宁夏荒漠草原植物群落的种间关系
2013-04-25谢应忠许冬梅
李 潮,谢应忠,许冬梅,李 侠
(宁夏大学农学院,宁夏 银川 750021)
种间关系指不同物种种群之间相互作用所形成的关系,是植物种群间相互联系、相互影响的反映,为植物群落结构和数量的重要特征之一[1]。在群落生态学研究中,通过对种间关系进行分析,可以确定趋向于生活在一起的物种,进而确定群落边界。因此,测定植物群落的种间关系,能够揭示植物种群间在空间上共同出现的程度,从而辨析植物是独立分布,还是相互吸引或相互排斥[2]。近年来,有关宁夏东部风沙区荒漠草原植物群落数量分析的研究主要集中在生态位测度[3-4]和群落排序[5]等方面,但对植物群落种间关系方面的研究未有深入报道。本研究通过实地调查,定量研究宁夏东部风沙区荒漠草原植物群落的种间关系,旨在阐明群落的组成和动态,完善该地区群落生态学的研究,可进一步为区域生态环境建设、生态安全维护提供理论指导,对该地区草地生态系统的可持续利用具有实用意义。
1 材料与方法
1.1研究区自然概况 研究区位于宁夏回族自治区盐池县(106°30′-107°47′ E,37°04′-38°10′ N),海拔1 295~1 951 m。地势南高北低,北临毛乌素沙地,南靠黄土高原,北侧为鄂尔多斯缓坡丘陵,南侧为黄土丘陵。
盐池县属于典型的中温带大陆性气候,光能丰富、热量偏少,年平均气温7.5 ℃,年均无霜期165 d。年平均风速2.8 m·s-1,冬春风沙天气较多。多年平均降水量280 mm,自西北向东南递增,其中70%降水集中在6-9月,且降水年际变率大。灰钙土为主要的土壤类型,其次是风沙土和黑垆土,土壤质地以沙土、沙壤和粉沙壤为主[5]。植被以荒漠草原为主,风蚀作用强烈。
1.2数据分析
1.2.1植物群落调查 2012年7月下旬在宁夏盐池县进行野外调查。采用随机取样法进行取样,结合样方法与样线法对植物群落进行调查,样线沿流动沙丘外围向里布设,在样线上根据群落变化布设样方,样方面积为1 m×1 m。共布设样方119个,测定植被盖度、密度、多度、频度,同时记录样方所在的地理位置、微地形等。
重要值(IV)计算公式为:IV=(相对盖度+相对密度+相对高度+相对频度)×100/4[3]。
1.2.2分析方法 依据调查结果,剔除频度<10%的物种后[6],以主要物种的重要值作为数量指标,用于种间关联的相关分析。
Jaccard关联指数:按照物种在样方内存在与否,获得种-样方的“0、1”二元矩阵,“0”代表不存在,“1”代表存在,用于计算种间关联。依据2×2列联表的a、b、c值计算Jaccard关联指数(JI),JI=a/(a+b+c)。式中,a为两个种都存在的样方数,b为B种存在而A种不存在的样方数,c为A种存在而B种不存在的样方数。
种间关联的2×2列联表及χ2检验:在2×2列联表的基础上,计算χ2值,可以判断两个物种关联与否。
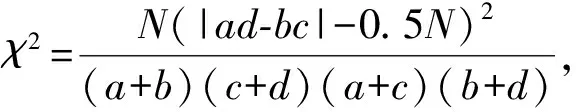


数据处理用Microsoft Ecxel 2007及SPSS 17.0数据处理软件完成。
2 结果与分析
2.1宁夏东部风沙区荒漠草原植物群落特征 在所有调查样方中,共鉴定出74种植物,分属21科57属,剔除频度<10%的物种后,剩余21种植物(表1)。其中,多年生草本占52.4%,一年生草本占33.3%,半灌木占14.3%。种数最多的3个科是禾本科、豆科和藜科。有一部分物种具有较为广泛的地理分布范围,如赖草在70个样地中出现,丝叶山苦荬出现在68个样地,黑沙蒿出现在62个样地。
2.2宁夏东部风沙区荒漠草原植物群落的种间关系 宁夏东部风沙区荒漠草原植物群落21个主要物种210个物种对数间的χ2检验、Pearson相关系数和Spearman秩相关系数的检验结果显示,χ2检验共有21个物种对数为显著正关联,25个物种对数为显著负关联;Pearson相关系数有12个物种对数为显著正相关,6个物种对数为显著负相关;Spearman秩相关系数有21个物种对数为显著正相关,10个物种对数为显著负相关(表2)。
2.2.1χ2检验与Jaccard关联指数 校正后的χ2检验值排出的半矩阵图表达的是种间联结的程度,这有助于对群落中植物种间关系的认识。图1所示,正关联的物种对数为103个,负关联的物种对数为106个,无关联的物种对数为1个。正、负关联物种对数分别占总物种对数的49.05%和50.48%。物种间呈正关联可能是因一个物种依赖于另一个物种而存在,或两者受生物因子和非生物因子影响而共存。负关联则可能是因竞争、空间排挤或他感作用,或不同的环境要求[7]。负关联物种对数在植物群落中的出现率较大,说明该区域的植物群落不稳定,当受到外界因素干扰时,容易发生波动,群落仍处于发展演替之中。
χ2检验物种对数间呈极显著、显著关联的物种对数有46个,占总物种对数的21.9%。其中物种对数为显著正关联共有21个,占总物种对数的10.0%,物种对数为显著负关联共有25个,占总物种对数的11.9%,呈显著正、负关联的物种对数相差较小。群落系统发育或群落演替过程愈成熟,其植物种类组成也愈稳定,种间关系也愈趋向正关联,以保证物种间的共存[8]。物种对数间呈极显著、显著关联的物种对数不多,说明该地区植物种群之间的关联性较弱。
Jaccard关联指数的半矩阵图表达的是物种共同出现的百分率。由图2看出,JI为0的物种对数为22个,占总物种对数的10.5%;JI值在0~0.2的物种对数有150个,占总物种对数的71.4%,说明植物群落的主要物种间关联程度很小,存在相当程度的独立分布格局。同时,JI值大于0.4的物种对数为2个,仅占总物种对数的0.95%,这也进一步说明该地区植物群落的种间关联性相对较弱。
2.2.2Pearson 相关分析与Spearman秩相关分析χ2检验只涉及种的存在与否,不能反映物种对数间的数量变化关系,因此,采用Pearson 相关系数和Spearman 秩相关系数进一步定量分析植物群落的种间关系。通过Pearson相关分析,呈显著相关的物种对数为18个,其中呈极显著正相关的6个,呈显著正相关的6个,呈极显著负相关的两个,呈显著负相关4个(图3、表2)。Spearman秩相关分析表明,呈显著相关的物种对数为31个,其中呈极显著正相关的18个,呈显著正相关的3个,呈极显著负相关的5个,呈显著负相关5个(图4、表2)。Spearman秩相关分析和χ2检验得出的呈显著相关的物种对数均多于Pearson相关分析的结果(表2),原因在于,基于Pearson相关系数的统计分析对于数据的分布类型十分敏感,只有在数据呈近似正态分布的时候才能被采用,否则就可能导致错误的结论。而自然界大多数物种多呈簇生分布,所以Pearson相关分析的结果带有一定的局限性[9]。
比较图1和图4,在Spearman秩相关分析和χ2检验中,呈正相关的物种对数的数量相同,但χ2检验中呈显著正相关的物种对数在Spearman秩相关分析中均为极显著正相关。而对于呈显著负相关的物种对数的数量,χ2检验明显多于Spearman秩相关分析,这是由于χ2检验是定性分析,只涉及物种在样地内存在与否,不涉及物种的数量特征,不能非常准确地描述物种对数间的关联性强弱。因此,本研究只对Spearman秩相关分析结果进行说明。
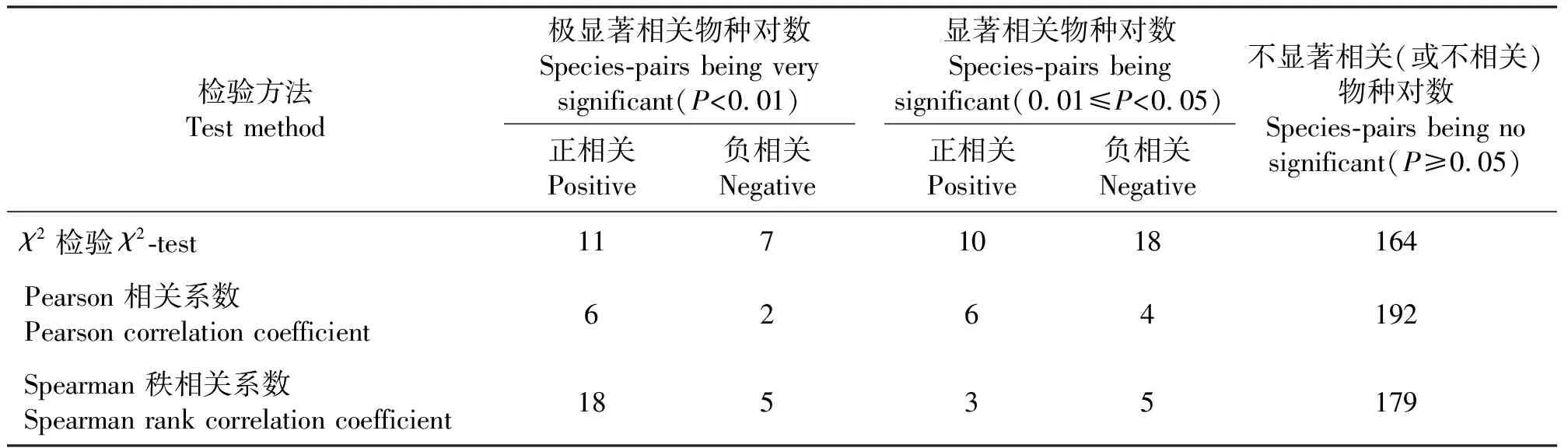
表2 宁夏东部风沙区荒漠草原21种主要植物种间关系的3种检验结果Table 2 Results of interspecific relationship produced by χ2-test, Pearson’s correlation coefficient, Spearman’s rank correlation coefficient, respectively

图1 宁夏东部风沙区荒漠草原植物群落种间关联性的χ2检验半矩阵图Fig.1 The semi-matrix of χ2-test among 21 major species
Spearman秩相关分析呈极显著正相关的物种对数为18个,如紫翅猪毛菜、地锦、狗尾草、尖头叶藜两两物种之间都呈极显著正相关。这是因为这4种植物都为一年生草本,它们之间相互依赖,资源利用方式相似,对生境的适应和反应相同。而物种对数间呈极显著负相关,表明两物种由于空间排挤、竞争,以及不同的资源利用方式,对生境要求不同,出现生态位的分离[10]。
3 讨论
3.1种间关联测定 种间关联测定有多种方法,主要有χ2检验、共同出现百分率法、方差比率法、关联系数、分布相似性百分率法、相关系数、点相关系数等[11]。本研究应用的是χ2检验和Jaccard关联指数两种方法。其中,在χ2检验中呈极显著和显著正关联的物种对数,其JI值均较大。如丝叶山苦荬-狗尾草的JI值是0.41,χ2检验中也呈极显著关联,这说明Jaccard联结指数和χ2检验在反映物种间的关联程度方面较为一致。但是,JI值的运用也有局限性,如赖草-丝叶山苦荬的χ2检验值很小,物种对数间并没有表现出显著关联性,而其JI值是0.41。在测定正、负关联的过程中,赖草-丝叶山苦荬是所有物种对数里唯一一个无关联的物种对,其ad=bc,说明赖草、丝叶山苦荬两个物种随机出现,无关联。同时从χ2检验的数学角度来看,这两个种出现的概率恰与概率统计的期望值相等,说明赖草-丝叶山苦荬间完全独立。以上表明χ2检验方法和Jaccard关联指数应该同时使用才能更准确、更合理地描述和测定种间关联程度。

图2 宁夏东部风沙区荒漠草原植物群落种间关联性的Jaccard关联指数半矩阵图Fig.2 The semi-matrix of Jaccard association index among 21 major plant species

图3 宁夏东部风沙区荒漠草原植物群落种间关联性的Pearson相关系数半矩阵图Fig.3 The semi-matrix of Pearson’s correlation coefficient among 21 major species

图4 宁夏东部风沙区荒漠草原植物群落种间关联性的Spearman秩相关系数半矩阵图Fig.4 The semi-matrix of Spearman’s rank correlation coefficient among 21 species
单独运用JI值的另外一个局限是在χ2检验中呈显著负关联的物种对,JI值没有表现出相应数值较大的特征,这与χ2检验中呈显著正关联物种对的JI值较大的情况不同。这种情况可能是由于Jaccard关联指数公式的分子为a,a表示含有两个种A和种B的样方数,当物种对数间呈正关联时a较大,而当物种对数间为负关联时,a较小,b、c较大,而分母(a+b+c)的值不管是正关联还是负关联,始终变化不大。因此,当物种对数间为正关联时,a较大,所以JI值也较大,可以较好地反映正关联的强弱。而当物种对数间为负关联时,a较小,其JI值也较小,所以,Jaccard联接指数不能较好地反映物种对数间负关联性的显著情况。同时,χ2检验也有其局限性,因为χ2检验使用的是存在与否的二元数据,不能区分关联程度的大小。总体来看,Jaccard联接指数和χ2检验都有助于客观测定两物种种间关联性,一定程度上可以说明物种对数间的相互关系,并表明其关联程度。因此,在实际运用中,要结合两种方法使用才能更好地反映种间关系[12]。
3.2种间相关测定 对于χ2检验定性分析的局限性,Spearman秩相关分析方法可以有效规避这一部分。如短花针茅-紫翅猪毛菜、短花针茅-尖头叶藜、短花针茅-雾冰藜在χ2检验中均为显著负相关关系,而在Spearman秩相关分析结果里并无显著相关关系。这可能是由于短花针茅在χ2检验中使用的二元数据,与Spearman秩相关分析中使用的重要值数据有较大差异,因为不同样地中的重要值数据不相等且数值差距也较大,所以分析过程中所使用数据的差异导致了两种方法出现不同的分析结果。重要值的数量特征更细化、更确切地描述了群落的特征,所以使用Spearman秩相关分析方法可以更合理地对种间相关性进行测定。种间相关可以弥补χ2检验利用二元数据进行分析的不足,尤其是避免了伴生种、偶见物种对种间关系研究的影响[9]。
3.3种间关系与群落演替 对于草地植物群落的种间关系与群落演替之间的关系,很多学者已对此做了研究。刘珏宏等[6]发现内蒙古锡林郭勒草原研究区处于演替中期,未形成总体关联性,呈正相关状态。种间关联较为松散,总体处于相对共存的状态。房飞等[13]对高寒草原植物群落进行研究,发现原始生态种之间多呈正关联,处于退化阶段的物种则表现出无关联或负关联,一定程度上反映了群落演替的逆向过程。在本研究中,物种对数间呈显著、极显著相关的物种对数远低于呈不显著相关的对数,说明该区域植物群落的种间关系松散。物种对数间呈显著正相关或显著负相关,说明群落在长期的演替过程中,各种群之间既相互依存又相互排斥。一部分种群占据适宜的生态位,和谐相处,而另一部分种群独立性较强或进行竞争[8],所以该区域植物群落尚处于不断的发展演替之中。通过对群落种间关系进行研究,可以找出群落内和群落间的联系,有助于认识植物群落的分布规律和动态演替过程,并为区域生态环境建设和合理利用草地资源提供依据。
4 结论
宁夏东部风沙区荒漠草原植物群落的种间关联性较弱,旱生丛状禾草、杂类草和一年生草较多,并有大量沙生植物,群落稳定性低。物种对数间呈负关联物种对数多于呈正关联物种对数,呈极显著和显著相关的物种对数所占比例较小,仅为14.76%(Spearman秩相关分析结果),物种对间竞争水平低,群落结构简单化,群落种间关系松散,物种趋向于独立分布,仍处于群落演替之中。
[1] 王伯荪.植物种群学[M].广州:中山大学出版社,1989:115-120.
[2] 吴征镒.中国植被[M].北京:科学出版社,1980.
[3] 张晶晶,许冬梅.宁夏荒漠草原不同封育年限优势种群的生态位特征[J].草地学报,2013,21(1):73-78.
[4] 程中秋,张克斌,刘建,等.宁夏盐池荒漠草原区天然草地植物生态位研究[J].水土保持研究,2011,18(3):37-40.
[5] 方楷,宋乃平,安慧,等.宁夏盐池荒漠草原植被的数量分类和排序[J].生态学杂志,2011,30(12):2719-2725.
[6] 刘珏宏,高慧,张丽红,等.内蒙古锡林郭勒草原大针茅——克氏针茅群落的种间关联特征分析[J].植物生态学报,2010,34(9):1016-1024.
[7] 李博.生态学[M].北京:高等教育出版社,2000:122-124.
[8] 杜道林,刘玉成,李睿.缙云山亚热带烤树林优势种群间联结性研究[J].植物生态学报,1995,19(2):149-157.
[9] 张峰,上官铁梁.山西翅果油树群落种间关系的数量分析[J].植物生态学报,2000,24(3):351-355.
[10] 张金屯,焦蓉.关帝山神尾沟森林群落木本植物种间联结性与相关性研究[J].植物研究,2003,23(4):458-463.
[11] 彭少麟,周厚诚,郭少聪,等.鼎湖山地带性植被种间关系研究[J].植物学报,1999,41:1239-1244.
[12] 李军玲,张金屯,袁建英.关帝山亚高山灌丛群落和草甸群落优势种的种间关系[J].草地学报,2004,12(2):113-119.
[13] 房飞,胡玉昆,张伟,等.高寒草原植物群落种间关系的数量分析[J].生态学报,2012,32(6):1898-1907.
