硅对番茄生长及光合作用与蒸腾作用的影响
2013-09-12曹逼力徐坤石健辛国凤刘灿玉李秀
曹逼力,徐坤,石健,辛国凤,刘灿玉,李秀
(作物生物学国家重点实验室,农业部黄淮地区园艺作物生物学与种质创制重点实验室,山东农业大学园艺科学与工程学院,山东泰安271018)
硅是地壳和土壤中第二丰富的元素,但多以硅酸盐结晶或沉淀形式存在,能被植物吸收利用的有效硅含量较低[1]。植物中的硅多以二氧化硅胶(SiO2·nH2O)形态分布于表皮细胞和细胞壁,但物种间硅含量存在极大差异[2]。由于硅几乎无处不在,很难按Arnon等[3]的方法证明其为植物必需营养元素[4]。但Takahashi等研究表明,所有土壤种植的植物均含硅[5],且硅对水稻[6]、甘蔗[7]等多种植物的促长作用已被大量研究所证实,若去掉营养液中的硅元素会导致水稻[8]生长异常。Ma等[8]研究表明,水稻施硅可促进叶片及叶鞘表皮细胞“角质-双硅层”的形成,减少叶面蒸腾;供硅充足的水稻植株叶片叶绿素含量、气孔导度、净光合速率均显著提高[9]。丁燕芳等[10]研究表明,硅可提高干旱胁迫条件下小麦幼苗叶绿素含量,显著提高净光合速率、蒸腾速率及水分利用率。邹春琴等[11]研究证实,硅可降低向日葵叶片气孔导度、木质部汁液流速及蒸腾速率,提高水分利用率;硅亦可增加黄瓜幼苗叶片及表皮层、栅栏组织的厚度,降低叶片比表面积,减少水分蒸腾[12],并增强甜瓜叶片的光合速率,降低化瓜率,提高早期产量[13]。白菜施硅后,叶绿体排列更加紧密,基粒片层增加,光合速率和蒸腾速率均提高[14];大豆幼苗的光合速率及水分利用率随土壤有效硅含量增加的同时,其蒸腾速率也相应提高[15]。
番茄(Lycopersicon esculentum)是世界广泛栽培的主要蔬菜品种之一,合理施硅可显著促进番茄植株生长,提早开花,增加产量及果实 Vc含量[16]。Stamatakis等[17]的研究也证实,硅可显著提高番茄果实硬度及Vc、β-胡萝卜素、番茄红素等含量,并有效防止脐腐病的发生,但硅对番茄水气交换特性的研究鲜见报道。为此,本文研究了不同硅水平对番茄生长及叶片光合作用和蒸腾作用等的影响,以期揭示番茄对硅的响应特性,为生产中合理施用硅肥提供理论依据。
1 材料与方法
1.1 试验设计
本试验在2010年预备试验的基础上,于2011年在山东农业大学蔬菜试验站日光温室内进行。供试番茄品种为“粉蒂”,于2月18日穴盘基质育苗,至3叶1心时,用去离子水洗净根部基质,以聚乙烯泡沫板打孔固定植株,栽植于塑料盘(40 cm×30 cm×12 cm)内,每盘6株,采用Hoagland营养液进行培养。
试验设4个营养液硅(Si)水平,分别为:0 mmol/L(CK)、0.6 mmol/L(T1)、1.2 mmol/L(T2)、1.8 mmol/L(T3),每处理栽植9盘,共36盘,每12盘作为1个区组,共3次重复。供试营养液用去离子水配制,所用硅源为K2SiO3·nH2O,加硅处理由K2SiO3引入的K+通过调节Hoagland营养液中KNO3的用量进行扣除,由此引起的NO-3损失用稀硝酸补充。营养液pH值用0.01 mol/L的H2SO4和NaOH调节至6.0左右。番茄生长过程中,用气泵为营养液供氧,每3 d更换一次营养液。
1.2测定项目与方法
番茄生长至第一花穗开花时,取植株样,将各部位分开,测定株高、茎粗及根、茎、叶鲜质量,并分别置干燥箱中于105℃杀青30 min,75℃烘干至恒重,以Vorm法[18]测定植株硅含量。此外,选取植株上数第3片展开功能叶进行相关指标的测定:叶片色素含量采用80%丙酮提取,比色法测定[19];叶片光合速率(Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)和蒸腾速率(Tr)等水气交换参数用英国Hansatech公司生产的TPS-Ⅱ型光合仪测定,并根据相关参数,计算叶片瞬时水分利用效率(WUEi)和气孔限制值(Ls),WUEi=Pn/Tr,Ls=1-Ci/Ca[20],式中 Ca 表示外界 CO2浓度。
1.3 数据处理
试验数据采用Microsoft Excel 2003软件进行处理,DPS 7.5软件进行统计分析和处理间差异显著性检验(Duncan新复极差法)。
2 结果与分析
2.1 硅对番茄生长的影响
表1显示,营养液中添加适宜浓度的硅,可显著促进番茄的生长,但硅水平过高,则效果不明显。如T1、T2处理的株高分别比 CK增加了 9.51%、15.88%,茎粗分别增加了3.95%、11.18%,但T3处理的株高、茎粗与CK均无显著差异。虽然T1、T2、T3处理的根系干质量分别比 CK增加了13.12%、23.77%、16.49%,但T3处理的茎、叶干质量与CK均无显著差异,而T1、T2处理的茎、叶干质量则分别比CK提高了16.47%、14.76%和25.58%、29.47%。

表1 不同硅水平对番茄生长的影响Table 1 Effects of different silicon levels on the growth of tomato plants
2.2 硅对番茄不同器官硅含量的影响
从图1可以看出,番茄根、茎、叶中硅(SiO2)含量随营养液中硅浓度的提高而增加,除T2和T3处理的根系硅含量差异不显著外,其它处理各器官硅含量均存在显著差异,如CK番茄叶片硅含量仅0.56 mg/g,DW,T1、T2、T3处理叶片硅含量分别比CK高250.90%、403.59%、552.69%。不同处理番茄植株硅含量均为叶>根>茎,可能与不同器官的功能有关,根是吸收养分和向地上部运输的器官,茎是运输器官,而叶为营养合成器官,故大量的硅向叶集中。CK处理的番茄植株含少量硅,可能与幼苗3叶前在基质中生长吸收的部分硅在新生器官再分配有关。
2.3 硅对番茄叶片色素含量的影响
由表2可知,适当提高营养液中硅浓度,可提高番茄叶片光合色素含量,但硅水平过高,则会降低光合色素含量,如T1、T2处理的叶片叶绿素含量分别较CK增加17.73%和18.42%,但T3处理则比CK降低了5.76%,类胡萝卜素含量也有相似的变化趋势,这与硅浓度对番茄生长的影响相似。各处理叶片叶绿素与类胡萝卜素含量的比值则无显著差异。
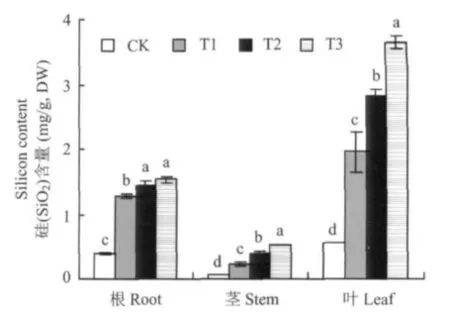
图1 不同硅水平对番茄各器官硅含量的影响Fig.1 Effects of different silicon levels on silicon contents in different organs of tomato

表2 不同硅水平对番茄叶片光合色素的影响Table 2 Effects of different silicon levels on photosynthetic pigments of tomato leaves
2.4 硅对番茄叶片光合作用特性的影响
本试验条件下(图2),不同处理番茄叶片的净光合速率(Pn)的日变化规律相似,均为不对称的双峰曲线(图3A),但处理间Pn除在7:00差异不显著外,其它时间均以T2处理较高,T1次之,CK和T3较低,且尤以13:00时差异最为显著。此时,T3处理的Pn仅为CO26.11 μmol/(m2·s),与 CK 无显著差异,而T2、T1处理分别比CK高49.84%和32.44%。不同处理番茄叶片气孔导度(Gs)的日变化动态与Pn不同,但仍以T2处理较高,T3较低,T1、CK居中(图3B),11:00时 Gs达峰值时,T1、T2处理分别比CK高6.98%和11.63%,而T3则比CK低3.49%。由于午间各处理番茄叶片的胞间CO2浓度(Ci)均降低(图3C),而气孔限制值(Ls)均升高(图3D),表明13:00时Pn的降低主要以气孔限制为主。

图2 试验环境因子日变化动态Fig.2 Diurnal changes of environmental factors related to the experiment
2.5 硅对番茄叶片蒸腾速率和瞬时水分利用效率的影响
图4显示,各处理番茄叶片蒸腾速率(Tr)的日变化均呈单峰曲线,但11:00前除T3处理的Tr较低外,T1、T2与CK均无显著差异,11:00后处理间的差异渐趋增加,13:00 Tr达峰值时,Tl、T2、T3处理分别比CK降低了7.42%、11.47%和23.08%。番茄叶片瞬时水分利用效率(WUEi)以T2较高,CK较低,T1、T3居中,如在11:00 WUEi达峰值时,T1、T2、T3处理分别比CK高22.22%、35.47%和17.52%,但一天内T1处理的WUEi除中午前后略高于T3外,早、晚则与T3无显著差异。
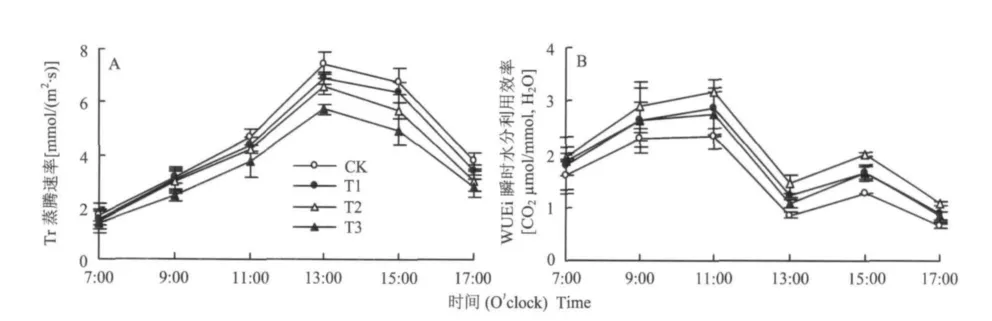
图4 不同硅水平对番茄叶片蒸腾速率及瞬时水分利用效率日变化的影响Fig.4 Effects of different silicon levels on diurnal changes of Tr and WUEi in tomato leaves
3 讨论
3.1 番茄对硅的吸收利用特性
高等植物对硅的吸收分为主动吸收、被动吸收和排斥吸收三种类型[5],但 Nikolic 等[21]通过68Ge示踪研究发现,番茄对硅的吸收属被动排斥吸收机制,亦即从外部溶液到根部皮层的被动扩散和通过运载体介导的从皮层细胞到质外体的主动排斥共存。Mitani[22]等研究证实,番茄施硅培养 8 h 后,其伤流液中的硅仍低于体外营养液硅水平,而培养至37 h,则其伤流液中的硅可接近体外培养液水平,表明番茄对硅的吸收能力较弱,但随着培养时间的延长,番茄植株体内可积累一定量的硅[5]。梁永超等[16]研究也证实,50 mg/L和100 mg/L硅水平培养的番茄植株,其茎的硅含量分别比对照增加145%和200%,叶片硅含量分别比对照增加60%和130%,且施硅处理的番茄植株生长量显著高于对照。本试验也得出了相似的结果,即番茄根、茎、叶中硅含量均随营养液硅水平的提高而增加,且以叶片硅含量最高,根系次之,茎最低(图1),同时适宜的硅水平显著促进了番茄植株的生长(表1)。
3.2 硅对番茄光合作用和蒸腾作用的影响机理
研究表明,硅可显著提高水稻[9]、甜瓜[13]叶片的叶绿素含量,对棉花[23]、大豆[15]、生姜[24]等叶片叶绿素含量无显著影响,但几乎所有的研究表明,适量施硅可显著提高作物的光合速率[10,25]。本试验结果表明,适宜的硅水平可显著增加番茄叶片叶绿素和类胡萝卜素的含量,提高光合速率,而硅水平过高,则叶片色素含量下降,光合速率也随之降低(表2,图3)。硅增强番茄光合作用的原因可能与适量硅水平提高了叶片的叶绿素含量有关,尤其较高的叶绿素b含量有利于LHCⅡ复合体的组装及稳定,而LHCⅡ在类囊体膜中除能进行光能的吸收和传递外,还可维持类囊体膜的结构、调节激发能在2个光系统之间的分配[26];其次,适量施硅,番茄叶片较高的类胡萝卜素含量可增强光合电子传递量子效率和抵御午间强光的能力,从而减轻光抑制程度[27];第三,适量施硅可抑制叶肉细胞光合活性的降低[14,28],因为午间光合午休时,适量施硅番茄叶片Gs较高,而其Ci较低(图3)。
本试验结果还表明,施硅显著降低了番茄叶片的蒸腾速率,且随着营养液硅浓度的升高,降幅增大(图 4),这与前人在黄瓜[12]、甜瓜[13]等作物上的研究结果相似。Kim等[29]的研究表明,水稻叶片水分散失的主要途径包括气孔蒸腾和角质层蒸腾,而硅可促进角质-双硅层结构的形成,抑制水分的角质层蒸腾。Gao[30]等研究结果证实,硅也可降低玉米叶片的气孔蒸腾,而不仅仅是叶片角质层蒸腾。本研究中,适宜硅浓度处理的番茄叶片气孔导度显著升高,可见硅不是通过促进气孔关闭来降低蒸腾作用。邹春琴[11]等研究发现,施硅处理的向日葵木质部汁液流速较不施硅降低了23%,而1 g直径为7 nm的硅胶颗粒吸附表面积高达400 m2[31],植物体内即使只沉积了少量的硅胶颗粒,其对水分的吸附作用也相当可观。结合本试验中施硅处理番茄植株硅含量显著增加的现象,推测硅在木质部导管壁的大量沉积,增强了木质部导管的亲水性,从而降低了番茄植株的蒸腾作用。此外,硅还可通过影响水稻叶片气孔大小和密度,降低蒸腾速率[32]。而关于硅对番茄叶片气孔特征的影响,尚需进一步研究。
4 结论
1)适量施硅可显著促进番茄植株的生长,增加株高、茎粗及根、茎、叶生长量,以1.2 mmol/L硅浓度处理效果较好,如进一步提高硅浓度至1.8 mmol/L,则会抑制番茄生长。
2)施硅可显著提高番茄植株各器官的硅含量,且随营养液中硅浓度的提高而显著增加茎、叶中的硅含量,但较高的硅浓度并未显著提高根系的硅含量。
3)适当提高硅浓度可显著增加番茄叶片的光合色素含量,提高光合速率,以1.2 mmol/L硅浓度处理较佳,而当硅浓度达1.8 mmol/L时,番茄叶片的光合色素含量及光合速率反而降低。
4)随硅浓度的增加,番茄叶片的蒸腾速率显著降低;同时,施硅亦显著提高了番茄叶片的瞬时水分利用效率,其中以硅浓度为1.2 mmol/L的瞬时水分利用效率较高。
[1] 梁永超,张永春,马同生.植物的硅素营养[J].土壤学进展,1993,21(3):7-14.Liang Y C,Zhang Y C,Ma T S.Silicon nutrition of plant[J].Prog.Soil Sci.,1993,21(3):7-14.
[2] Ma J F,Miyake Y,Takahashi E.Silicon as a beneficial element for crop plants[J].Silicon Agric.,2001,8:17-39.
[3] Arnon D I,Stout P R.The essentiality of certain elements in minute quantity for plants with special reference to copper[J].Plant Physiol.,1939,14(2):371-375.
[4] Liang Y C,Sun W C,Zhu Y G,Christie P.Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants:A review[J].Environ.Poll.,2007,147(2):422-428.
[5] Takahashi E,Ma J F,Miyake Y.The possibility of silicon as an essential element for higher plants[J].Comm.Agric.Food Chem.,1990,2(2):99-102.
[6] 薛高峰,孙万春,宋阿琳,等.硅对水稻生长、白叶枯病抗性及病程相关蛋白活性的影响[J].中国农业科学,2010,43(4):690-697.Xue G F,Sun W C,Song A L et al.Influence of silicon on rice growth,resistance to bacterial blight and activity of pathogenesisrelated proteins[J].Sci.Agric.Sin.,2010,43(4):690-697.
[7] 黄海荣,Bokhtiar S M,徐林,等.硅肥对甘蔗光合特性和产量的影响[J].广西农业科学,2009,40(12):1564-1569.Huang H R,Bokhtiar S M,Xu L et al.Effect of silicon fertilization on yield and photosynthetic attributes in sugarcane(Saccharum officinarum L.hybrid)[J].Guangxi Agric.Sci.,2009,40(12):1564-1569.
[8] Ma J F,Yamaji N.Functions and transport of silicon in plants[J].Cell.Mol.Life Sci.,2008,65(19):3049-3057.
[9] 高臣,刘俊渤,常海波,等.硅对水稻叶片光合特性和超微结构的影响[J].吉林农业大学学报,2011,33(1):1-4.Gao C,Liu J B,Chang H B et al.Effects of silicon on rice leaf photosynthesis and ultrastructure[J].J.Jilin Agric.Univ.,2011,33(1):1-4 .
[10] 丁燕芳,梁永超,朱佳,李兆君.硅对干旱胁迫下小麦幼苗生长及光合参数的影响[J].植物营养与肥料学报,2007,13(3):471-478.Ding Y F,Liang Y C,Zhu J,Li Z J.Effects of silicon on plant growth,photosynthetic parameters and soluble sugar content in leaves of wheat under drought stress[J].Plant Nutr.Fert.Sci.,2007,13(3):471-478 .
[11] 邹春琴,高霄鹏,刘颖杰,等.硅对向日葵水分利用效率的影响[J].植物营养与肥料学报,2005,11(4):547-550.Zou C Q,Gao X P,Liu Y J et al.Effects of silicon on water use efficiency in sunflower with solution culture[J].Plant Nutr.Fert.Sci.,2005,11(4):547-550.
[12] 周秀杰,赵红波,马成仓,李清芳.外源硅对黄瓜叶片组织结构和保水能力的影响[J].生态学杂志,2009,28(3):556-559.Zhou X J,Zhao H B,Ma C C,Li Q F.Effects of exogenous silicon on leaf structure and water-holding capacity of cucumber plant[J].Chin.J.Ecol.,2009,28(3):556-559.
[13] 卢钢,曹家树.硅对甜瓜早熟性及光合特性的影响[J].园艺学报,2001,28(5):421-424.Lu G,Cao J S.Effects of silicon on earliness and photosynthetic characteristics of melon[J].Chin.J.Hortic.Sci.,2001,28(5):421-424.
[14] 宋阿琳,李萍,李兆君,梁永超.硅对镉胁迫下白菜光合作用及相关生理特性的影响[J].园艺学报,2011,38(9):1675-1684.Song A L,Li P,Li Z J,Liang Y C.Effect of silicon on photosynthesis and its related physiological parameters of Brassica campestris L.ssp.chinensis(L.)Makino grown under Cd stress[J].Chin.J.Hortic.Sci.,2011,38(9):1675-1684 .
[15] 李清芳,马成仓,李韩平,等.土壤有效硅对大豆生长发育和生理功能的影响[J].应用生态学报,2004,15(1):73-76.Li Q F,Ma C C,Li H P et al.Effects of soil available silicon on growth,development and physiological functions of soybean[J].Chin J.Appl.Ecol.,2004,15(1):73-76.
[16] 梁永超,陈兴华,马同生,等.硅对番茄生长、产量与品质的影响[J].江苏农业科学,1993,4:48-50 Liang Y C,Chen X H,Ma T S et al.Effect of silicon on growth,yield and quality of tomato[J].Jiangsu Agric.Sci.,1993,4:48-50.
[17] Stamatakis A,Papadantonakis N,Savvas D et al.Effects of silicon and salinity on fruit yield and quality of tomato grown hydroponically[J].Acta Hortic.(ISHS),2003,609:141-147.
[18] Van der Vorm P D J.Dry ashing of plant material and dissolution of the ash in HF for the colorimatric determination of silicon[J].Commun.Soil Sci.Plant Anal.,1987,18(11):1181-1189.
[19] 赵世杰,刘华山,董新纯.植物生理学实验指导[M].北京:中国农业科学技术出版社,1998.Zhao S J,Liu H S,Dong X C.Techniques of plant physiological experiment[M].Beijing:Chinese Agricultural Science and Technology Press,1998.
[20] Farquhar G D, SharkeyT D. Stomatalconductanceand photosynthesis[J].Annu.Rev.Plant Physiol.,1982,33:317-345.
[21] Nikolic M,Nikolic N,Liang Y C et al.Germanium-68 as an adequate tracer for silicon transport in plants.characterization of silicon uptake in different crop species 1[J].Plant Physiol.,2007,143(1):495-503.
[22] Mitani N,Ma J F.Uptake system of silicon in different plant species[J].J.Exp.Bot.,2005,56(414):1255-1261.
[23] 李清芳,马成仓.土壤有效硅对棉花幼苗营养代谢的影响[J].中国农业科学,2003,36(6):726-730.Li Q F,Ma C C.Effect of available silicon in soil on nutritive metabolism of cotton seedling[J].Sci.Agric.Sin.,2003,36(6):726-730 .
[24] 张国芹,徐坤,王兴翠,等.硅对生姜叶片水、二氧化碳交换特性的影响[J].应用生态学报,2008,19(8):1702-1707.Zhang G Q,Xu K,Wang X C et al.Effects of silicon on exchange characteristics of H2O and CO2in ginger leaves[J].Chin J.Appl.Ecol.,2008,19(8):1702-1707.
[25] 朱佳,梁永超,丁燕芳,李兆君.硅对低温胁迫下冬小麦幼苗光合作用及相关生理特性的影响[J].中国农业科学,2006,39(9):1780-1788.Zhu J,Liang Y C,Ding Y F,Li Z J.Effect of silicon on photosynthesis and its related physiological parameters in two winter wheat cultivars under cold stress[J].Sci.Agric.Sin.,2006,39(9):1780-1788.
[26] 郭春爱,刘芳,许晓明.叶绿素b缺失与植物的光合作用[J].植物生理学通讯,2006,42(5):967-972.Guo C A,Liu F,Xu X M.Chlorophyll-b deficient and photosynthesis in plants[J].Plant Physiol.Commun.,2006,42(5):967-972.
[27] Demming-A dams B,Adams W W.Photoprotection and other responses of plant to high light stress[J].Annu.Rev.Plant Physiol.Plant Mol.Biol.,1992,43:599-626.
[28] 谢寅峰,杨万红,陆美蓉,等.模拟酸雨胁迫下硅对髯毛箬竹光合特性的影响[J].应用生态学报,2008,19(6):1179-1184.Xie Y F.Yang W H,Lu M R et al.Effects of silicon on photosynthetic characteristics of Indocalamus barbatus under simulated acid rain stress[J].Chin J.Appl.Ecol.,2008,19(6):1179-1184.
[29] Kim S G,Kim K W,Park E W,Choi D.Silicon induced cell wall fortification of rice leaves:a possible cellular mechanism of enhanced host resistance to blast[J].Phytopathology,2002,92(10):1095-1103.
[30] Gao X P,Zou C Q,Wang L J,Zhang F S.Silicon improves water use efficiency in maize plants[J].J.Plant Nutr.,2004,27(8):1457-1470.
[31] Wang J, Naser N. Improved performance of carbon paste ampermeric biosensors through the incorporation of fumed silica[J].Electroanalysis,1994,6(7):571-575.
[32] Kang Y S.Effects of silicon stomatal size and frequency in rice plants[J].Korean J.Crop Sci.,1991,36(1):70-73.
