鸡IGFBP-3基因多态性及其与生产性能的相关性
2013-08-23金崇富葛兆建杨智青陈长宽陈应江
金崇富, 葛兆建, 杨智青, 时 凯, 陈长宽, 陈应江
(江苏沿海地区农业科学研究所,江苏 盐城 224002)
胰岛素样生长因子结合蛋白-3(IGFBP-3)主要存在于血液中,它对IGF-I和IGF-II具有很高的亲和力,是血液中最为丰富的IGFBP结合形式。血液中95%的IGF-I和IGF-II都与IGFBP-3结合,因此,IGFBP-3对于 IGF作用的发挥起着重要作用[1]。IGFBP-3属于超家族中的高亲和力类型,基因定位于2p12214,mRNA长2.4 kb,多肽链由264个氨基酸(含18个Cys)组成,分子量约29 000,一级结构中存在3个N2型糖基化位点和肝素结合位点,在Ser位点上可以被磷酸化,其氨基酸序列中还存在入核定位序列(NLS)。目前有关IGFBP-3基因多态性的研究主要集中在牛[2]、羊[3-4]和猪[5]上,有关鸡IGFBP-3基因的多态性研究鲜有报道,仅检索到金崇富等[6]和俞亚波等[7]的报道。本研究拟利用PCR-SSCP、PCR-RFLP及DNA测序技术,对IGFBP-3基因多态性进行研究,以期为鸡繁殖、生长发育性状的遗传标记研究提供资料。
1 材料与方法
1.1 试验材料
200只母鸡血样采自江苏沿海地区农业科学研究所生态养殖基地,同时记录其生长性状(初生重,4、8、12、16 周龄体质量)及繁殖性状(开产日龄、开产体质量、开产蛋质量、300日龄产蛋数)。翅静脉采集血样1.5 ml,肝素钠抗凝,-20℃保存。常规苯酚-氯仿法抽提基因组DNA,对基因组DNA进行OD值测定,计录浓度后备用。
1.2 引物设计及PCR扩增
根据鸡的IGFBP-3基因序列(GenBank登录号为NC_006089)设计IGFBP-3基因8个外显子的测序用引物P1~P10,用于直接测序。引物由上海生工生物工程有限公司合成。由于第4外显子较长,序列比较特殊,没能设计出合适的引物,故本试验没有对第4外显子进行研究。10对引物的详细信息见表1。

表1 引物信息Table 1 The information of primers used in this study
1.3 DNA 池构建
在稀释好的200份DNA样品中,每份取5 μl混合在同一管内,作为池DNA,用于单核苷酸多态性(SNP)的搜索[8]。
1.4 PCR 扩增
IGFBP-3基因引物最佳PCR扩增体系:DNA 1.0 μl,10 × Buffer 2.0 μl,Mg2+(25 mmol/L)2.2 μl,上下游引物(10 pmol/μl)各 1.0 μl,dNTPs(10 mmol/L)0.8 μl,Taq DNA 聚合酶(2 U/L)0.2 μl,ddH2O 11.8 μl,总体系 20.0 μl。
IGFBP-3基因引物PCR反应循环参数:95℃预变性6 min;95℃变性30 s,退火(温度见表1)30 s,72℃延伸30 s,30个循环;72℃延伸10 min,10℃保存。
1.5 PCR-SSCP 分析
将2.5 μl PCR产物加入7.5 μl的变性缓冲液中,98℃变性10 min,然后迅速冰浴5 min。将变性好的PCR产物点样加入到10%的聚丙烯酰胺凝胶中,250 V电压预电泳10 min后在140 V电压下电泳8~24 h。电泳结束后银染显色并拍照。
1.6 PCR-RFLP 分析
MspⅠ 酶切反应体系:PCR扩增产物10.0 μl,限制性内切酶 Msp Ⅰ 0.4 μl,10 × Buffer 2.0 μl,ddH2O 7.6 μl,总体系20.0 μl,37 ℃酶切过夜,酶切产物用10%的聚丙烯酰胺电泳分析,电泳结束后银染显色并拍照。
1.7 数据分析
配合下列模型进行最小二乘方差分析,比较鸡IGFBP-3各基因型之间的差异(对各引物的基因型单独分析):yij=μ+Gi+eij,其中:yij为性状测定值;μ为群体平均值;Gi为基因型效应;eij为随机残差效应。
所有统计分析均采用SPSS统计软件的GLM(General linear model)过程完成,多重比较采用LSD法。
2 结果
2.1 DNA测序结果
以DNA为模板,用引物P1~P8扩增的产物直接测序,结果表明,在第160 bp(引物P1扩增结果)和1 087 bp(引物P2-1扩增结果)处出现双峰,表明在这2个位置发生突变(图1),分别记为T160G位点和C1087T位点。其中在第160 bp处突变位于第1外显子和第1内含子连接部分,属于内含子突变。

图1 PCR产物直接序列结果Fig.1 The sequencing results of PCR products
2.2 酶切位点的发现
通过已知突变利用Primer premier 5软件的Enzyme功能寻找酶切位点,结果发现C1087T位点的多态性导致了MspⅠ酶切位点的突变(图1)。
2.3 PCR-RFLP 结果
IGFBP-3基因第2外显子1 087 bp处(C1087T位点)的C→T的突变,产生了MspⅠ酶切位点(C↓CGG)多态性,经酶切消化后产生大小分别为161、161/95/66、95/66的3种带型(图2),分别命名为CC、CT、TT 基因型。
2.4 PCR-SSCP 结果
P1引物扩增片段采用PCR-SSCP分析,结果表明,P1扩增片段有3种带型,定义为AA、AB和BB(图3)。
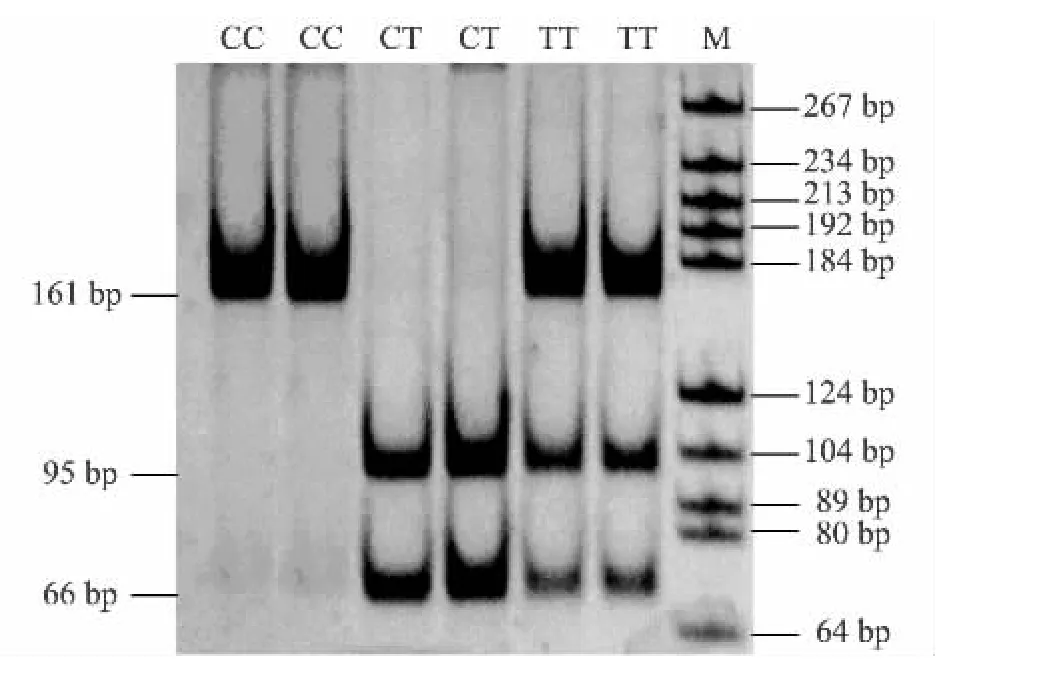
图2 IGFBP-3基因第2外显子P2-1引物的PCR产物MspⅠ酶切结果Fig.2 Identification of PCR products of exon 2 in IGFBP-3 gene by MspⅠdigestion
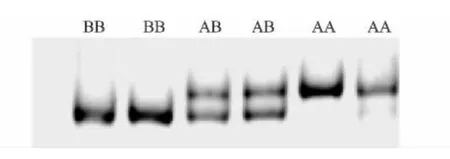
图3 引物P1的SSCP分析结果Fig.3 SSCP analysis of primer P1 by PCR
2.5 序列分析结果
PCR产物片段测序结果表明,BB型与AA型相比在160 bp处发生了T→G突变(图4);TT型与CC型相比在1 087 bp处发生了C→T突变(图 5)。

图4 P1引物扩增产物AA与BB型的序列比较Fig.4 Sequence alignment of genotypes AA and BB in chickens using primer P1

图5 P2-1引物扩增产物CC与TT型的序列比较Fig.5 Sequence alignment of genotypes CC and TT in chickens using primer P2-1
2.6 IGFBP-3基因与生产性能的关联分析
2.6.1 IGFBP-3基因T160G位点与生长性状的关联分析 T160G位点不同基因型对京海黄鸡母鸡生产性能的关联分析见表2。AA、AB、BB 3种基因型在开产体质量、开产蛋质量、300日龄产蛋数、4周龄体质量12周龄体质量16周龄体质量方面均无显著差异(P>0.05),AB基因型开产日龄显著低于AA基因型(P<0.05),初生重显著高于BB基因型(P<0.05),8周龄体重显著高于AA基因型(P<0.05)。
2.6.2 IGFBP-3基因C1087T位点与生长性状的关联分析 C1087T位点不同基因型对母鸡生产性能的关联分析见表3。CC、CT、TT 3种基因型在开产体质量、开产蛋质量、300日龄产蛋数、初生质量以及4、8、12、16周龄体质量方面均无显著差异(P>0.05)。仅CT基因型开产日龄显著低于TT基因型(P <0.05)。
3 讨论
3.1 内含子突变的影响
内含子在原始转录产物的加工过程中被切除,不包含在成熟mRNA的序列中,在每个外显子和内含子的接头区,有一段高度保守的共有序列,即每个内含子的5'端起始的两个核苷酸都是GT,3'端末尾的两个核苷酸都是AG,这是RNA剪接的信号,这种接头形式称为GT-AG法则[9]。近年来人们对内含子的功能提出许多新的见解,其中最重要的一个方面是内含子在基因表达调控中起着重要作用[9]。目前在许多基因的内含子中均发现了基因表达的调控元件,其中大多为增强元件,通常认为基因的第1个内含子对其表达具有重要的调控作用。基因的编码区突变以及内含子与外显子交界处的突变可能导致基因的结构、功能发生变化[10]。
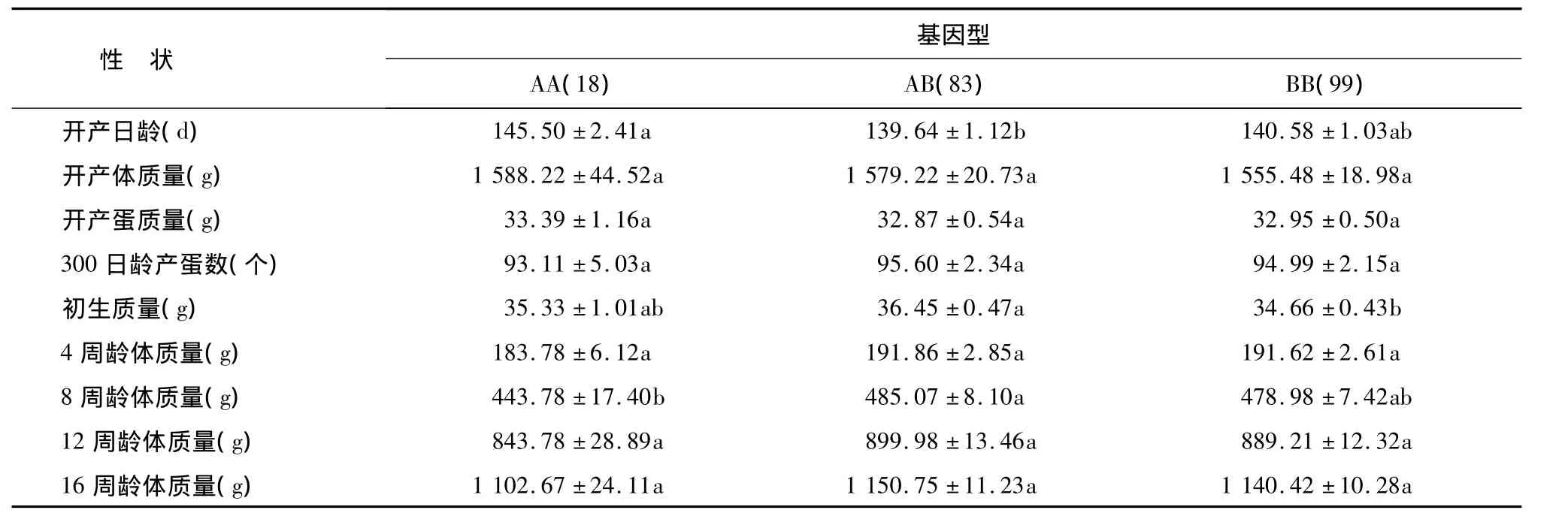
表2 T160G位点基因型与生产性能的相关性Table 2 Association of genotypes with production performance based on T160G site

表3 C1087位点基因型与生产性能的相关性Table 3 Association of genotypes with production performance based on C1087 site
本试验在IGFBP-3基因的外显子1和内含子1结合处发现了1个SNP位点,即内含子1的5'端起始的第2个核苷酸T突变为G,打破了 GT-AG法则,从而影响了RNA剪接的信号,可能导致IGFBP-3基因的结构和功能发生改变,进而对IGFBP-3基因的表达产生影响,最终影响鸡的生产性能。
3.2 IGFBP-3基因的多态性及其与生产性能的关联分析
1997年,Maciulla等[11]利用 PCR-RFLP方法首次报道IGFBP-3基因的651片段的HaeⅢ酶切多态性,经序列分析发现,第299 bp处的C→A的颠换(GGCC→GGAC)导致一个HaeⅢ限制性酶切位点的丢失而产生了HaeⅢ酶切多态性。张永宏等[12]以草原红牛为研究对象,采用PCR-SSCP方法检测IGFBP-3基因的多态性,并将不同基因型与部分屠宰性状进行相关分析,结果在第2内含子处发现多态性位点,该位点的多态性对草原红牛的眼肌面积有显著影响(P<0.05)。沈敏等[13]采用PCR-SSCP方法对中国美利奴羊和哈萨克羊中IGFBP-3基因的多态性进行了检测,并对不同基因型与中国美利奴羊部分羊毛性状间的关联性进行了分析,结果表明,不同基因型对部分羊毛性状有一定影响。李美玉等[14]利用PCR-RFLP技术对7个山羊群体的501个个体进行了检测,结果发现:该位点与断奶山羊质量和眼肌面积显著相关(P<0.05),与3月龄体长和胸围极显著相关(P<0.01),与10月龄体长、胸围和12月龄体长显著相关(P<0.05)。张润峰等[15]以IGFBP-3基因作为中国荷斯坦牛部分泌乳性状的候选基因,对中国荷斯坦牛群体中IGFBP-3基因座多态性与泌乳性状进行相关分析,结果发现IGFBP-3基因座与产奶量、乳蛋白率和体细胞评分显著相关(P<0.05)。高雪等[16]采用PCR-SSCP技术分析了牛类胰岛素生长因子结合蛋白-3(IGFBP-3)基因在南阳牛、鲁西牛和中国西门塔尔牛3个牛品种中的遗传多态性,结果表明:第2外显子在第8 069 bp处发生单碱基突变T→C,并导致苯丙氨酸变为亮氨酸。
本试验对鸡IGFBP-3基因2个SNPs位点与繁殖性状(开产日龄、开产体质量、开产蛋质量和300日龄产蛋数)进行关联分析,结果显示:T160G位点AB基因型开产日龄显著低于AA基因型(P<0.05);C1087位点CT基因型开产日龄显著低于TT基因型(P<0.05)。初步推测IGFBP-3基因T160G和C1087可作为鸡早期产蛋性状的一个候选分子标记,用于标记辅助选择。
对鸡IGFBP-3基因2个SNPs位点与生长性状(初生质量、4、8、12、16 周龄体质量)的关联分析结果显示:T160G位点AB基因型初生重显著高于BB基因型(P<0.05),8周龄体质量显著高于AA基因型(P<0.05),AB基因型为体质量的优势基因型,但AB基因型为杂合子,交配后会发生基因分离的现象,不能固定,生产中难以利用该位点来提高鸡的体质量。C1087位点对鸡的生长性状没有显著影响。
[1]SHAW L C,GRANT M B.Insulin like growth factor-1 and insulin-like growth factor binding proteins:Their possible roles in both maintaining normal retinal vascular function and in promoting retinal pathology[J].Endocrine & Metabolic Disorders,2004,5:199-207.
[2]刘 哲,吴建平,马彦男,等.奶牛 IGFBP-3基因部分序列PCR-SSCP多态性与产奶量和生长性能的相关性[J].农业生物技术学报,2009,17(3):445-450.
[3]于 娇,陈 宏,蓝贤勇,等.5种家畜IGFBP-3基因遗传变异研究[J].西北农林科技大学学报:自然科学版,2007,35(3):33-37.
[4]胡沈荣,于 娇,蓝贤勇,等.西农萨能奶山羊IGFBP-3多态与产羔数、体尺性状的关联分析[J].西北农林科技大学学报:自然科学版,2007,35(11):1-5.
[5]刘德武.猪生长相关基因的单核苷酸多态性研究[D].广州:华南农业大学,2005.
[6]金崇富,王金玉,王慧华,等.京海黄鸡IGFBP-3基因外显子1和内含子1部分序列多态性及其与生长繁殖性状的相关性[J].中国畜牧兽医,2010,37(9):132-135.
[7]俞亚波,王金玉,金崇富,等.6个鸡种IGFBP-3基因第2外显子MspⅠ酶切位点的遗传多态性研究[J].中国家禽,2011,33(17):15-18.
[8]崔建勋,杜红丽,张细权,等.利用DNA池和测序技术快速筛查SNPs及估算基因频率[J].遗传学报,2005,32(4):372-377.
[9]李 宁,陈 宏.动物遗传学[M].北京:中国农业出版社,2002.
[10]邱晓云,卢大儒.内含子在基因表达调控中的作用[J].国外医学:遗传学分册,1996,19(1):44-48.
[11]MACIULLA J H,ZHANG H M,DENISE S K.A novel polymorphism in the bovine insulin-like growth factor binding protein-3(IGFBP-3)gene[J].Anim Genet,1997,28:375.
[12]张永宏,高 妍,赵志辉,等.草原红牛IGFBP-3基因多态性及与部分屠宰性状的相关性分析[J].中国兽医学报,2010,30(4):549-551.
[13]沈 敏,王文君,李 宁,等.IGFBP-3基因多态性及其与中国美利奴羊部分羊毛性状的关联性分析[J].遗传,2008,30(9):1182-1186.
[14]李美玉,李 兰,闵令江,等.山羊IGFBP-3基因的遗传分析及其与经济性状的关系[J].畜牧兽医学报,2008,39(12):1647-1653.
[15]张润峰,陈 宏,雷初朝,等.IGFBP-3基因PCR-RFLP多态性与中国荷斯坦牛泌乳性状的相关分析[J].中国畜牧杂志,2006,42(3):9-11.
[16]高 雪,徐秀容,许尚忠,等.牛类胰岛素生长因子结合蛋白3基因PCR-SSCP分析[J].农业生物技术学报,2006,14(4):474-477.
