科尔沁沙地家榆林的种子散布及幼苗更新
2013-09-11杨允菲白云鹏李建东
杨允菲,白云鹏,李建东
(东北师范大学草地科学研究所植被生态科学教育部重点实验室,长春 130024)
植物的种群更新和生态位空间扩展依赖于繁殖体的传播,植物的繁殖体具有多样性,除了有性生殖的种子,还有营养繁殖的构件[1],虽然营养繁殖的构件更有利于种群更新,但对生态位空间扩展的可动性是非常有限的,其种群更大的空间扩展仍依赖于种子散布[2-3]。在漫长的趋同适应和进化过程中,植物形成了借助各种传播因子的种子散布类型[1]。其中,风播植物是易于远距离传播的重要类型之一,容易将种子传播到其他群落。虽然风播植物受风向、风速和立地条件影响,其种子散布过程较为复杂[4-5],但从种子源向外扩散往往具有某些规律性格局[6-8],便于定量描述和预测群落动态[9-12],甚至开展种群生活史策略等进化生态学机理的深入分析[13-14]。所以,有关风播植物种子散布的研究迄今已有大量报道[15-22]。
家榆(Ulums pumila)是科尔沁沙地原生森林植被类型最具有代表性的建群种[23],果实具宽翅,是典型的风播植物。历史上广泛形成大面积单一优势种家榆林,但目前很少连片分布,只在沙丘连绵、地形起伏多样的局域地段保留原植被景观,大部分被农田切割为岛屿状分布,或完全被人工杨树林所替代。尽管如此,这些残余家榆林对于维护立地乃至沙地区域生态平衡及生物多样性等仍在发挥着重要的作用。
本研究以科尔沁沙地具有代表性的原生家榆林为对象,通过对林缘和孤立株不同方向从基部向外的有序取样与林下不同生境的随机取样调查,分析家榆林内和不同方向林缘的种子沉降特征;林缘和孤立树的种子散布格局,以及林下不同生境和林缘不同距离的幼苗分布特征等,揭示家榆林的维持与更新现状,为家榆林保护提供科学参考。
1 研究地点和方法
1.1 研究区的自然概况
研究区位于吉林省西部的前郭尔罗斯蒙古族自治县查干花种畜场三场(44°36.385'N,124°06.521'E)。该区属北温带大陆性季风气候,年平均气温4.5℃,1月份极端最低气温-36.1℃,7月份极端最高气温37℃,无霜期135—140 d,年降雨量400—500 mm之间,多集中在7、8月份,约占全年降水量的2/3。春季多风,春夏秋三季以西南风为主,冬季则多刮西北风,5月份平均风速为4.1 m/s[24]。研究样地为起伏较大而不宜开垦的固定沙丘群,面积在500 hm2左右,是现时保存较好的榆树疏林“岛屿”,具有原生外貌景观,最大树龄在60a以上。家榆成株树干弯曲,中上部多分枝,不同龄株高差异不大,为6—8 m,但主径和树冠幅有较大差异,在沙丘的阴坡和阳坡家榆主径较粗,树冠较大;而生长在丘顶和丘底的植株相对主径较细,树冠较小。土壤类型为风沙土。群落中木本层种类构成简单,除建群种家榆外,仅伴生少量黄榆(Ulmus macrocarpa)、桑(Morus alba)、山杏(Amygdalus sibirica)。层间植物有草白蔹(Ampelopsis aconitifolia)。草本层以狗尾草(Setaria viridis)、马唐(Digitaria sanguinalis)和绿珠藜(Chenopodium acuminatum)、黄蒿(Artemisia scoparia)等一年生、二年生植物为优势类群,伴生糙隐子草(Celeistogenes squarrosa)、燕麦芨芨草(Achenatherum avinoides)、羊草(Leymus chinensis)等多年生植物[23]。林缘开阔地多为放牧场,面积大的多开垦为农田。
1.2 种子散布取样
2009年5月30日,在家榆种子全部脱落后分别做如下取样:
1)林下和林窗的随机取样。按沙丘部位,林下设置4个小生境的样地:阳坡、阴坡、丘顶、丘底,各样地的家榆生长较为均匀一致,郁闭度为0.7以上,株距1.4—3.2 m,地表覆盖枯叶和1年生草本枯株;林窗设置2个小生境样地,丘顶为草层盖度在30%左右的黄蒿群落,面积为20 m×40 m,丘底为草层盖度在80%左右的糙隐子草+羊草群落,面积为20 m×50 m。每个样地取样均10次重复。
2)林缘和孤立树的顺序取样。用GPS仪定位方向。在不少于80 m开阔且地表相对平坦、草层盖度在60%—80%左右的林缘草地作为样地。由于这样的地段多数被开垦为农田,所以林缘仅东南和西南2个方向符合取样标准,每个方向3个纵断面重复。为了真实反映风向对家榆种子散布的影响,结合当地的风向特点,以半径不少于50 m且具有代表性和地表平坦的结实成株孤立树(树高8 m左右)作为样本,3株重复,按顺(东北)、逆(西南)风向及其他方向的取样。同林缘一样,由于局部地形的原因或被开垦,在其他方向中仅有西北、东南2个方向符合取样标准,全体顺序取样均从树基部开始,用米尺向外拉一条样线,每1 m设一个样方做各纵断面的单位面积取样,至不再出现家榆的翅果为止。全部调查样方的面积均为20 cm×20 cm。计数样方内完整而没有霉变,即当年落下的白色或淡黄色的家榆翅果(种子)数量。
1.3 幼苗更新取样
2009年8月15日,家榆幼苗已经生长为15—20 cm,个别已经出现分枝。在沙丘的阳坡和阴坡、沙丘顶上和底下的林下,以及沙丘顶上和底下的林窗,即种子沉降调查的6个生境,分别做家榆实生苗数量的随机调查,10个样方重复;在林缘,先用米尺拉出一个1 m宽,20 m长的样线断面,再从树基部开始向外每1 m作为一个调查样方,顺序记录家榆实生苗数量,3个断面重复。全部调查样方的面积均为1 m×1 m。
1.4 数据分析
分别将取样面积的种子数量换算为常规的1 m2单位面积的数量。统计林下和林窗种子和实生苗数量、林缘和孤立树不同方向断面累积单位面积种子数量,并做其生境或不同方向的多重比较。林缘和孤立树各方向单位面积种子数量先计算出重复取样的平均值后,再分别计算其相对频率:
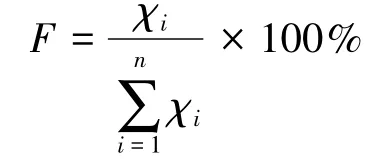
式中,F为i距离种子的频度;x每单位面积的种子数;∑x为各距离单位面积种子数之和;i=1,2,3,…,n。
根据统计分布原理[25],拟采用2种理论分布的密度函数形式来反映不同方向断面依次远离种子源单位面积的种子散布频度的格局。
(1)韦伯分布
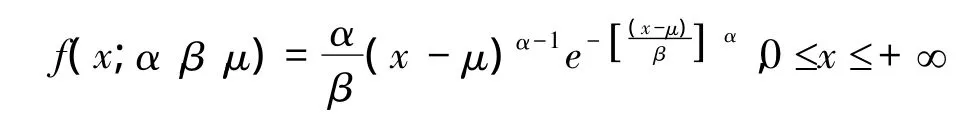
式中,α>0为形状参数;β>0为尺度参数;μ≥0为位置参数。
(2)对数-正态分布
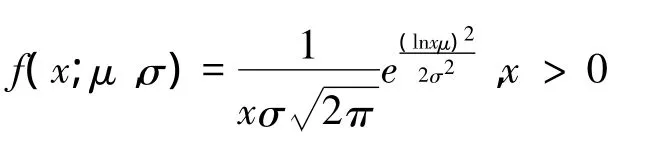
式中,μ和σ分别为自然对数的平均数和标准差。
对林缘和孤立树各方向断面远离种子源单位面积种子散布的观测值和由韦伯分布理论频度还原的期望值逐一做相关性分析。利用Microsoft Excel 2003和SPSS统计软件(Version12.0,SPSS Inc.,Chicago,IL,USA)处理与分析数据
2 结果与分析
2.1 林下和林窗的种子沉降特征
家榆种子沉降的密度,林下以阴坡和阳坡明显大于丘顶和丘底,林窗又以丘顶明显大于丘底(图1)。其中,林下的丘底又明显大于和丘顶。竖线的标准误也可以在一定程度上反映生境内种子分布的均匀性,从图1还可以看出,在林下,阴坡和阳坡种子沉降的数量较丘顶和丘底的变异性大,在林窗丘顶较丘底的变异性大,反映了种子密度越大越不均匀的种子沉降特征。
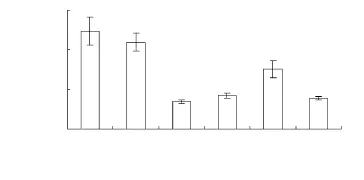
图1 不同生境家榆的种子密度(M±SE,不同字母在P≤0.05水平上差异显著)Fig.1 The densities of seeds of Ulums pumila in different hatitats(Means±standard error,the different letter means significant difference at P ≤0.05;U1:understory in the shade slope sand dune,U2:understory in the sunny slope sand dune,U3:understory at the top of sand dune,U4:understory on the bottom of sand dune,G1:canopy gap at the top of sand dune,G2:canopy gap on the bottom of sand dune,the same below)
2.2 林缘和孤立树不同方向的种子沉降特征
家榆向周围不同方向种子连续散布的距离,林缘2个方向和孤立树4个方向(图2)均以当地多逆风频率的西南方向最近,分别是15 m和13 m,并且累积数量均最少,分别为(1383.3±147.4)粒和(683.3±122.8)粒;孤立树以多顺风频率的东北方向最远,为25 m,并且累积数量也最多,为(6191.7±733.2)粒,孤立树的东南方向也显著多于西南方向,达4.2倍。由此反映了家榆不同方向种子沉降的不均匀性特征。
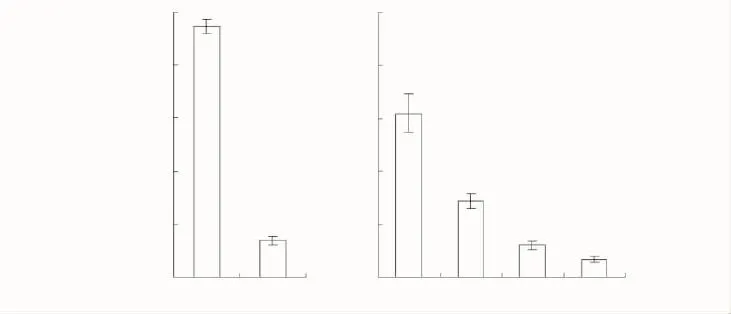
图2 林缘和孤立树不同方向纵断面单位面积累积的家榆种子数量Fig.2 The total cumulative seeds of Ulums pumila per unit area away from seed source in woodland edges and isolated tree along vertical sections of different directions
2.3 林缘和孤立树不同方向种子散布格局
经韦伯分布和对数-正态分布2种理论分布密度函数参数拟合的适合性检验,无论林缘,还是孤立树的各个方向顺序远离种子源散布到各单位面积种子数量频度均达到了极适合水平(<0.500)。其中,又均以韦伯分布密度函数拟合的普遍更好(表1)。各方向纵断面单位面积种子散布的平均观测值频率及其韦伯分布拟合曲线见图3,各方向密度平均观测值和由韦伯分布理论频度还原的期望值及其极显著(P<0.01)的相关性直线见图4。由此反映了尽管家榆不同方向种子沉降的数量有较大差异,但仍具有相同的种子散布格局。
2.4 不同生境和林缘的幼苗数量特征
经统计比较,家榆幼苗的密度,林下以阴坡明显大于阳坡、丘顶明显大于丘底,林窗也是丘顶明显大于丘底(图5);林缘3个样带在19 m内实生苗密度的平均值呈连续性分布,在15 m处出现最大值(12.0±5.7)株/m2,既没有规律性变化,不同间距实生苗(0.3±0.3)—(12.0±5.7)株/m2的差异大多数也均未达到显著水平(P>0.05)(图5);此外,从其标准误线也可看出,各生境及林缘不同距离的变异性均较大,反映了家榆幼苗分布的不均匀性特征。
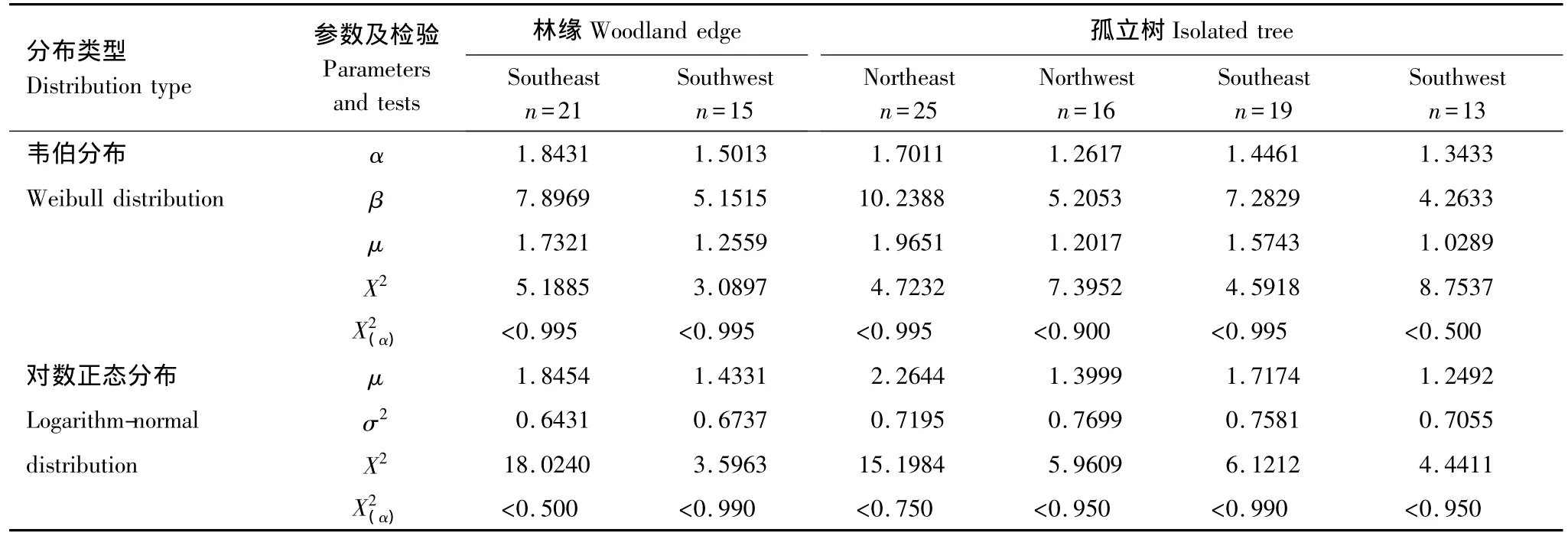
表1 林缘和孤立树不同方向纵断面家榆种子散布频度的两种分布参数与X2检验Table 1 Parameters and X2tests of two distribution types on seed dispersal frequency of Ulmus pumila away from tree bases in woodland edges and isolated trees along vertical sections of different directions
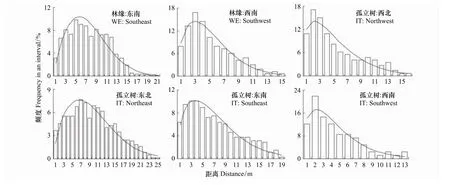
图3 林缘和孤立树不同方向纵断面单位面积家榆种子散布的平均观测值频率及其韦伯分布拟合曲线Fig.3 The seed dispersal frequencies of observed average values(column)of Ulums pumila per unit area and the expected curves of Weibull distribution away from seed source in woodland edges(WE)and isolated trees(IT)along vertical sections of different directions
3 结论与讨论
家榆种子沉降的空间变异性在生境间和生境内将分别具有不同的作用因素。其中,不同生境间种子沉降数量的差异主要与种子源量的差异有关。一般是植株越大,结实越多,样地内沙丘的阴坡和阳坡小生境的家榆植株普遍比丘顶和丘底小生境的大,致使种子密度出现林下以阴坡和阳坡明显大于丘顶和丘底的空间变异。而同一小生境种子分布的均匀性则与种子再传播有关。家榆翅果具典型的风播表型,沉降于地表后,受大地形、微地形,以及地表的覆盖或裸露程度等不同影响,一些种子还将二次或多次传播[3-4]。沙丘坡的微地形较丘顶和丘底更复杂多样,种子的可动性也更大;盖度小的丘顶林窗要比盖度大的丘底林窗种子的可动性更大,致使这些小生境内种子分布更不均匀。
林缘2个方向在15—21 m内、孤立树4个方向在13—25 m内,家榆的种子呈连续分布,不同方向的散布距离及其断面累积单位面积的种子数量均存在不同程度的差异。这主要是种子脱落时的风向和风速的差异所致,因为风速的大小将会影响种子传播距离,风向将会影响不同方向种子沉降的数量,通常顺风向频率大的方向将大于逆风向的方向[13-14,26]。家榆孤立树以东北方向最多、最远,林缘和孤立树均以西南方向最少、最近。这恰好与该地区夏初多刮西南风[24],东北属于多顺风频率的方向,西南则为多逆风频率的方向相一致。
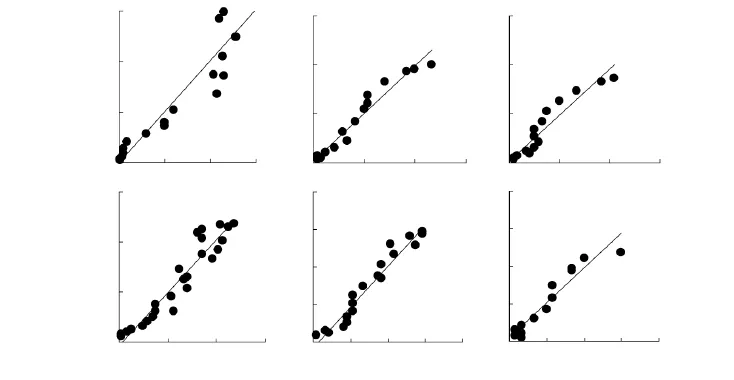
图4 林缘和孤立树各方向纵断面家榆种子密度观测值与韦伯分布理论频度还原的期望值的相关性Fig.4 Correlation on between observed values of seed densities of Ulums pumila and the restored expected values through theoretical frequency of Weibull distribution in woodland edges(WE)and isolated trees(IT)along vertical sections of different directions
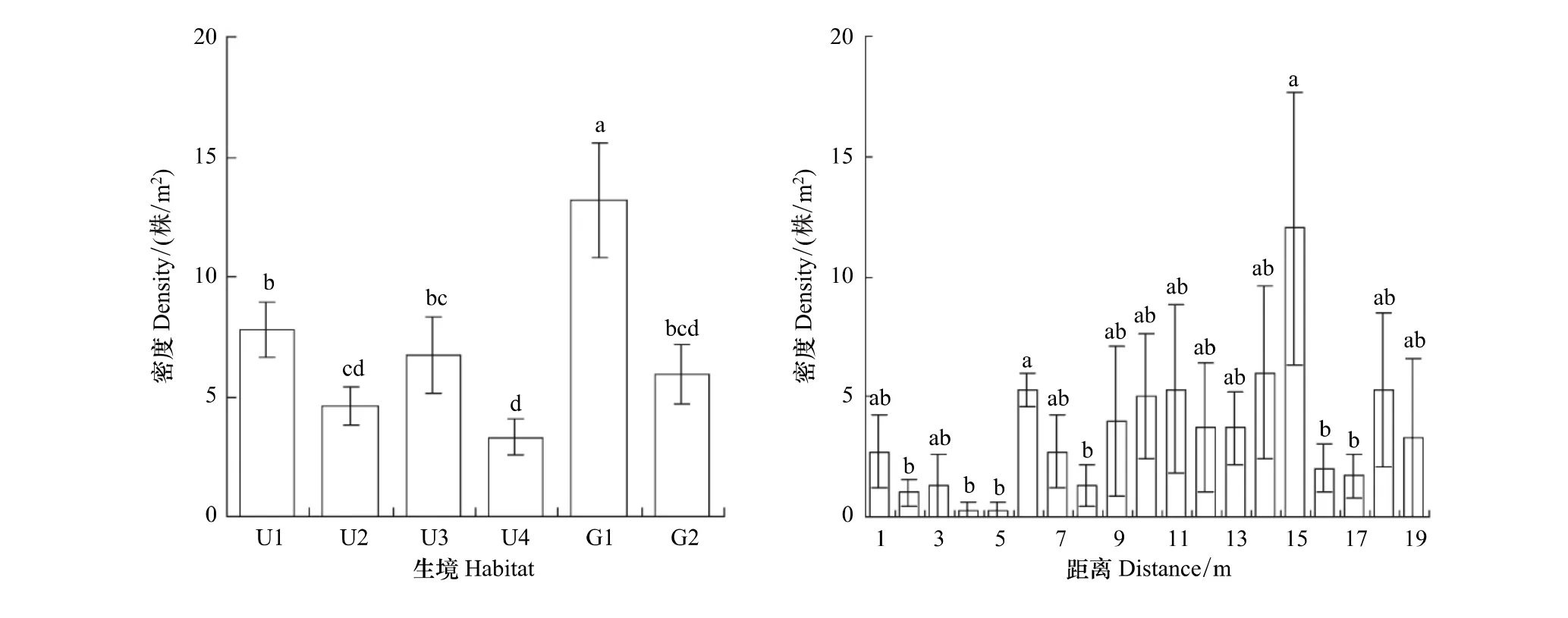
图5 不同生境和林缘不同距离家榆幼苗密度Fig.5 The seedling densities of Ulums pumila in the different habitats and in different distance of woodland edges
在家榆林缘和孤立树的6个方向上,顺序远离家榆种子源散布到各单位面积种子数量的频度同时适合于韦伯分布和对数-正态分布,意味着尽管受风向和风速的影响,家榆向不同方向散布的种子数量和距离和数量均存在或大或小的差异,但散布格局却是一致的。在种子散布格局的定量刻画模型中,韦伯分布的密度函数拟合的形状参数α的大小范围可以很好地解释种子散布机制[14,26]。即将α<1时视为“近距离”种子散布,α>1时视为“远距离”种子散布,α=1时视为“随机距离”种子散布。林缘和孤立树6个方向家榆种子散布的韦伯分布参数同时出现α>1,表明在科尔沁沙地自然条件下,无论是林缘还是孤立树,无论是顺风向还是逆风向,家榆均具有稳定的“远距离”种子散布机制。
在种子散布两个半月以后,研究样地家榆实生树苗在林下和林窗6个生境内均有大量分布,在林缘19 m内的每1 m的间距中呈连续性分布,意味着家榆林不仅具有较强的幼苗更新力,也具有较大的空间扩展力。但是,目前在林缘很少见到不同龄级的幼株,这可能与频繁放牧的人为扰动有关。仅就家榆同生群幼苗在较大范围内呈连续性较高密度分布的现象,不难解释在科尔沁沙地天然条件下目前仍具备着形成单一优势种家榆林的生物学基础。由此也可以推测:如果不加限制和人为扰动,在历史上曾形成大片家榆林的科尔沁沙地,现存的家榆林仍可以很好地进行自然更新,仍可以在适宜的沙地生境逐渐扩大其分布空间。
经统计分析,无论林下还是林缘,种子沉降的数量与幼苗数量之间并没有相关性,这可能与幼苗调查时间有关。因为植物在苗期抗逆性弱,家榆为短命种子,在适宜的条件下,落地的种子很快就萌发,幼苗却极少能存活下来。有关家榆幼苗在干旱春季自然萌发与存活状况有待深入研究。
[1] Drake D R.Relationships among the seed rain,seed bank and vegetation of a Hawaiian forest.Journal of Vegetation Science,1998,9(1):103-112.
[2] Sagnarda F,Pichota C,Dreyfusa P,Jordano P,Fady B.Modelling seed dispersal to predict seedling recruitment:recolonization dynamics in a plantation forest.Ecological Modelling,2007,203(3/4):464-474.
[3] Howe H F,Smallwood J.Ecology of seed dispersal.Annual Review of Ecology and Systematics,1982,13:201-228.
[4] Nathan R,Safriel U N,Noy-Meir I,Schiller G.Spatiotemporal variation in seed dispersal and recruitment near and far from Pinus halepensis Trees.Ecology,2000,81(8):2156-2169.
[5] Xiao Z S,Zhang Z B,Wang Y S.Effects of seed size on dispersal distance in five rodent-dispersed fagaceous species.Acta Oecologica,2005,28(3):221-229.
[6] Fernando B,Salvador T.Temporal and spatial patterns of seed dispersal in two Cistus species(Cistaceae).Annuals of Botany,2002,89(4):427-434.
[7] Zhang Y B,Li J W,Zhang H,Zou D L,Wu F P,Cheng C L,Li J Q,Li S Y.Spatiotemporal patterns of seed dispersal in Populus euphratica.Acta Ecologica Sinica,2005,25(8):1994-2000.
[8] Han Y Z,Wang Z Q.Spatial pattern of Manchurian ash seed dispersal in secondary hardwood forests.Acta Phytoecologica Sinica,2002,26(1):51-57.
[9] Peart D R.Species interactions in a successional grassland.I.Seed rain and seedlings recruitment.Journal of Ecology,1989,77:236-251.
[10] Moles A T,Drake D R.Potential contribution of the seed rain and seed bank to regeneration of native forest under plantation pine in New Zealand.New Zealand Journal of Botany,1999,37(1):83-93.
[11] Godoy J A,Jordano P.Seed dispersal by animals:exact identification of source trees with endocarp DNA microsatel lites.Molecular Ecology,2001,10(9):2275-2283.
[12] Zou L,Xie Z Q,Li Q M,Zhao C M,Li C L.Spatial and temporal pattern of seed rain of Abies fargesii in Shennongjia Nature Reserve,Hubei.Biodiversity Science,2007,15(5):500-509.
[13] Yang Y F.The study on seed dispersal of Puccinellia tenuiflora on alkalization meadow in the Songnen Plain of China.Acta Ecologica Sinica,1990,10(3):288-290.
[14] Yang Y F,Zhu L.Pattern of seed dispersal of Hordeum brevisubulatum of alkalizied meadow in Songnen Plain of China.Acta Botanica Sinica,1994,36(8):636-644.
[15] Dow B D,Ashley M V.Microsatellite analysis of seed dispersal and parentage of samplings in bur oak,Quercus macrocarpa.Molecular Ecology,1996,5(5):615-627.
[16] He T H,Krauss S L,Lamont B B,Miller B P,Enright N J.Long-distance seed dispersal in a metapopulation of Banksia hookeriana inferred from a population allocation analysis of amplified fragment length polymorphism data.Molecular Ecology,2004,13(5):1099-1109.
[17] Nathan R.Long-distance dispersal of plants.Science,2006,313(5788):786-788.
[18] Schnabel A,Nason J D,Hamrick J L.Understanding the population genetic structure of Gleditsia triacanthos L.:seed dispersal and variation in female reproductive success.Molecular Ecology,1998,7(7):819-832.
[19] Michael E.Seed Ecology.London and New York:Chapman and Hall,1985:57-116.
[20] Liu Z G,Zhu J J,Yuan X L,Wang H X,Tan H.On seed rain and soil seed bank of Larix olgensis in montane regions of eastern Liaoning Province,China.Acta Ecologica Sinica,2007,27(2):579-587.
[21] Liu J M,Zhong Z C.Nature of seed rain,the seed bank and regeneration of a Castanopsis fargesii community of Fanjing Mountain.Chinese Journal of Plant Ecology,2000,24(4):402-407.
[22] Yang Y F,Bai Y P,Li J D,Li L.Spatial patterns of the seed dispersal in Hemiptelea davidii woodland in Keerqin sandy land,China.Chinese Journal of Applied Ecology,2010,21(8):1967-1973.
[23] Li J D,Yang Y F.Structure of different types on components of plant species in Ulmus open forest in the Songnen Plains of China.Acta Agrestia Sinica,2003,11(4):277-282,300-300.
[24] http://www.weather.com.cn/cityintro/101060801.shtml
[25] Fang K T,Xu J L.Statistical Distribution.Beijing:Science Press,1987:283-298.
[26] Yang Y F,Zhu L.Analysis on the mechanism of seed dispersal of Puccinellia chinampoensis on alkalized meadow in Songnen Plain of China.Acta Botanica Sinica,1995,37(3):222-230.
参考文献:
[7] 张玉波,李景文,张昊,邹大林,武逢平,程春龙,李俊清,李帅英.胡杨种子散布的时空分布格局.生态学报,2005,25(8):1994-2000.
[8] 韩有志,王政权.天然次生林中水曲柳种子的扩散格局.植物生态学报,2002,26(1):51-57.
[12] 邹莉,谢宗强,李庆梅,赵常明,李传龙.神农架巴山冷杉种子雨的时空格局.生物多样性,2007,15(5):500-509.
[13] 杨允菲.松嫩平原碱化草甸星星草种子散布的研究.生态学报.1990,10(3):288-290.
[14] 杨允菲,祝玲.松嫩平原碱化草甸野大麦的种子散布格局.植物学报,1994,36(8):636-644.
[20] 刘足根,朱教君,袁小兰,王贺新,谭辉.辽东山区长白落叶松(Larix olgensis)种子雨和种子库.生态学报,2007,27(2):579-587.
[21] 刘济明,钟章成.梵净山栲树群落的种子雨、种子库及更新.植物生态学报,2000,24(4):402-407.
[22] 杨允菲,白云鹏,李建东,李丽.科尔沁沙地刺榆林种子散布的空间格局.应用生态学报,2010,21(8):1967-1973.
[23] 李建东,杨允菲.松嫩平原榆树疏林植物组成的结构型.草地学报,2003,11(4):177-282,300-300.
[25] 方开泰,许建伦.统计分布.科学出版社,1987:283-298.
[26] 杨允菲,祝玲.松嫩平原碱化草甸朝鲜碱茅种子散布机制的分析.植物学报,1995,37(3):222-230.
