the peri-Gondwanan terrains, less commonly in Tindouf.Thus, their abundance decreased markedly from the palaeoequator to subtropical palaeolatitudes to the south.
The exclusive influence of the Central Meseta is observed in taxa such asEndostaffella? spp., “Millerella”spp.,Parabiseriella moroccoensis,Kasachstanodiscus bykovae,PseudocornuspiraandTubispirodiscus simpli‑cissimus(see Cózaret al., 2008b, 2011).The former taxa are unknown in Algeria (Béchar)(based on data in Sebbar and Lys, 1989; Mametet al., 1994; Sebbar, 1997, 1998),even though some of them are only rarely recorded in western European basins.
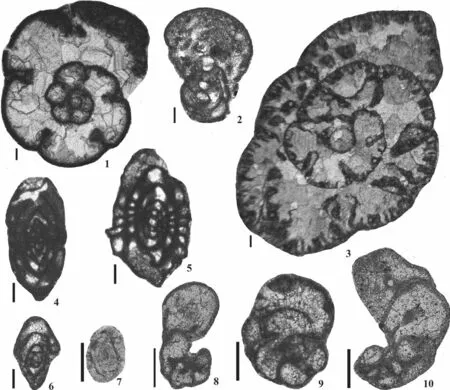
Fig.5 Common Saharan taxa (scale bar = 100 µm).1, 2-Janischewskina delicata Malakhova, bed M, samples Pc4117, Pc4119,Early Serpukhovian; 3-Bradyina cribrostomata Rauzer-Chernoussova and Reitlinger, bed M, sample Pc4118, Early Serpukhovian;4-Eostaffella mirifica Brazhnikova, bed M, sample Pc4118, Early Serpukhovian; 5-Eostaffella tenebrosa Vissarionova, bed M,sample Pc4117, Early Serpukhovian; 6-Eostaffella postproikensis Vdovenko, bed K, sample Pc4101, Late Brigantian; 7-Brenck‑leina rugosa (Brazhnikova), bed O, sample Pc4129, Late Serpukhovian; 8-“Biseriella” scaphoidea (Reitlinger), bed U, sample Pc4203, Bashkirian; 9-Globivalvulina moderata Reitlinger, bed P, sample Pc4141, Late Serpukhovian; 10-Globivalvulina kamensis Reitlinger, bed U, sample Pc4196, Bashkirian.
In addition, compared to the rich assemblages of the Central Meseta of Morocco in Adarouch (Cózaret al., 2008b,2011), some of the taxa that occur there (Vissarionovellatujmasensis,Endothyranopsis pechorica,E.eocompressa,Eostaffella tenebrosa,Biseriella paramoderata,Globival‑vulina paraeogranulosa,Loeblichia ukrainica,Pseudoen‑dothyra illustria,Eostaffella mutabilis,E.amabilis and Howchiniasp.2)are rarely recorded in western Laurussia.Some of these taxa are not recorded in SW Spain, but, surprisingly, some occur in the Pyrenees.It could be suggested that they reached the Pyrenees via Algeria or vice versa.However, they do not all occur in Béchar or other Saharan basins.This problem is attributed to possible misidentifica-tions of the taxa by previous workers.
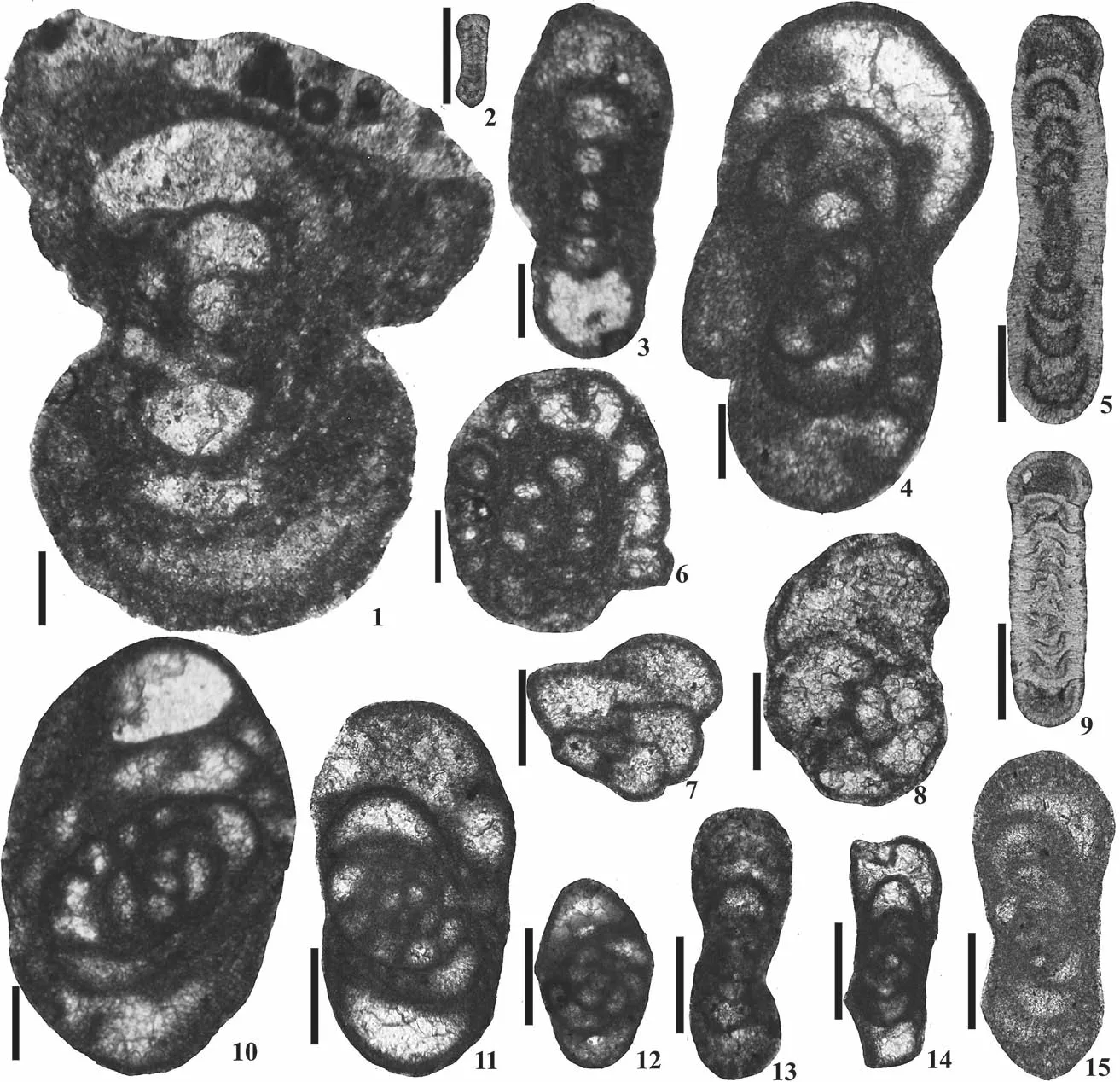
Fig.6 Taxa of a western peri-Gondwanan influence (scale bar = 100 µm).1-Endothyranopsis pechorica (Rauzer-Chernoussova),bed L, sample Pc4107, Early Serpukhovian; 2-Juvenile of Tubispirodiscus simplicissimus (Browne and Pohl), bed J, sample Pc4099,Late Brigantian; 3-Endothyranopsis eocompressa (Skipp), bed F, sample Pc4087, Early Brigantian; 4-Vissarionovella tujmasensis(Vissarionova), bed L, sample Pc4106, Early Serpukhovian; 5-Kasachstanodiscus bestubensis (Marfenkova), bed A, sample Pc4151,Late Asbian; 6-Euxinita pendleiensis (Cózar et al.), bed L, Pc4106; 7-Biseriella paramoderata (Cózar and Somerville), bed S4, sample Pc4159, Late Serpukhovian; 8-Parabiseriella moroccoensis (Cózar and Somerville), bed M, sample Pc4115, Early Serpukhovian;9-Kasachstanodiscus bykovae (Marfenkova), bed J, sample Pc4099, Late Brigantian; 10-Vissarionovella donzelli (Cózar and Vachard), bed M, sample Pc4113, Early Serpukhovian; 11-Praeostaffellina macdonaldensis (Cózar et al.), bed L, sample Pc4104, Early Serpukhovian; 12-Praeplectostaffella asymmetrica (Rozovskaya), bed M, sample Pc4114, Early Serpukhovian; 13-Millerella? sp.(cf.Cózar et al., 2011), bed N, sample Pc4125, Early Serpukhovian; 14-Endostaffella? sp.1 (cf.Cózar et al., 2011), bed L, sample Pc4111,Early Serpukhovian; 15-Endostaffella? sp.2 (cf.Cózar et al., 2011), bed L, sample Pc4106, Early Serpukhovian.
2.2.3Laurussian influence
The Laurussian influence is also highly marked in the assemblages of Tindouf, particularly from basins in the UK and Ireland, with taxa such asProtoinsolentitheca,Parabradyina,Parajanischewskina,Pojarkovella,Spino‑thyra,Bibradya,Globivalvulina bulloidesand Archaedis-cidae nov.gen.(Fig.7).Protoinsolentitheca,Parabradyi‑naandParajanischewskinaare common in Ireland and the UK, whereasProtoinsolentithecaandParajanischewskinaare rare in southern France (Vachard and Cózar, 2004;Cózar and Somerville, 2006).Protoinsolentithecais unknown in SW Spain, but it has been recorded in the Central Meseta (Oulmés region, Vachard and Tahiri, 1991;Adarouch region, Cózaret al., 2011).In the Saharan basins, these three genera are also unknown.Pojarkovellais common in the UK and Ireland, from the Holkerian up to the Brigantian.It is present in S France, but is rare in SW Spain (even represented by endemic species)and is known in the Holkerian of the Central Meseta (Vachard and Tahiri, 1991).Furthermore, an imprecise “Pojarko‑vella/Nibelia” is observed in the stratigraphic range charts of Mametet al.(1994)for the Viséan of Béchar.Spino‑thyraandBibradyaare rare in Ireland and the UK, and not recorded in other intermediate basins, even in the Sahara.They seem completely lacking in S France despite numerous investigations in this region.Only one specimen ofBibradya inflatais present in SW Spain (Sánchezet al.,1991, pl.3, fig.2, asJanischewskinasp.).Globivalvulina bulloidesand Archaediscidae nov.gen.are only known in the Midland Valley and Archerbeck Borehole (Solway Basin)of Scotland (Brady, 1876; Cózaret al., 2008a, 2010;Cózar and Somerville, 2013), and thus they may have migrated from the UK.
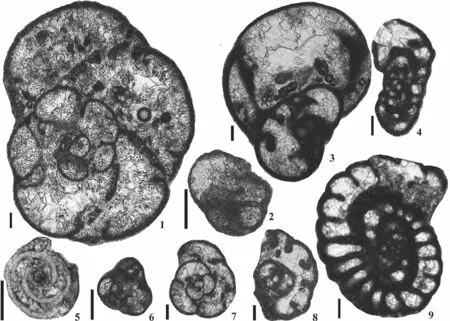
Fig.7 Taxa of a Central European influence (scale bar = 100 µm).1-Praejanischewskina brigantiensis (Cózar and Somerville), bed P, sample Pc4141, Late Serpukhovian; 2-Globivalvulina bulloides (Brady), bed U, sample Pc 4203, Bashkirian; 3-Mirifica mirifica(Rauzer-Chernoussova), bed P, Pc4141, Late Serpukhovian; 4-Pojarkovella nibelis (Durkina), bed O, sample Pc4136, Late Serpukhovian; 5-Archaediscidae new genus (cf.Cózar et al., 2010), bed K, Pc4101, Late Brigantian; 6-Protoinsolentitheca fundamenta(Vachard and Cózar), bed E, sample Pc4081, Early Brigantian; 7-Spinothyra pauciseptata (Rauzer-Chernoussova), bed L, sample Pc4111, Early Serpukhovian; 8-Bibradya inflata Strank, bed P, sample Pc4139, Late Serpukhovian; 9-Pojarkovella n.sp.2, bed K,simple Pc4100, Late Brigantian.
2.3 Tectonic constraints
Some of the taxa highlighted above suggest a rathercomplex scenario, with multiple possible migration routes to be incorporated in any reconstruction.The lithostratigraphical, tectonic and palaeogeographical data of the regions cannot be ignored.The Palaeozoic rocks in SW Spain represent an amalgamation of terrains, separated by strikeslip faults with a dominant sinistral movement, representing part of the Variscan Orogen (Blakey, 2007).Strikeslip basins and sinistral displacement along the main faults were demonstrated to be active during the Serpukhovian(Cózar and Rodríguez, 2004; Cózaret al., 2006), even up to the Duckmantian in continental basins, and some faults in SW Spain were considered as transform faults.The sinistral displacement along those faults was estimated to be hundreds of kilometers (Wagner, 1999).In contrast, the Central Meseta in Morocco moved along the South Atlas Transform Fault, with two dextral displacements during the Tournaisian and Early Viséan, and a final sinistral movement in the Late Viséan.According to recent studies,this would place more or less the Central Meseta in its current position relative to the Palaeozoic rocks south of the Atlas Transform Fault (Hoepffneret al., 2005; Michardet al., 2008).This model implies that the tectonic events were completed before the Serpukhovian.However, recent data suggest that the configuration of the Azrou-Khenifra Basin in the Adarouch area of northern Central Morocco(Fig.1)was not established until after the Serpukhovian,which was still tectonically active, although displacement on the faults were not quantified (Cózaret al., 2008b,2011).Owing to the significant biogeographical and lithostratigraphical differences between basins north and south of the Atlas Transform Fault, the scenario might be similar to that in SW Spain, where hundreds of kilometres of lateral displacement(s)along the transform fault could be involved.This active tectonism during Serpukhovian times can be also supported for SW Spain and the Central Meseta by the absence, in both areas, of the classical cycles (cyclothems).These comprise a regular alternation of thin carbonates and thicker siliciclastic rock intervals,which are observed in the Tindouf region and in the western Palaeotethys in more stable regions, such as the UK,Ireland, and the Maritime Provinces of Canada (Ramsbottom, 1973; von Bitteret al., 2007).Within those terrains or microplates can be also included the so-called South-Portuguese Zone (Lotze, 1945)and the Sehoul Block of northern Morocco (Fig.1D).Unfortunately, though, due to the predominantly deep water turbiditic nature of the successions, the palaeobiogeographical database from these areas is very poor.These two areas seem to be related to each other (Simancaset al., 2005).
3 Significant data on rugose coral ass‑emblages
Analysis of the distribution of Mississippian rugose corals from the Tindouf Basin has revealed some anomalies compared to other Carboniferous basins in North Africa and Europe (see Rodríguezet al., 2011, in press);they are summarized here.
Two colonial rugose genera and species,Lonsdaleia du‑plicataandActinocyathus floriformis, present in the Tindouf Basin, are unknown from northern Moroccan basins,but are common in NW Europe, Russia and NoVa Scotia(Poty, 2002; Rodríguez and Somerville, 2007; von Bitteret al., 2007)in the Upper Viséan and Serpukhovian.Also,Kizilia, a solitary rugosan coral is common in the Tindouf Basin, but is scarce in the Azrou-Khenifra Basin (N Morocco)and elsewhere in Europe.
Several rugose coral taxa appear later (i.e., in the Serpukhovian)in the Tindouf Basin compared to their first appearances in other basins in N Morocco and/or NW Europe.These includePalastraea regia,Tizraia berkhlii,Ac‑tinocyathus floriformis and Aulophyllum fungites, which typically first occur in the Brigantian of the Palaeotethys(Rodríguez and Somerville, 2007; Said and Rodríguez,2007).Moreover,Tizraia berkhliiappears to be endemic to North Africa, being recognized in the Azrou-Khenifra,Jerada and Tindouf Basins (Said and Rodríguez, 2007;Aretz, 2010; Saidet al., 2013).
The progressive increase in diversity and abundance of corals in assemblages throughout the Djebel Ouarkziz Formation (especially from the Late Brigantian to the Early Bashkirian; beds H to U, Fig.3)may reflect the continuing influence of favourable ocean currents for migration and improved marine connections with the Palaeotethys region during transgressions, as well as local factors, such as less detrital input.This pattern of increasing diversity and abundance through the formation is similar to that shown by the foraminifers.
The biostrome at the top of the Djebel Ouarkziz Formation is important not only for correlating between other Saharan basins (e.g., with Taoudenni, 800 km to the south,Reggan and Ahnet >1000 km to the southeast), but also for establishing that several rugose genera and species survived into the Early Bashkirian (Actinocyathus saritsche‑vae,Dibunophyllum bipartitum,Diphyphyllum maximum,Palaeosmilia murchisoni,Palastraea regia, andSiphono‑dendronspp.).Interestingly, several of these “Mississippian” genera have been reported from the Bashkirian inArctic Canada (Fedorowskiet al., 2012)and Nova Zemlya(Kossovaya, 1996).Inversely, elsewhere in western Europe and South China, many taxa disappeared at the end of the Serpukhovian during the Mid-Carboniferous crisis event (Kossovaya, 1996; Wanget al., 2006; McGheeet al., 2012).
Palaeogeographic reconstruction based on the distribution of rugose corals is difficult because of the apparent isolated position of the Tindouf Basin in Saharan Morocco compared to the central and northern Moroccan basins(e.g.,Azrou-Khenifra)north of the Anti-Atlas mountain range.The simple north-to-south marine corridor from southern Europe to the Sahara, as envisaged by previous workers (e.g., Legrand-Blainet al., 1989), is complicated by the absence of key taxa in North Morocco (Rodríguezet al., 2012; Saidet al., 2013).
Thus, there must have been, at times, some partial isolation of basins in North and Central Morocco and South Spain.A probable migration route to and from the Tindouf Basin may have also occurred to the northeast, in the Béchar Basin, but a westward connection to the Rheic Ocean is also a possibility, especially as many taxa in Tindouf also occur in NW Europe.This affinity with the rugose coral faunas of the British Isles and Belgium was noted by Semenoff-Tian-Chansky (1974, 1985).
4 Discussion
4.1 Migration routes and provinciality of foraminifers
In general, the main provenance for the Middle to Late Mississippian foraminifers is located in eastern Europe or further east (e.g., Vdovenko, 1980), as well as for the rugose corals (Fedorowski, 1981), which crossed into the western platforms of Laurussia in western Europe (Belgium, N France, UK and Ireland).From those basins can be intuitively proposed a main corridor to the Tindouf Basin, crossing between SW Spain and the Central Meseta of North Morocco, where the majority of the assemblages are present, but our data do not confirm this hypothesis.
It is necessary to take into consideration the non-preserved Carboniferous basins in the Betic Cordillera, the Balearic Islands and the Rif in northern Morocco (based on the presence of Carboniferous pebbles and boulders preserved in post-Early Bashkirian debris flows,e.g.,see Bourrouilh and Lys, 1978; Herbig and Mamet, 1985;Flügel and Herbig, 1988), together with the data from the Pyrenees and Montagne Noire (e.g., Vachard, 1977; Perret,1993; Pille, 2008).They all constitute a different microplate of peri-Gondwana, considered as a palaeobiogeographical subprovince within this sector of the western Palaeotethys Province (realm)(Fig.8).This is highlighted, in part, also by their endemic calcareous algae described in Vachardet al.(2008), Pille (2008)and Cózaret al.(2009).These features were used to define geographically the Eovelebitella Subprovince.This algal genus is well constrained to this subprovince, being absent in northwestern Palaeotethys platforms, western and eastern peri-Gondwanan terrains,as well as in the Saharan Gondwanan platforms.The single uncertain boundary for this subprovince is to the east,where algal assemblages could be questionably assigned to this subprovince as far as the Carnic Alps.According to Pille (2008), the calcareous algae of this Eovelebitella Subprovince had a much greater influence on, and affinity with, the eastern basins of the northern margin of Gondwana, such as in Morocco and Algeria, than in SW Spain,and the UK.This main Laurussian-Gondwanan corridor coincides with that proposed for brachiopod migrations by Legrand-Blainet al.(1989), denominated as the Western Mediterranean Variscan zones.Thus, the Eovelebitella Subprovince would overlap with that defined with brachiopods only in part.The poor knowledge on foraminifers in some of those outcrops (Balearic Islands, Moroccan Rif,Betic Cordillera)prevents further comparison.This agrees with the published palaeogeographical reconstructions(e.g., Scotese, 2002; Blakey, 2007).However, secondary routes are needed to explain the unusual biogeographical relationships between the foraminifers of those basins.
Some of the taxa recorded in SW Spain, the Central Meseta and Tindouf are not recorded in the UK or N France, but they are recorded in other Saharan basins (e.g.,Béchar).This is related to the influence of the Mediterranean Subprovince.This suggests that one of the main routes from eastern Europe was split by an emergent landmass situated between northern and southern France (see Scotese, 2002; Blakey, 2007, 2008), the Variscan mountain range.Thus, some foraminifers and calcareous algae directly reached those North African basins, without any representatives present in NW European basins.The Montagne Noire microfossils show intermediate characteristics, but are clearly closer to the Mediterranean Subprovince (Fig.8).This route seems to be also the dominant one for the rugose corals in Tindouf via Béchar.
There are some foraminiferal taxa that are exclusively shared by the Central Meseta and Tindouf, but they were located in neighbouring areas, and many local factors and currents could permit the interchange of fauna.It isnoteworthy that taxa which suggest a direct provenance of the foraminifers from SW Spain are not recorded in the Central Meseta.The latter aspect suggests that there also existed a corridor or current connecting directly SW Spain and Tindouf (Fig.8).

Fig.8 Sketch model of the inferred geographic position of the Mississippian foraminiferal subprovinces with the main migrations routes and faunal interchange in the western Palaeotethyan basins.The black area indicates regions with little or absence of faunal interchange, possibly related to the presence of unfavourable facies or areas of uplift.
The most striking route seems to have been established directly from the UK to Tindouf, without involving any intermediate basin(s).This migration can be only related to ocean currents, allowing the transport of the foraminifers, as well as overprinting the glacioeustatic influence of Gondwanaland in all those tectonically passive margins.Those basins have to be open to the west, and were connected somewhere with the Rheic Ocean (Fig.8).Thus, an oceanic influence was more marked there than in the intermediate basins, where emergent landmasses and platforms modified oceanic currents.This corridor is inconsistent with all the published palaeogeographic reconstructions,and it suggests that the Variscan mountain range was not a continuous emerging landmass, and in turn, some marine connections had to exist through this potential barrier.
In summary, four subprovinces can be proposed for this western extremity of the Palaeotethys Province for the Mid-Late Mississippian (Fig.8): the Atlantic Subprovince(to the NW), the Mediterranean Subprovince (to the NE)and the Saharan Subprovince (to the E and SE).These three subprovinces enclose a fourth subprovince, referred to as the West peri-Gondwanan Subprovince.
The Atlantic Subprovince includes basins in Ireland,UK, Belgium, N France, and the Maritime Provinces of E Canada (Fig.8)(which can also be identified with rugose corals; see Fedorowski, 1981; Hill, 1981; Poty, 2002;von Bitteret al., 2007).Owing to their faunal similarities,the peri-Gondwanan terrains of Sierra Morena (SW Spain)and Central Meseta of North Morocco, show a closer affinity to those of the Atlantic Subprovince than further to the south in the Sahara.Nevertheless, the foraminiferal assemblages are sufficiently different, as to constitute a fourth subprovince, the West peri-Gondwanan Subprovince.Assemblages of the latter subprovince also suggest the existence of marine straits or corridors through the developing Variscan Mountains range.
The Mediterranean Subprovince includes peri-Gondwanan basins in Montagne Noire and the Pyrenees.According to the calcareous algae, this subprovince includes also SE Spain, Balearic Islands and the Rif of northern Morocco (Fig.8), constituting the main bulk of the Eovelebitella Subprovince of Pille (2008).However, the scarce current knowledge on foraminifers from the latter outcrops prevents both subprovinces being synonymized.The provenance of some taxa in the Mediterranean Subprovince can be considered as coming directly from eastern European basins, and with less interchange between the NW European basins.However, this domain also shows some interchange of fauna with other peri-Gondwanan terrains(e.g., SW Spain), and a frequent interchange with the cratonic platforms of Gondwana.
The Saharan Subprovince includes Tindouf, Béchar,Reggan and Ahnet-Mouydir (Fig.8), as well as some other basins in the western Sahara;e.g., Taoudenni and Libya(Massa and Vachard, 1979).
The shared endemic or partly-endemic taxa suggest that there is little interchange of fauna between the Mediterranean and the Atlantic Subprovinces (Fig.8).This can be readily explained by the emerging landmasses of the Variscan Mountains (Scotese, 2002), which split this sector of the western Palaeotethys into northern and southern domains.There is also some interchange of fauna between the Mediterranean Subprovince and the West peri-Gondwanan Subprovince.No landmass is supposed to be present there(e.g., Legrand-Blainet al., 1989; Scotese, 2002).However,the presence of deep-water shales and turbidite facies in the Pyrenees and NW Spain suggests that too great a water depth, as well as the absence of currents did not permit the migration of the benthic fauna.A similar case can be suggested for the very limited or non-existent faunal inter-changes between the West peri-Gondwanan Subprovince(Central Meseta/SW Spain)and most of the Saharan basins, except for Tindouf.This deep water area corresponds to the later formed Atlas Mountains.The Tindouf Basin seems to be the best connected area with West peri-Gondwana and Laurussia.This might be due to the east-west orientation of the Tindouf Basin, with an open ocean to the west, where ocean currents could exist.This area was denominated as the Tindouf Gulf by Choubert (1952).The rest of the Saharan basins seem to be open to the north(e.g., Scotese, 2002; Blakey, 2007)with a different palaeogeographic orientation.This different orientation only allowed the migration of the benthic faunas between Tindouf and Béchar.The Mediterranean Subprovince had to be connected by means of currents with the Saharan basins, but owing to the poor database in most of them, it is not yet understood which basin provided the passage for those faunas.It must be assumed that the linking basin was Béchar, which was situated in lower palaeolatitudes.Thus,it can be suggested that, subsequently, the neighbouring basin should be Tindouf.Further investigations are needed to substantiate this potential corridor.
4.2 Palaeobiogeographic and palaeoclimatic factors
Perhaps the main challenge is to try to reconstruct the exact position of these subprovinces, and in particular, the position of the mobile belts and basins.Palaeolatitudinal data are sparse in most of those areas, and the most reliable data place Ireland and Britain 10° south of the palaeoequator during the Viséan;i.e., in the main zone of tropical shallow-water carbonate production, and approximately at the equator during the Serpukhovian.The present-day difference in latitude between the studied basins is around 25°,which would place the Saharan basins in an approximate palaeolatitudinal position around 35° south for the Viséan and 25° south for the Serpukhovian;i.e., almost beyond the main zone of tropical carbonate production.Evaluating the thick to extremely thick carbonate successions (including reefs and oolites)and evaporites in some Saharan basins (e.g., Béchar and Tindouf; see Pareyn, 1959, 1961;Mametet al., 1966; Conrad, 1985; Bourqueet al., 1995),suggests that this palaeolatitudinal position does not seem all that plausible.It could be estimated that those basins should be located less than 20° south of the equator, within the southern tropical belt, in areas with maximum production of carbonates (see Scholleet al., 1983; Kiesslinget al., 2003).In addition, plate tectonics may have increased the present-day distance of these basins, or the palaeolatitudinal position of the UK and surrounding basins was farther north of the equator during the Serpukhovian.This palaeolatitude could be reduced more, if we take into account that Carboniferous carbonate basins in Algeria are situated in the centre of the Sahara desert, south of the Atlas Transform Fault, and some occur even further south in Mauritania and Mali (see Conrad, 1985; Fig.1).Unfortunately, we do not know the magnitude of the lateral displacements of the transform faults observed in SW Iberia and the Atlas Mountains.It is possible that the resulting distance between the UK and the Sahara could have been reduced after the Variscan orogenesis.However, during most of the Mississippian times, those areas were extensive basins, and the distance could be larger.Those data might be in agreement with the published maps of Scotese(2002).However, we differ from him significantly in our palaeoclimatic reconstruction, because (1)he positioned the Saharan basins in the arid climatic zone, whereas the fauna and microflora of the platforms clearly suggest that,for the most part, they lay within the more humid tropical zone.Only in the lower levels of the Serpukhovian,could an arid climate be permissible for this area, based on the presence of gypsum deposits in the Tindouf, Reganne and Ahnet Basins.In addition, (2)this reconstruction is in disagreement with the palaeodiversity observed in the Sahara that should be greater near the palaeoequator than in higher palaeolatitudes.The foraminiferal palaeodiversity of the Saharan basins is similar or even higher than in basins apparently situated at the palaeoequator, and readily,much higher than in the intermediate mobile belts to the north of the Atlas Transform Fault.
As a secondary consequence of this relative position of the basins, the influence of the supposed glaciations on those faunas is considered irrelevant for the Gondwanan platforms.This is because the higher amount of survivors of the Viséan/Serpukhovian crisis should be located close to the palaeoequator, further north of the main nuclei of ice on Gondwanaland (Scotese, 2002).Yet, analysis of the Tindouf Basin faunas clearly shows that the result is the opposite.There is a higher proportion of survivors and some of these constitute the longest stratigraphic ranges of foraminiferal taxa in the Saharan basins (a feature also mirrored by rugose corals).This fact also reinforces the models of oceanic currents as one of the main controlling factors for the distribution of the taxa in these subprovinces, as well as the establishment of more favourable platform conditions within a tropical setting for a longer duration.This would involve not only higher water temperatures on the platform, but also greater tectonic stability for these intracratonic basins of northern Gondwana,with periodic transgressions introducing rich and diverse foraminiferal assemblages.
5 Conclusions
The northern flank of the Tindouf Syncline in southern Morocco contains a continuous, well exposed Carboniferous succession, with limestones of Late Asbian to Early Bashkirian age containing rich and diverse foraminiferal and rugose coral assemblages.These provide new faunal data on the poorly known Saharan basins.
The palaeobiogeographical relationship of the Tindouf Basin with the Palaeotethyan basins is complex.Although there is a predominance of cosmopolitan taxa for the Palaeotethys, it is recognized that there was an influence of basins in NW Europe, such as Britain and Ireland.Some taxa are recorded in both NW Europe and Tindouf without any contributions from basins located within intermediate areas.Also, there is a direct influence from SW Spain,yet some of these taxa are absent from the Central Meseta in North and Central Morocco, which was supposed to be the bridge between Europe and North Africa.The neighbouring Saharan basins in Algeria probably do not solve the problem, because they seem to present distinct foraminiferal assemblages.Analysis of the data suggests that this sector of the western Palaeotethys can be subdivided into four palaeobiogeographical subprovinces: the Atlantic Subprovince (UK, Ireland, N France, Belgium),the Mediterranean Subprovince (Pyrenees, Montagne Noire, Betic Cordillera, Rif, Balearic Islands), and the Saharan Subprovince (Béchar, Reganne, Ahnet-Mouydir and Tindouf).In between, areas of mixed faunal assemblages occur (Central Meseta, SW Spain), which constitute a fourth subprovince, the West peri-Gondwanan Subprovince.
One of the scarce but clear conclusions is that the Central Meseta of North Morocco cannot be considered as being emplaced in its current position during Mississippian times, although its final emplacement is still unknown.It was distinct from the northern Palaeotethyan basins as well as the southern Saharan basins.The data also suggest that the current distance between Laurussian (e.g., England)and Gondwanan (i.e., Tindouf)platforms was likely reduced during the Pennsylvanian, and/or the position of the Atlantic Subprovince could be located north of the palaeoequator.
Analysis of the Tindouf Basin faunas shows that there is a high diversity and abundance of foraminiferal taxa, with a higher proportion of survivors and longer stratigraphic ranges, features also mirrored by rugose corals.This emphasizes the longevity of the carbonate platform in a tropical setting, where periodic transgressions introduced new assemblages, and oceanic currents are interpreted as one of the main controlling factors for the distribution of the taxa.Moreover, not only were water temperatures on the platform higher, but also was the tectonic stability greater.It is considered that the effects of the first phases of the Gondwanan glaciation were minimal on the Tindouf faunas.
Acknowledgements
Field work was carried out thanks to the collaboration of the Department of Mine Development of the Ministère de l′Energie et des Mines of Morocco.Field work was funded by the Spanish Ministerio de Innovación y Ciencia (research project CGL2009-10340BTE)and ongoing research development (research project CGL2012-30922BTE).This paper has benef ited from the constructive comments of Prof.Franz Fürsich (Erlangen).
Aretz, M., 2010.Rugose corals from the upper Viséan (Carboniferous)of the Jerada Massif (NE Morocco): taxonomy, biostratigraphy, facies and palaeobiogeography.Palaeontologische Zeitschrift, 84(3): 323-344.
Aretz, M., 2011.Corals from the Carboniferous of the central Sahara(Algeria): the collection of “Marie Legrand-Blain”.Geodiversitas, 33(4): 581-624.
Blakey, R.C., 2007.North America Palaeogeographic Maps.http://jan.ucc.nau.edu/~rcb7/nam.html.
Blakey, R.C., 2008.Gondwana paleogeography from assembly to breakup; a 500 m.y.odyssey.Special Paper-Geological Society of America, 441: 1-28.
Bourque, P.A., Madi, A., Mamet, B.L., 1995.Waulsortian-type bioherm development and response to sea-level fluctuations: Upper Viséan of Béchar basin, western Algeria.Journal of Sedimentary Research, B65: 80-95.
Bourrouilh, R., Lys, M., 1978.Sédimentologie et micropaléontologie d′olistostromes et coulées boueuses du Carbonifère des zones internes bético-kabylo-rifaines (Méditerranée occidentale).Annales de la Société Géologique du Nord, 97(1): 87-94.
Brady, H.B., 1876.A monograph of Carboniferous and Permian foraminifera (the genus Fusulina excepted).Palaeontographical Society of London, 30: 1-166.
Choubert, G., 1952.Histoire géologique du Domaine de l′Anti-Atlas.Notes e Mémoires du Service Géologique du Maroc, 100: 77-194.
Conrad, J., 1985.North Africa-Northwestern and central Saharan areas: Tindouf Basin.In: Wagner, R.H., Winkler, R.H., Prins,C.F., Granados, L.F., (eds).The Carboniferous of the WorldII: Australia, Indian subcontinent, South Africa, South America and North Africa.IUGS Publication No.20.Madrid: Instituto Geológico y Minero de España/ Empresa Nacional Adaro de Investigaciones Mineras S.A, 325-327.
Cózar, P., 2003.Foraminiferal fauna and zonation from the Early Carboniferous of Guadiato Area (SW Spain): comparison with European and North African foraminiferal zonal schemes and its palaeobiogeographical implications.In: Ahr, W.M., Harris, P.M., Morgan, W.A., Somerville, I.D., (eds).Permo-Carboniferous carbonate platforms and reefs.Tulsa: SEPM, Special Publication 78, and AAPG, Memoir 83: 162-177.
Cózar, P., Rodríguez, S., 2004.Pendleian marine carbonates from SW Spain: sedimentology, biostratigraphy and structure.Geological Journal, 39: 25-47.
Cózar, P., Said, I., Somerville, I.D., Vachard, D., Medina-Varea, P.,Rodríguez, S., Berkhli, M., 2011.Potential foraminiferal markers for the Visean-Serpukhovian and Serpukhovian-Bashkirian boundaries -a case-study from Central Morocco.Journal of Paleontology, 85(6): 1105-1127.
Cózar, P., Somerville, I.D., 2006.Significance of the Bradyinidae and Parajanischewskina n.gen.for biostratigraphic correlations in the late Viséan (Mississippian)in western Paleotethyan basins.Journal of Foraminiferal Research, 36: 262-272.
Cózar, P., Somerville, I.D., 2013.The Carboniferous Archerbeck Borehole, near Canonbie (Dumfriesshire, southern Scotland):biostratigraphic revision of the late Asbian to early Pendleian succession using foraminiferans and regional correlations.Earth and Environmental Sciences, Transactions of the Royal Society of Edinburgh, 103, in press.
Cózar, P., Somerville, I.D., Rodríguez, S., Mas, R., Medina-Varea,P., 2006.Development of a late Viséan (Mississippian)mixed carbonate/siliciclastic platform in the Guadalmellato Valley(southwestern Spain).Sedimentary Geology, 183: 269-295.
Cózar, P., Somerville, I.D., Burgess, I., 2008a.New foraminifers in the Viséan/Serpukhovian boundary interval of the Lower Limestone Formation, Midland Valley, Scotland.Journal of Paleontology, 82: 906-923.
Cózar, P., Somerville, I.D., Burgess, I., 2010.Foraminiferal, calcareous algal and problematica assemblages from the Mississippian Lower Limestone Formation in the Midland Valley, Scotland.Earth and Environmental Science Transactions of Royal Society of Edinburgh, 100: 297-309.
Cózar, P., Vachard, D., 2001.Dainellinae subfam.nov.(Foraminiferida du Carbonifère Inférieur), revision et nouveaux taxons.Geobios, 34: 505-526.
Cózar, P., Vachard, D., Somerville, I.D., Berkhli, M., Medina-Varea,P., Rodríguez, S., Said, I., 2008b.Late Viséan-Serpukhovian foraminiferans and calcareous algae from the Adarouch region (central Morocco), North Africa.Geological Journal, 43: 463-485.
Cózar, P., Vachard, D., Somerville, I.D., Pille, L., Medina-Varea, P.,2009.Revision and new species of the Late Palaeozoic dasyclad algaWindsoporellaandEovelebitella.Palaeontographica B, 282:40-67.
Fabre, J., 1955.Deux polypiers du Viséen supérieur de Tindouf (Sahara NW).Caninophyllum archiaci(M.Edw & H.)var.densanov.var.etCaninophyllum coronatumnov.sp.Annales de la société géologique du Nord, 75: 8-15.
Fedorowski, J., 1981.Carboniferous corals: distribution and sequence.Acta Palaeontologica Polonica, 26(2): 87-160.
Fedorowski, J., Bamber, E.W., Baranova, D.V., 2012.An unusual occurrence of Bashkirian (Pennsylvanian)rugose corals from the Sverdrup Basin, Arctic Canada.Journal of Paleontology, 86:979-995.
Flügel, E., Herbig, H.G., 1988.Mikrofazies karbonischer Kalkgerölle aus dem Paläozoikum des Rif (Marokko): ein Beitrag zur Paläogeographie der westmediterranen Paläotethys im Karbon.Facies,19: 271-300.
Herbig, H.G., Mamet, B.L., 1985.Stratigraphy of the limestone boulders, Marbella Formation (Betic Cordillera, Southern Spain).In: Compte Rendu 10 Congrès International de Stratigraphie du Carbonifère, Madrid, 1: 199-212.
Hill, D., 1981.Rugosa and Tabulata.In: Teichert, C., (ed).Treatise on Invertebrate Paleontology Part F (Supplement 1).Kansas:Geological Society of America and University of Kansas Press,Boulder Colorado and Lawrence, 2: 1-762.
Hoepffner, Ch., Soulaimani, A., Piqué, A., 2005.The Moroccan Hercynides.Journal of African Earth Sciences, 43: 144-165.
Kiessling, W., Flügel, E., Golonka, J., 2003, Patterns of Phanerozoic carbonate platform sedimentation.Lethaia, 36: 195-225.
Kossovaya, O., 1996.The mid-Carboniferous rugose coral recovery.In: Hart, M.B., (ed).Biotic Recovery from Mass Extinction Events, Geological Society London Special Publications, 102:187-199.
Legrand-Blain, M., Conrad, J., Coquel, A., Lejal-Nicol, A., Lys, M.,Poncet, J., Semenoff-Tian-Chansky, P., 1989.Carboniferous Palaeobiogeography of North Africa, In: Compte Rendu 11 Congrès International de Stratigraphie et Géologie du Carbonifère,Beijing, 4: 210-230.
Lotze, F., 1945.Zur Gliederung der Varisziden der Iberischen Meseta.Berlin: Teotektonishe Forschungen, 6: 78-92.
Lys, M., 1979.Micropaléontologie (Foraminifères)des formations marines du Carbonifère saharien.In: 8émé Congrès International du Stratigraphie du Carbonifère, Moscow, 2: 37-47.
Lys, M., 1985.North Africa -Foraminifera.In: Wagner, R.H., Winkler Prins, C.F., Granados, L.F., (eds).The Carboniferous of the World II: Australia, Indian subcontinent, South Africa, South America and North Africa.IUGS Publication, 20: 354-364.
Mamet, B.L., Choubert, L.G., Hottinger, G., 1966.Notes sur le Carbonifère du Jebel Ouarkziz.Étude du passage du Viséen au Namurien d´après les foraminifères.Notes du Service Géologique du Maroc, 27(198): 7-21.
Mamet, B.L., Madi, A., Bourque, P.A., Sebbar, A., 1994.Foraminifères carbonifères du Grand Erg occidental, Bassin de Béchar,Algérie.Bulletin de la Société belge de Géologie, 103: 51-61.
Massa, D., Vachard, D., 1979.Le Carbonifère de Lybie Occidentale:biostratigraphie et micropaléontologie; position dans le domainetéthysien d′Afrique du Nord.Revue Institut Français Pétrole, 34(1): 3-65.
McGhee, G.R.Jr., Sheehan, P.M., Bottjer, D.J., Droser, M.L.,2012.Ecological ranking of Phanerozoic biodiversity crises: The Serpukhovian (early Carboniferous)crisis had a greater ecological impact than the end-Ordovician.Geology, 40: 147-150.
Menchikoff, N., Hsu, T.Y., 1935.Les polypiers Carbonifères du Sahara Occidental.Bulletin Société Géologique de France 5, 5:229-261.
Michard, A., Hoepffner, C., Soulaimani, A., Baidder, L., 2008.Chapter 3, The Variscan Belt.In: Michard, A., Saddiqi, O., Chalouan,A., Frizon de Lamotte, D., (eds).Continental Evolution: The Geology of Morocco.Springer-Verlag Berlin Heidelberg: Lecture Notes in Earth Sciences 116, 65-131.
Pareyn, C., 1959.Les récifs carbonifères du Grand Erg occidental.Bulletin de la Société géologique de France, Série 7, 1: 347-364.
Pareyn, C., 1961.Les Massifs Carbonifères du Sahara Sud-Oranais.Publications du Centre de Recherches sahariennes, Série Géologie, 1: 1-324.
Perret, M.F., 1993.Recherches micropaléontologiques et biostratigraphiques (conodontes-foraminifères)dans le Carbonifère pyrénéen.Strata, série 2, Mémoires 21: 1-597.
Pille, L., 2008.Foraminifères et algues calcaires du Missississippien supérieur (Viséen supérieur-Serpukhovien): rôles biostratigraphique,paléoécologique et paléogéographique aux échelles locale, régionale et mondiale.Thèse Université de Lille, 3: 226p., 255p., 72 plates (or 150p.).
Poty, E., 2002.Stratigraphy and paleogeography of Carboniferous Rugose Corals of Nova Scotia.In: Hills, L.V., Henderson, C.M.,Bamber, E.W., (eds).Carboniferous and Permian of the World.Canadian Society of Petroleum Geologists, Memoir 19: 580-587.
Ramsbottom, W.H.C., 1973.Trangressions and regressions in the Dinantian: a new synthesis of British Dinantian stratigraphy.Proceedings of the Yorkshire Geological Society, 39: 567-607.
Rodríguez, S., Somerville, I.D., 2007.Comparisons of rugose corals from the Upper Viséan of SW Spain and Ireland: implications for improved resolutions in late Mississippian coral biostratigraphy.In: Hubmann, B., Piller, W.E., (eds).Fossil Corals and Sponges, Proceedings of the 9th International Symposium on Fossil Cnidaria and Porifera, Graz, 2003.Austrian Academy of Sciences, Schriftenreihe der Erdwissenschaftlichen Kommissionen 17: 275-305.
Rodríguez, S., Somerville, I., Said, I., Cózar, P., 2011.An upper Viséan (Asbian-Brigantian)coral succession at Djebel Ouarkziz(northern Tindouf Basin, southern Morocco).Abstracts, XI International Symposium on Fossil Cnidaria and Sponges, Liege,August 19-29, 2011.Kölner Forum Geologie und Paläontologie 19: 144-146.
Rodríguez, S., Somerville, I., Said, I., Cózar, P., 2012.Late Viséan coral fringing reef at Tiouinine (Morocco): implications for the role of rugose corals as building organisms in the Mississippian.Geological Journal, 47: 462-476.
Rodríguez, S., Somerville, I.D., Said, I., Cózar, P., 2013.An upper Viséan (Asbian-Brigantian)and Serpukhovian coral succession at Djebel Ouarkziz (northern Tindouf Basin, southern Morocco).Rivista Italiana di Paleontologia e Stratigrafia, in press.
Said, I., Rodríguez, S., 2007.Description ofTizraia berkhliigen.et sp.nov.(Rugosa)from Adarouch Area (Brigantian, NE Central Morocco).Coloquios de Paleontología, 57: 23-35.
Said, I., Somerville, I.D., Rodríguez, S., Cózar, P., 2013.Mississippian coral assemblages from the Khenifra area, Central Morocco:biostratigraphy, biofacies, palaeoecology and palaeobiogeography.Gondwana Research, 23: 367-379.
Sánchez, J.L., Comas-Rengifo, M.J., Rodríguez, S., 1991.Foraminíferos del Carbonífero de Los Santos de Maimona (Badajoz, SO de España).Boletín de la Real Sociedad de España de Historia Natural (Sección Geología), 86: 101-147.
Scholle, P.A., Bebout, D.G., Moore, C.H., 1983.Carbonate Depositional Environments.AAPG Memoir, 33: 1-708.
Scotese, C.R., 2002.PALEOMAP website.http://www.scotese.com.
Sebbar, A., 1997.Biostratigraphie (foraminifères)du Carbonifère Moyen, bassin de Béchar-Mezarif, Algérie.Annales de la Société Géologique de Belgique, 120: 205-215.
Sebbar, A., 1998.Foraminifères et Algues calcaires du Carbonifère,bassin de Reggane, Sahara central, Algérie.Bulletin du Service Géologique de l′Algérie, 9: 123-147.
Sebbar, A., Lys, M., 1989.Biostratigraphie du Carbonifère inférieur:Serpukhovien du Djebel Arlal, Bassin de Béchar, Algérie.Revue de Micropaléontologie, 32: 53-62.
Sebbar, A., Preat, A., Mamet, B.L., 2000.Microfaciès et biozonation de la rampe mixte carbonifère du bassin de Tindouf, Algérie.Bulletin du Centre de Recherches Elf Exploration et Production,22: 203-239.
Semenoff-Tian-Chansky, P., 1974.Recherches sur les Tétracoralliaires du Carbonifère du sahara Occidental.Ed.Cent.Natural Recherche Science Séries 6.Paris: Sc.Terre, 30: 1-316.
Semenoff-Tian-Chansky, P., 1985.Corals.In: Wagner, R.H., Winkler Prins, C.F., Granados, L.F., (eds).The Carboniferous of the World, II, Australia, Indian subcontinent, South Africa, South America and North Africa.IUGS Publication, 20: 374-381,431-435.
Simancas, J.F., Tahiri, A., Azor, A., Gonzalez Lodeiro, F., Martinez Poyatos, D.J., El Hadi, H., 2005.The tectonic frame of the Variscan-Alleghanian orogen in Southern Europe and Northern Africa.Tectonophysics, 398: 181–198.
Vachard, D., 1977.Etude stratigraphique et micropaléontologique du Viséen de la Montagne Noire (Hérault, France).Mémoires Institut géologique Université de Louvain, 29: 111-195.
Vachard, D., Cózar P., 2004.Insolentithecaemend.,Protoinsolen‑titheca, n.gen., and Caligellidae emends, permanent cysts of Palaeozoic Foraminifera? Rivista Italiana di Paleontologia e Stratigrafia, 110: 591-603.
Vachard, D., Gaillot, J., Pille, L., Blazejowski, B., 2006.Problems on Biseriamminoidea, Mississippian-Permian biserially coiled Foraminifera: A reappraisal with proposals.Revista Española de Micropaleontología, 38: 453-492.
Vachard, D., Pille, L., Aretz, M., Herbig, H.G., Cózar, P., 2008.Eovelebitella(dasyclad algae)and importance of its Brigantian subprovince within the western Palaeotethys.Kölner Forum für Geologie und Pälaontologie, 15: 120-121.
Vachard, D., Tahiri, A., 1991.Foraminifères, algues et pseudoalgues du Viséen de la région d´Oulmès (Maroc).Géologie Méditerranéenne, 43: 21-41.
Vdovenko, M.V., 1980, Vizeyskiy yarus.Zonal′noe raschlenenie i paleozoogeograficheskoe rayonirovanie (po foraminiferam)(The Viséan stage.Zonal subdivision and palaeozoogreographical zonation based on foraminiferans).Akademiya Nauk Ukrainskoy SSR, Institut Geologicheskikh Nauk, 175.
von Bitter, P.H., Giles, P.S., Utting, J., 2007.Biostratigraphic correlation of major cycles in the Windsor and Codroy groups of Nova Scotia & Newfoundland, Atlantic Canada, with the Mississippian substages of Britain and Ireland.In: Wong, T.E., (ed).Proceedings of the XVth International Congress on Carboniferous and Permian Stratigraphy.Amsterdam: Royal Netherlands Academy of Arts and Sciences, 513-533.
Wagner, R.H., 1999.Peñarroya, a strike-slip controlled basin of Early Westphalian age in southwest Spain.Bulletin of the Czech Geological Survey, 74: 3-24.
Wang, X.D., Wang, X.J., Zhang, F., Zhang, H., 2006.Diversity patterns of Carboniferous and Permian rugose corals in South China.Geological Journal, 41: 329-343.
杂志排行

Journal of Palaeogeography的其它文章
- The palaeobiogeography of South American gomphotheres
- The response of deltaic systems to climatic and hydrological changes in Daihai Lake rift basin,Inner Mongolia, northern China
- Review of research in internal-wave and internal-tide deposits of China
- Classifications, sedimentary features and facies associations of tidal f lats
- Mesozoic lithofacies palaeogeography and petroleum prospectivity in North Carnarvon Basin, Australia
- Facies-succession and architecture of the thirdorder sequences and their stratigraphic framework of the Devonian in Yunnan-Guizhou-Guangxi area,South China