密度结构对大青川红松人工林细根生物量与根长密度的影响
2013-09-06陈硕芃王韶仲王政权谷加存
陈硕芃,王韶仲,王政权,谷加存
(东北林业大学林学院,哈尔滨 150040)
红松是我国东北地区地带性顶级群落阔叶红松林中的主要树种,对于发挥区域尺度的生态和社会服务功能具有重要意义[1]。红松材质优良,寿命长,是培育优质大径材的重要人工林树种之一。建国60多年来,在红松人工林培育领域,国内陆续开展了广泛的研究,主要集中在红松生长节律和影响因子[2-3]、人工林群落生物量分配[4-6]、幼林抚育与林分生长和生物量积累[7-8],以及抚育间伐对林分生物量和生产力的影响[9-11]等多个方面的研究。林分抚育对红松人工林结构、生物量和生产力有重要的影响。李俊清和王业遽[12]对红松人工幼林研究表明,伐除50%阔叶树的林分总蓄积量和生产力均明显高于伐除全部阔叶树,分别相差28%和72%;张春锋等[10]对23a人工阔叶红松林的研究显示,中等间伐强度使得红松平均胸径和林分蓄积量分别提高了85%和61%。然而,以往研究主要关注了林分结构、抚育措施与林木地上部分(特别是干材)的联系,而对于树木根系,特别是细根(一般指直径≤2 mm)的生物量的影响了解较少。
树木的细根(及其共生真菌)不仅是养分和水分吸收的重要器官,而且是重要的碳汇(Carbonsink)。细根现存量占总生物量的比例不大,但其不断地进行着出生、生长、衰老和死亡的周转过程,因此,细根周转消耗了大量的光合产物[13-14]。据估计,全球尺度上,假定细根一年周转一次,将消耗陆地生态系统净初级生产力的30%左右[15]。同时,细根的死亡也是植物向土壤输入养分的重要途径,在一些森林生态系统中细根的凋落量超过了地上部分的一至数倍[16]。然而,由于细根取样、分析会消耗大量的时间和人力,与地上部分的研究相比,有关红松细根生物量与生产力的研究仍相对匮乏。虽然已有一些研究[17-22]对阔叶红松林和红松人工林的细根生物量及其动态进行了报道,但这些研究中均以直径小于2 mm作为划分细根的标准。近十年国内外树木根系研究显示,以直径小于2 mm来定义细根并据此估计其生物量、生产和周转存在着很大的误差[23-24]。因为根系统中实际上只有那些直径细小的、根系末端的低级根 (根尖为1级根,其母根为2级根,以此类推)具有吸收功能并进行着快速的周转[25-27]。Guo 等[26]对中国温带23个树种的研究显示,几乎所有树种只有前三级根才具有皮层组织,是主要的吸收根。根据他们的研究结果,红松前三级根的平均直径均小于0.5 mm,因此,选择直径小于0.5 mm来划分吸收根与非吸收根更具有代表性和生物学意义,但是未见采用此划分标准进行红松根系研究的相关报道。
本研究在红松核心分布区内的小兴安岭伊春市大青川林场,选择3种不同密度结构的红松人工林林分:低密度(简称“低红”,处理A),高密度(简称“高红”,处理B)和阔叶树占比例较大(简称“高阔”,处理C)。于2011年9月末通过根钻法,研究了不同直径等级(≤0.5 mm,0.5~1.0 mm,1.0~2.0 mm,>2.0 mm)根系生物量(SB,standing biomass)和根长密度(RLD,root length density),及其在不同土壤深度(土层)的空间分布规律。主要目的是比较林分密度结构对红松根系,特别是吸收根(≤0.5 mm)的SB和RLD的影响;同时也比较了林分密度结构对阔叶树种总体根系SB与RLD的影响。本研究旨在为红松人工林选择适宜的营林措施提供必要的理论依据。
1 材料与方法
1.1 研究地概况与样地设置
研究林分位于伊春市带岭区大青川林场(129°02'54~129°02'56E,46°58'57~ 45°59'03N)。该地区属温带大陆性季风气候,平均海拔315 m,坡度为10°~15°,属于小兴安岭南麓。年平均气温-0.3℃,1月平均温度-23℃,7月平均气温20.9℃。年平均降水量676 mm,年平均蒸发量805.4 mm,无霜期100~120 d。土壤为暗棕壤,平均土层厚度在30~40 cm。红松人工林栽植于1969年,株行距1.5 m×2 m。由于林木自然稀疏和先锋阔叶树种的入侵,形成了红松与阔叶树混交的局面,但是红松保留密度和阔叶树比例存在较大的差异。在林分踏查的基础上,于2012年9月末,在红松密度相对较低、相对较高以及阔叶树比例相对较高的三种林分类型中(简称为处理A、B、C),见表1。分别随机设置3个重复固定标准地(20 m×30 m),进行了林木和林地的土壤调查与分析。由表1可见,处理B中单位面积红松数量最高(株数比例76%),处理C中单位面积阔叶树数量最高(株数比例46%),而处理A的红松数量(株数比例64%)与C(株数比例54%)接近,阔叶树数量(株数比例36%)与B(株数比例24%)接近。天然更新的阔叶树种类主要包括水曲柳(Fraxinus mandshurica)、胡桃楸(Juglans mandshurica)、黄波罗(Phellodendronamurense)、山桃稠李(Prunus maackii)和少量的白桦(Betula platyphylla)。由于乔木层林冠庇荫,灌木和草本数量较少,盖度不足30%。

表1 不同密度结构红松人工林林分与土壤特征Tab.1 Characteristics of stand and soil in korean pine plantation with three densities
1.2 根系取样与形态测定
2011年10月中旬,在每块样地(共9块)内随机选择5个样点,去除地表凋落物,采用钢制土钻(内径5.8 cm)获取包含根系的土芯。考虑红松根系的分布特征[21],取样深度设为 30 cm,间隔10 cm取样(每个样点分3层)。每个样地内合计样品数为15,总计145个土芯。样品装入编好号的封口袋,置于冷藏箱内的冰块上,在4 h内带回实验室。在室内分析时,先将土芯放入小桶内充分浸泡(约2 h),以便于根系与土壤分离;随后用40目(0.42 mm)筛网在流水中冲洗土壤颗粒和杂质,最后放在直径15 cm、装有去离子水冰块的培养皿中,在低温下区分红松根系和阔叶树根系(二者差异非常明显)[18],按根系直径(≤0.5mm,0.5~1.0mm,1.0~2.0mm,>2.0mm)进行分级。阔叶树根系没有再按照树种进行区分,因为确认上存在较大的难度和误差。在每一样地内,随机选取1个样点的根系,按照取样层次,将红松和阔叶树根系依据直径等级,用EPSON EXPRESSION 10000XL彩色扫描仪进行扫描(分辨率设为400 dpi),用根系形态分析软件(WinRhizo 2004b,Regent instruments Inc,Canada)计算单个根的长度(mm)、平均直径(mm)和总长度(cm)。形态分析后的根系置于65℃烘干至恒重(0.000 1 g),然后计算比根长(单位生物量干重的根长)[28],依据单个土芯各直径等级的生物量干重和对应的比根长,计算各直径等级根系单位面积生物量和根长密度。
1.3 数据分析
数据计算和处理,均以单个土芯计算的单位面积生物量和根长密度数据为基础。将每一样地内各样点的3层(土芯)根系生物量和根长密度加和,作为该样点0~30 cm深度上的根生物量和根长密度,5个样点平均计算该样地的对应指标,然后3个重复样地相应指标平均,作为该处理下0~30 cm深度上的根系生物量和根长密度;相应地,各处理对应土壤深度(0~10 cm,10~20 cm,20~30 cm)上根系生物量和根长密度平均值也按前述方法计算。在单因素方差分析的基础上,采用多重比较的方法(LSD法),比较不同处理对0~30 cm土壤深度,以及各土壤层次红松和阔叶树各直径等级根系生物量和根长密度的影响;对于红松、阔叶树直径小于2 mm根系,采用三因素方差分析的方法,检验实验处理(3种密度结构)、直径等级(3个等级,≤0.5mm,0.5~1.0mm,1.0~2.0 mm)和土壤深度(3层,0~10 cm,10~20 cm,20~30 cm)对红松根系生物量和根长密度的影响,显著性水平设为P=0.05。数据统计分析采用软件SPSS for Windows 19.0(SPSS公司,美国)完成。
2 研究结果
2.1 不同处理根系生物量和根长密度比较
不同处理红松根系生物量和根长密度(0~30 cm土层之和)均存在差异。直径小于2.0 mm根系总的生物量在处理A(低红)、处理B(高红)和处理C(高阔)分别为300.9、456.2和221.8 g/m2,根长密度分别为2 611.3,4 914.5,1 965.3 m/m2;而大于2.0 mm根系总生物量在处理处理A、处理B和处理 C分别为464.8、262.0和533.3 g/m2,根长密度分别为74.0、168.3和83.0 m/m2。各处理间红松根系生物量和根长密度的差异主要表现在直径小于1.0 mm的根群上,如图1和图2所示。其中B处理吸收根(直径≤0.5 mm)生物量和根长密度均显著高于处理A和C,且0.5~1.0 mm根生物量和根长密度显著高于处理A;处理A和C在各直径级上均无显著的差异,但是直径≤0.5 mm和0.5~1.0 mm根群生物量和根长密度均比处理A较高。然而,1.0~2.0 mm和直径>2.0 mm直径根群生物量与根长密度在不同处理间均没有显著差异。
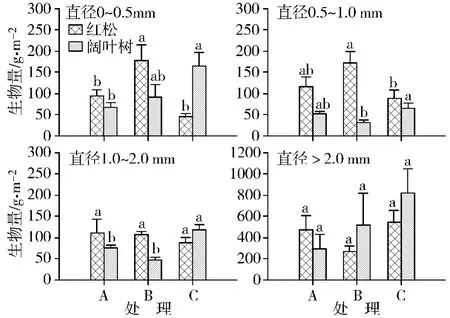
图1 不同处理下红松与阔叶树各径级根系生物量 (0~30 cm土层)处理A:低红,处理B:高红,处理C:高阔;相同径级内不同字母表示处理间差异显著,下同Fig.1 Standing biomass(0-30 cm soil depth)of roots in different diameter size in korean pine plantation with three densities.Treatment A:lower portion of korean pine,Treatment B:higher portion of korean pine,Treatment C:higher portion of hardwoods.Different lowercase letters within each root diameter size classes indicate significant difference among the treatments
相比较,各直径等级上,处理C阔叶树根系生物量和根长密度均高于处理A和B,而处理A和B之间均没有显著的差异。除了0.5~1.0 mm根群,处理C内阔叶树各径级根生物量均高于相应红松,而处理A和B则以红松占优势(处理B直径>2.0 mm根群除外),如图3所示;与根生物量的关系不同,各处理阔叶树直径≤0.5 mm和>2.0 mm根群的根长密度均大于红松,而各处理直径0.5~1.0 mm和1.0~2.0mm根群则以红松占优势(处理C中1.0~2.0 mm根群除外),如图4所示。

图2 不同处理下红松与阔叶树各径级根系根长密度 (0~30 cm土层)Fig.2 Length density(0-30 cm soil depth)of roots in different diameter size in korean pine plantation with three densities.Labels are same to Fig.1

图3 不同处理红松人工林各径级根系生物量空间分布特征Fig.3 Standing biomass of roots in different diameter size along soil depths in korean pine plantation with three densities.Different lowercase letters within each soil depth indicate significant difference among the treatments
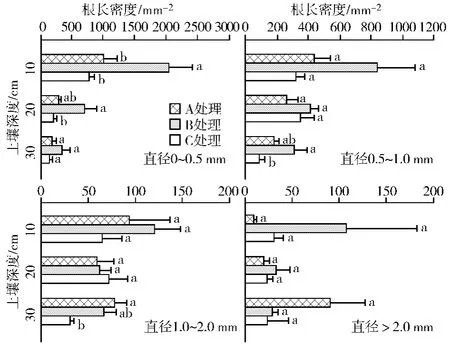
图4 不同处理红松人工林各径级根系根长密度空间分布特征Fig.4 Length density of roots in different diameter size along soil depths in korean pine plantation with three densities.Different lowercase letters within each soil depth indicate significant difference among the treatments.Labels are similar to Fig.3
2.2 不同处理根系生物量和根长密度垂直分布的差异
对直径小于2.0 mm根系而言,各处理内不同径级根系生物量与根长密度均随着土壤深度增加而减少;而直径大于2.0 mm根系在各处理间没有一致的规律性。直径小于2.0 mm根系生物量和根长密度主要集中在土壤表层0~10 cm,在处理A中直径≤0.5 mm、0.5~1.0 mm和1.0~2.0 mm根群生物量分布在表层的比例分别占0~30 cm总生物量(直径≤2.0 mm)的 65.9%、48.4%和41.7%;处理B中对应比例为61.5%、50.9%和44.4%,处理C为49.8%、44.9%和37.3%。根长密度也表现出类似的规律。此外,根系生物量在处理间的差异,主要体现在表层的0~0.5 mm和0.5~1.0 mm根群中。处理C与处理A和B相比,红松吸收根(直径≤0.5 mm)生物量在表层分布的比例明显减少(处理 A、B和 C分别为65.9%、61.5%和49.8%),而在20~30 cm则明显增加(处理 A、B和 C分别为 13.7%、11.6%和23.8%),但是根长密度的分布则没有如此大的变化。直径大于2.0 mm根群生物量和根长密度在各处理间没有显著的差异,且在各土层处理间没有一致的规律出现。
2.3 影响根系生物量和根长密度的因子分析
对于直径小于2.0 mm的各根群,除了生物量没有受到直径等级的显著影响外,林分密度结构、直径等级和土壤深度对根系生物量和根长密度均存在着显著的影响,见表3。此外,这些因子之间的交互作用,均对根系生物量和根长密度有显著的影响(P<0.05),表明林分密度结构对根生物量和根长密度的影响随着直径等级和土壤深度而发生改变。然而,林分密度结构、直径等级和土壤深度三者之间的交互作用则不显著。

表2 林分处理、直径等级和土壤深度 (土层)对红松根系 (≤2 mm)生物量和根长密度三因素方差分析表Tab.2 Results of three-way ANOVA(analysis of variance)testing the effect of stand density,diameter size,and soil depth on fine root(≤2 mm)standing biomass and length density
3 讨论
本研究中,不同密度结构红松人工林细根(直径≤2.0 mm)生物量(0~30 cm土层)为222~456 g/m2,存在较大的差异,但是波动范围处在以往细根(≤2.0 mm)研究结果的范围之内。杨丽韫和李文华[18]的研究表明,长白山天然阔叶红松林在10月份细根生物量(0~50 cm土层)约为250 g/m2,稍高于本研究中的低密度红松林分(处理A);朱胜英等[21]报道帽儿山地区红松人工林细根生物量(0~30 cm土层)为263 g/m2,这比本研究高阔叶树林分(处理C)稍高,而稍低于密度红松林分(处理A);郭忠玲等[20]对长白山天然阔叶红松林细根生物量(0~20 cm土层)的研究结果为449 g/m2,与高密度红松林分(处理B)的细根生物量很接近;但是,单建平等[17]等对长白山天然阔叶红松细根生物量(0~30 cm土层)为504.9 g/m2,高于本研究中的最高值和其他类似的研究。因此,通过比较可以发现,林分类型(天然林与人工林)、林龄和林分密度结构以及研究者的根系取样深度,均是导致红松细根生物量出现差异的原因[22]。尽管本研究是基于一次取样,但是获得的结果与其他研究具有较好的可比性。
根长密度是评价树木养分吸收能力的一个重要指标[23-29]。本研究显示,吸收根(直径≤0.5 mm,0~30 cm)的根长密度在高红(处理B,3 102.2 m/m2)中显著高于低红(处理A,1495.9 m/m2)和高阔(处理C,1036.7 m/m2)林分。然而,以往红松根系研究中很少关注了根长密度的变异,使得无法进行树种内的比较。但是,帽儿山地区的研究显示,9月份水曲柳和落叶松人工林前3级根根长密度现存量分别约为2 000 m/m2和6 400 m/m2(仅包括0~10 cm土层)[29],如果考虑整个土壤剖面上的分布,那么这两个树种的根长密度可能均高于红松根系。不过,若考虑到阔叶树根系的存在,则本研究中红松人工林内所有木本植物根系的根长密度(处理 A、B和 C分别为4 763.9、7 471.8和9 019.1 m/m2)将远高于落叶松,而接近(处理B)或者高于水曲柳(处理C)的根长密度,这表明人工天然混交林分中树木对土壤资源的利用上可能更加充分,体现出了混交林比纯林在养分利用上的优势[30]。
林分结构的改变,不仅影响干材的生长和生物量[7,10],也 对 根 系 的 生 长 和 分 布 有 重 要 的 影响[31-33]。结果显示,在直径小于0.5 mm和0.5~1.0 mm根群中,高红(处理B)内红松根系生物量均高于其他2个处理。考虑到树木平均胸径和树高与低密度红松(处理A)接近,说明在相对较高的密度条件下,红松根系仍然有较为充分的生长空间,而不会对其地上部分的生长产生不利影响。此外,通过单位面积红松细根生物量与现存株数密度相比,发现单株水平上,直径小于0.5 mm根群的生物量处理 A、B和 C分别为886.4、950.3和383.1 g。这说明了一个非常重要的问题:较高密度下并没有显著降低红松单株地上部分(胸径和树高)的生长,而且促进了其吸收根的构建(处理B);而在红松低密度下,虽然地上部分生长有所增加,但是单株水平上吸收根系并不能充分地利用林地资源,造成地力的浪费(处理A);但是阔叶树的比例如果占的过高,则会严重地限制红松单株水平上吸收根系的发展,这可能主要是由阔叶树根长密度不成比例的增加引起的(处理C)。这些发现,加深了对林分结构如何影响红松林分、个体水平上吸收根生物量分配和资源获取这一问题的理解。
各处理林分中,直径小于2.0 mm根系生物量和根长的分布,均以表层(0~10 cm)所占比例最高(生物量:42.9%~53.5%,根长密度:54.1%~61.2%),其他研究也发现了相似规律。朱胜英等[21]的研究显示,帽儿山地区红松人工林中表层0~10 cm细根生物量占0~30cm土层的50.3%;杨丽韫等[19]研究了长白山原始阔叶红松林细根的垂直分布特征,指出0~10 cm土层红松和其他主要阔叶树种细根生物量占整个0~50 cm取样深度的68%。根系这样的垂直分布格局有助于其在资源丰富的表层更好地吸收养分或水分。但是,值得注意的是,与处理A和B相比,处理C中阔叶树所占比例的大幅增加,明显地降低了红松所有直径等级细根(直径≤2.0 mm)在0~10 cm的分布比例,尤其是吸收根(直径≤0.5 mm)。吸收根不仅在表层分布比例下降,而且在20~30 cm的分布比例增加。这说明,红松人工林密度结构的改变,会在较大程度上改变吸收根生物量的垂直分配格局,暗示了降低阔叶树组成比例的抚育间伐措施,将改善红松个体的养分利用状况,进而促进林分水平生物量和蓄积的提高。
4结论
通过比较密度结构对红松人工林根系生长的影响,并结合林分生长状况,在该研究区,维持人工林中红松适宜的株数密度,并保留一定的阔叶树,能够促进林木个体在地上和地下部分的生长,从而维持林分水平上较高的生物量和蓄积量。密度结构对根系生物量和根长密度的影响,主要表现在直径小于1.0 mm,特别是小于0.5 mm的吸收根上,这说明林分结构对吸收根生长的影响更强,而对直径相对较大的、以储存功能为主的根系影响较弱。因此,如果按照以往的细根划分标准(如直径≤2.0 mm),可能将这些本来没有差异的根群(如1.0~2.0 mm)包括进去,掩盖了一些重要的信息。未来红松根系生物量与周转的研究中,建议选择直径小于0.5 mm的根群作为细根研究中的基本取样单元。
】
[1]屈红军,牟长城.东北地区阔叶红松林恢复的相关问题研究[J].森林工程,2008,24(3):17-20.
[2]李景文,刘庆良.红松生长及其与气侯条件关系的研究[J].东北林业大学学报,1975(1):1-15.
[3]李克志.红松生长过程的研究[J].林业科学,1983,19(2):127-135.
[4]丁宝永,孙继华.红松人工林生态系统生物生产力及养分循环研究[J].东北林业大学学报,1989,17(2):35-39.
[5]李俊清,柴一新,张东力.人工阔叶红松林的结构与生产力[J].林业科学,1990,26(1):1-7.
[6]陈效群,刘文和,于源贵,等.阔叶红松森林群落生态位分类的研究[J].林业科技,1995,20(3):4-6.
[7]张云江,乔国宝,付振海,等.红松人工林透光抚育对林木生长与结实的影响[J].辽宁林业科技,1996,23(2):24-26.
[8]曾明洪,熊建宏,杨清培,等.南岭山地杉木人工林与天然林群落特征研究[J].江西林业科技,2008(5):10-15.
[9]胡万良,谭学仁.抚育间伐对红松人工林生物量的影响[J].辽宁林业科技,1999(2):13-16.
[10]张春锋,殷鸣放,孔祥文,等.不同间伐强度对人工阔叶红松林生长的影响[J].辽宁林业科技,2007,33(1):12-15.
[11]刘明国,韩玉芬,胡万良,等.红松人工林不同间伐强度效果分析[J].福建林业科技,2008,35(2):47-51.
[12]李俊清,王业遽.红松人工幼树透光伐效果的分析[J].应用生态学报,1991,2(1):8-13.
[13]Fahey T J,Hughes J W.Fine root dynamics in a northern hardwood forest ecosystem,Hubbard Brook Experimental Forest,NH[J].Journal of Ecology,1994,82(3):533-548.
[14]Hendricks J J,Hendrick R L,Wilson C A,et al.Assessing the patterns and controls of fine root dynamics:an empirical test and methodological review[J].Journal of Ecology,2006,94(1):40-57.
[15]Jackson R B.Mooney H A.Schulze E D.A global budget for fine root biomass,surface area,and nutrient contents[J].Proceedings of the National Academy of Sciences of the United States of America,1997,94(14):7362-7366.
[16]Steinaker D F,Wilson S D.Belowground litter contributions to nitrogen cycling at a northern grassland-forest boundary[J].Ecology,2005,86(10):2825-2833.
[17]单建平,陶大立,王 淼.长白山阔叶红松林细根周转的研究[J].应用生态学报,1993,4(3):241-245.
[18]杨丽韫,李文华.长白山原始阔叶红松林细根分布及其周转的研究[J].北京林业大学学报,2005,27(2):1-5.
[19]杨丽韫,罗天祥,吴松涛.长白山原始阔叶红松(Pinus koraiensis)林及其次生林细根生物量与垂直分布特征[J].生态学报,2007,27(9):3609-3617.
[20]郭忠玲,郑金萍,马元丹,等.长白山几种主要森林群落木本植物细根生物量及其动态[J].生态学报,2006,26(9):2855-2862.
[21]朱胜英,周 彪,毛子军,等.帽儿山林区6种林分细根生物量的时空动态[J].林业科学,2006,42(6):13-19.
[22]王树堂,韩士杰,张军辉,等.长白山阔叶红松林表层土壤木本植物细根生物量及其空间分布[J].应用生态学报,2010,21(3):583-589.
[23]Pregitzer K S,Deforest J L,Burton A J,et al.Fine root architecture of nine North American trees[J].Ecological Monographs,2002,72(2):293-309.
[24]Guo D,Li H,Mitchell R J,et al.Fine root heterogeneity by branch order:exploring the discrepancy in root turnover estimates between minirhizotron and carbon isotopic methods[J].New Phytologist,2008,177:443-456.
[25]Guo D,Xia M,Wei X,et al.Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species[J].New Phytologist,2008,180(3):673-683.
[26]Xia M,Guo D.Pregitzer K S,Ephemeral root modules in Fraxinus mandshurica[J].New Phytologist,2010,188(4):1065-1074.
[27]王向荣,王政权,韩有志,等.水曲柳和落叶松不同根序之间细根直径的变异研究[J].植物生态学报,2006,29(6):871-877.
[28]刘 佳,项文化,徐 晓,等.湖南会同5个亚热带树种的细根构型及功能特征分析[J].植物生态学报,2010,34(8):938-945.
[29]Wang Z,Guo D.Wang X,et al.Fine root architecture,morphology,and biomass of different branch orders of two Chinese temperate tree species[J].Plant and Soil,2006,288(1):155-171.
[30]Richards A E,Forrester D I,Bauhus J,et al.The influence of mixed tree plantations on the nutrition of individual species:a review[J].Tree Physiology,2010,30(9):1192-1208.
[31]杨丽韫,李文华.长白山不同生态系统地下部分生物量及地下C 贮量的调查[J].自然资源学报,2003,18(2):204-209.
[32]汪金松,范秀华,范 娟,等.地上竞争对林下红松生物量分配的影响[J].生态学报,2012,32(8):2447-2457.
[33]江民锦.不同间伐强度对杉木人工林下植被发育及生物量的影响[J].江西林业科技,2001(4):5-8.
