天然羊草草地绵羊采食的季节变化
2013-03-13陈文青张英俊
陈文青,张英俊
(中国农业大学动物科技学院,北京 100193)
羊草(Leymuschinensis)草地主要分布于欧亚草原以东,我国境内以松嫩平原和内蒙古高原以东地带为主[1]。羊草由于具有较高的适口性,是理想的放牧和饲草利用草种,并且羊草对生长条件要求较为宽松,使得羊草草地成为较适宜发展我国北方畜牧业的草地类型之一。然而,目前由于载畜量的过量超载已导致羊草草地覆盖度降低,甚至已导致部分地区出现退化和盐渍化现象[2]。一般来讲,放牧家畜在天然草地中的采食都表现出一定的选择性,尤其是在草地植被组成较为丰富且可利用牧草充足时,家畜对牧草的选择性采食表现的尤为强烈[3-4]。家畜的这种选择性采食会导致植物群落间或群落内产生不同的放牧压[5],而且相关研究[6-7]也表明家畜的选择性采食表现出明显的季节性差异,因此, 研究放牧家畜在羊草草地采食的季节性变化,根据家畜不同放牧季采食结构的变化,合理地调整畜群结构、制定不同的放牧制度具有重要的现实意义。
目前,国内对于家畜选择性采食的研究大多还局限于传统采食测定方法,如食道瘘管、显微镜法等,不仅对于家畜正常的采食行为造成了干预而且准确性不高。在国际上,饱和链烷技术已经被成功运用于家畜采食组分的估测[8-11],而且随后引入的长链醇和长链酸进一步完善了该技术[12-15],使得放牧家畜在天然草地中的采食组分测定成为可能。本研究利用饱和链烷与长链醇或长链脂肪酸的组合技术测定了绵羊在天然羊草草地不同月份采食组分的变化,以揭示放牧绵羊的自然食性及不同月份食物替代规律,并探讨了家畜选择性采食与草地群落结构间的关系。
1 材料与方法
1.1试验区概况 试验区位于河北省沽源县国家草地生态系统野外科学观测研究站,地理位置115°39′ E, 41°45′ N,海拔1 430 m。气候为半干旱大陆季风气候,年平均降水量430 mm,降水大多集中于7-9月,试验期间(6、7和8月)月平均气温分别为18.1、21.1和16.4 ℃。 羊草和苔草(Carexduriuscula)为该区优势植物,5月初开始萌发,维持常绿至9月底;猪毛蒿(Artemisiascoparia)、芦苇(Phragmitescommunis)及糙隐子草(Cleistogenessquarrosa)在试验区占有较大比例;针茅(Stipakrylovii)呈现分散分布;其他草种如皱黄芪(Astragalustataricus)、草地风毛菊(Saussureajaponica)、委陵菜(Potentillasupina)、蒲公英(Taraxacummongolicum)和伏毛山莓草(Sibbaldiaadpressa)在草地中所占比例较小。
1.2试验动物及管理 试验初期(放牧处理前3-5天)从当地畜群挑选体质量及身体状况(身体健康)基本一致的蒙古绵羊(母羊)10只,体质量平均(38±2.9) kg,使其在1.5 hm2羊草草地自由采食,放牧季分为3个阶段,分别为6月15日-7月15日、7月16日-8月15日、8月16日-9月15日(放牧季结束大约 50%地上植被被采食)。所有试验绵羊均长期放牧于羊草草地,试验期间水量供应充足。
1.3取样过程
1.3.1草样收集 在每月取样前,首先通过直接观察法观测绵羊采食牧草种类(3 d观测期)。根据绵羊采食牧草种类,取样期在放牧区随机剪取各采食草样。 由于考虑到植物表皮蜡质含量会随季节变化,因此,每月取样一次,然后将所取样品在65 ℃烘箱中烘48 h,过1 mm筛,随后进行饱和链烷、长链醇和长链脂肪酸的分析。
1.3.2粪样收集 在每个试验期的后7 d,利用粪袋连续收集绵羊粪样,收集后立即在65 ℃烘箱中烘48 h,过1 mm筛,然后将每只绵羊7 d的粪样进行均匀混合,取适量代表样进行饱和链烷、长链醇和长链脂肪酸的分析。
1.4饱和链烷、长链醇和长链脂肪酸的提取 粪样和草样中饱和链烷的提取方法按照文献[16-17]的方法。长链醇和长链脂肪酸的提取方法按照文献[18]的方法。具体方法如下:
①皂化:称取0.1 g粪样(草样0.2 g)装进耐热离心玻璃瓶(日本IWAKI玻璃公司),加入1.5 mL(草样为2 mL)1 mol·L-1氢氧化钾醇溶液(ethanolic KOH)。再加入1 mL含有内标液1和内标液2的溶液。内标液1含有饱和链烷n-Docosane(C22)和n-Tetratriacontane(C34)0.02 mg·mL,溶剂为庚烷;内标液2含有长链醇C27和长链脂肪酸C21,浓度分别为0.114和0.117 mg·mL-1,溶剂为庚烷与乙醇的混合液(体积比1∶1)。然后,拧紧瓶盖,放入90 ℃烘箱中烘16 h(过夜)。
②萃取:待样品冷却后加入1.4 mL庚烷(草样加2 mL)和0.6 mL蒸馏水,摇匀并置于50 ℃水浴锅中加热10 min。
③离心:2 000 r·min-1离心5 min。离心后取上清液于新试管中并重新加入1.4 mL 庚烷进行第2次萃取,再取上清液(重复步骤②和③)。之后,将取完上清液的装有样品残渣的玻璃瓶盖上玻璃盖,保存,留待下一步提取长链脂肪酸。
④过滤:称1.5 g硅胶装入漏管(日本IWAKI玻璃公司)中制成过滤装置,向漏管中均匀地加入2 mL 庚烷使硅胶达到饱和状态,然后将2次收集到的上清液过滤,再分别用1、2、1 mL的庚烷冲洗试管并加入到漏管中(确保黄色部分没有流出漏管),全部的过滤液用试管收集,这部分无色的液体为饱和链烷。然后用空气压缩机吹干,将吹干的装有饱和链烷的试管用保鲜膜密封保存。待气谱分析。
⑤配备庚烷和乙酸乙酯体积比为8∶2的混合溶液,取4 mL该溶液冲洗步骤④中用庚烷冲洗过的漏管,用试管收集过滤液,该过滤液即为长链醇(混合部分类固醇物质),然后用空气压缩机吹干,用保鲜膜密封保存。待气谱分析。
⑥往步骤③中装有样品残渣的玻璃瓶内先加入0.25 mL的5.8 mol·L-1盐酸(草样为0.4 mL),再加入2 mL庚烷,摇匀并置于50 ℃水浴锅中加热10 min。2 000 r·min-1离心5 min。离心后取上清液于新试管。
⑦重复步骤⑥。用空气压缩机吹干收集到的上清液。再溶于1 mL的庚烷中,水浴加热确保完全溶化。
⑧称1.5 g硅胶装入漏管中制成过滤装置,向漏管中均匀的加入3 mL乙酸乙酯和庚烷的混合液(体积比为8∶92),润洗硅胶,使硅胶达到饱和状态(这个时候不收集滤出液)。然后取0.5 mL步骤⑦中溶液加入漏管过滤,再用4 mL乙酸乙酯和庚烷混合液(体积比为30∶70)冲洗,试管收集。吹干,再溶于0.5 mL甲苯,然后添加1 mL硫酸甲醇溶液(20 mL浓硫酸溶于1 L甲醇)。用橡皮塞密封试管,加透明胶带固定。在50 ℃的烘箱中过夜。取出摇匀,先后加入1 mL 0.5 mol·L-1碳酸钾和1 mL庚烷,移出上清液于试管中,再加1 mL庚烷,再取上清液于试管中。吹干保存。
⑨定容:饱和链烷和长链醇,向吹干的干燥物中加入0.5 mL 庚烷,水浴摇匀使干燥物完全溶解,将溶液倒入气相色谱专用的玻璃瓶中封盖,上气谱分析。长链脂肪酸,向吹干的干燥物中加入0.5 mL 十二烷(dodecane),水浴摇匀使干燥物完全溶解,将溶液倒入气相色谱专用的玻璃瓶中封盖,上气谱分析。
所用仪器为日本岛津GC-2010配毛细管分流进样系统,AOC-20i自动液体进样器,FID (flame inoization detector)检测器,色谱柱为TC-1型毛细柱(30 m×0.25 mm×0.25 μm)。
1.5饱和链烷、长链醇和长链脂肪酸回收率的测定 在利用饱和链烷技术估测采食组分中,为获得较为准确的估测结果必须对指示剂回收率进行测定。根据直接观察法记录的家畜采食组分,将从试验区附近相似区域收获的各采食组分按一定比例饲喂6只舍饲绵羊。每天8:00和16:00分别进行饲草投喂(饲喂期7 d),每只绵羊饲喂量根据预饲期(6 d)的量进行调整,确保每只绵羊饲草剩余量<5%。依据收集的各绵羊全粪量和投喂的饲草总量计算饱和链烷(C23~C33)、长链醇(C20~C32)及长链脂肪酸(C20~C32)的回收率。回收率计算公式为:
式中,n为各指示剂回收率,Cfi为粪样中某指示剂浓度,Mf为粪样干物质质量,Cpi为投喂饲草中的某指示剂浓度,Mp为投喂饲草的干物质质量。
1.6草地植被结构和采食组分 在收集草样、粪样期间同时测定草地地上各草种干物质质量、植被高度以及相对频度。在试验区随机剪取12个1 m×0.5 m的样方(每个样方内草种分类剪取),65 ℃烘箱烘72 h后称量,计算各物种的地上干物质比。植被高度通过利用“HFRO sward-stick”在试验区随机测定各采食草种高度100次获得。频度的测定是通过利用0.1 m2的样圆,在样地沿随机方向跑出50~100次,计算每一草种在该投掷次数中出现的百分数。采食组分是根据植物样和粪样中饱和链烷与长链醇或长链酸的适当组合,利用专门软件‘Eatwhat’根据非负最小二乘法计算采食成分(即在日粮组成比较简单的情况下,采用联立方程组的方式估算出日粮组成。这时选作指示剂的饱和链烷的数目要与日粮组分的数目相同。当选作指示剂的饱和链烷的数目大于日粮组分的数目时,便要采用最小二乘法来计算,因为最小二乘法可以从几组答案中选出一个最优结果),粪样中饱和链烷、长链醇和长链脂肪酸的浓度均利用舍饲6只绵羊的平均回收率进行校正。校正方法为自由放牧条件下绵羊粪样中指示剂的浓度除以各指示剂粪样中的回收率。
1.7数据处理 数据处理软件采用 SAS 8.2。利用主成分分析法(PCA)分析植物间饱和链烷与长链醇组合,饱和链烷与脂肪酸组合,长链醇与脂肪酸组合及饱和链烷、长链醇和脂肪酸三者组合模式之间的差异,以确定最佳指示剂组合。统计分析中单只绵羊作为重复,不同月份间采食组分之间差异性分析通过proc GLM进行,所有采食组分在进行方差分析前均进行了三角转换。对粪样中饱和链烷、长链醇和长链脂肪酸浓度进行主成分分析,进一步分析家畜在不同月份之间的采食差异,并对第一、第二主成分进行了方差分析。家畜择食指数的计算利用以下公式[19]:
式中,D为择食系数,ri为家畜对某植物组分的采食比例,pi为该组分在草地中所占的比例。
2 结果与分析
2.1指示剂回收率 绵羊粪便中饱和链烷、长链醇和长链脂肪酸的回收率如表1所示。
2.2植被的季节变化 羊草、苔草和芦苇在6月均具有较高的垂直分布,7月达到最高,8月出现不同程度的下降;碱蓬虽然具有相同变化趋势,但其垂直分布较前三者低;猪毛蒿和旋复花的垂直分布在放牧期间基本维持稳定;蒲公英在6月的垂直分布最高,7―8月明显下降;滨藜的垂直分布随季节不断增加。植被的水平分布变化也较为复杂,羊草和苔草水平分布随季节迁移不断下降,但仍然维持在较高水平;猪毛蒿水平分布的变化与羊草和苔草具有相同趋势,但在各月份的分布明显处于较低水平;芦苇、碱蓬和蒲公英的水平分布持续增加;滨藜和旋复花的水平分布也较低而且呈现相反的变化模式(图1)。
植物地上组分(地上干物质比)是水平分布和垂直分布的综合体现,其季节变化如表2所示,羊草在整个放牧季所占比例明显高于其他草种(分别为40.61%、55.86%和37.46%);苔草在6和7月(分别为22.25%和12.17%)是除羊草以外的第二大主要组分,但在8月(8.32%)出现大幅下降;碱蓬在6和8月(分别为13.68%和19.67%)所占比例较大,7月的组成比例(6.50%)较低;芦苇在整个放牧季所占比例均不大,分别占到草地比例的3.98%、4.01%和11.32%;其他组分如滨藜、猪毛蒿、旋复花在草地中组成比例较小,为草地植被小组分植物。
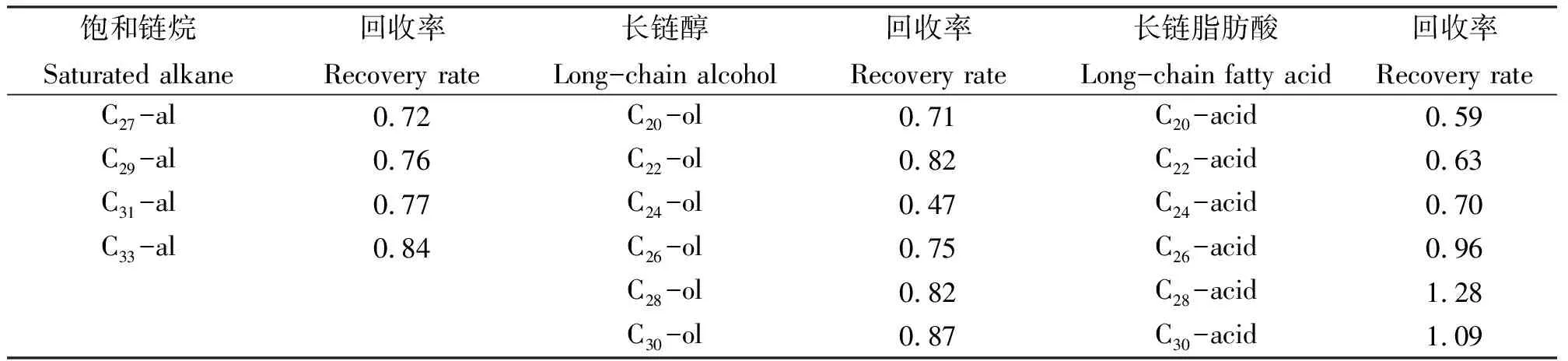
表1 绵羊粪便中饱和链烷、长链醇和长链脂肪酸的回收率Table 1 Recovery rates of saturated alkane, long-chain alcohols and fatty acids in sheep faeces
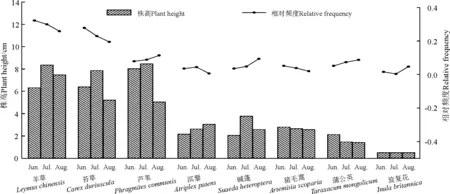
图1 放牧季试验区植被结构变化Fig.1 Structure of vegetation at experimental site during the grazing period
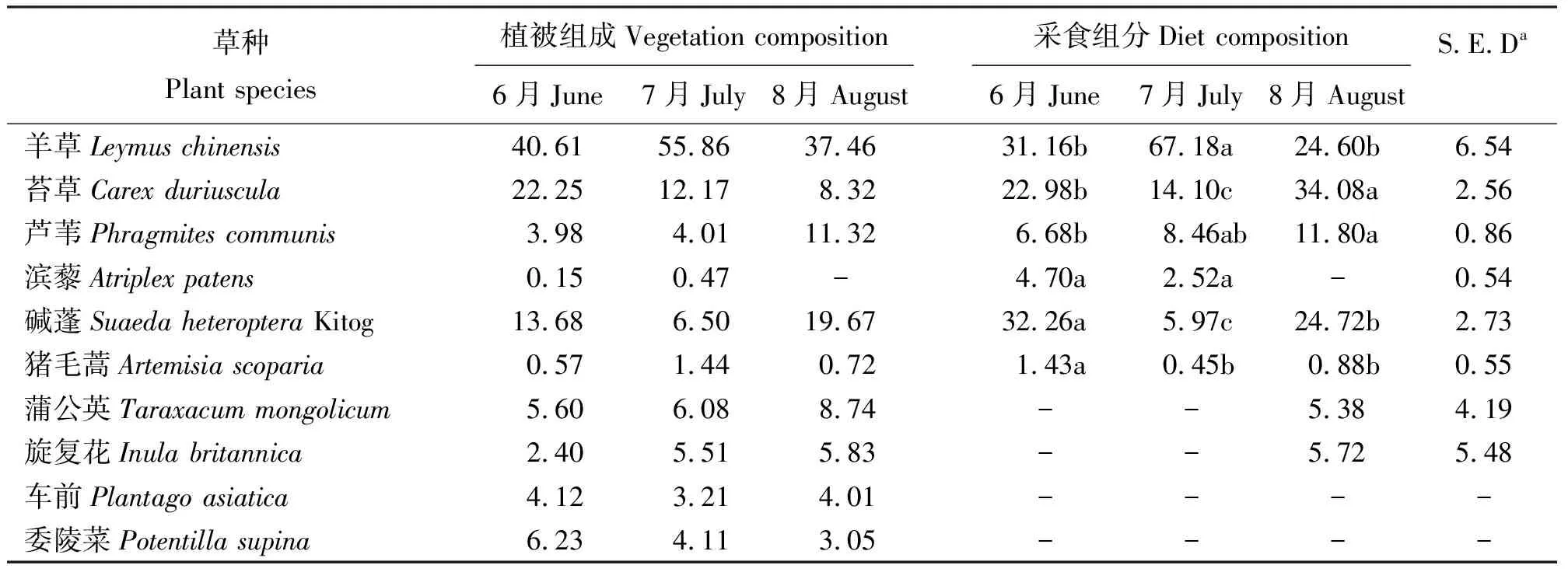
表2 季节对家畜采食组分的影响Table 2 Effect of different months on the diet composition of sheep grazing on Leymus chinensis grassland %
2.3采食组分的变化 对粪样中饱和链烷、长链醇和长链脂肪酸浓度n-alkane+alcohol+acid 进行主成分分析,结果显示不同放牧季3种指示剂出现明显的区分(图2)。在6、7、8月第1主成分(PC1)和第2主成分(PC2)所解释的变异量分别占总变异量的69.26%、78.19%和75.15%,6、7、8月的标准差分别为1.250、1.313和1.021。进一步对PC1和PC2进行方差分析,发现6、7、8月PC1和PC2均差异显著(P<0.05)。
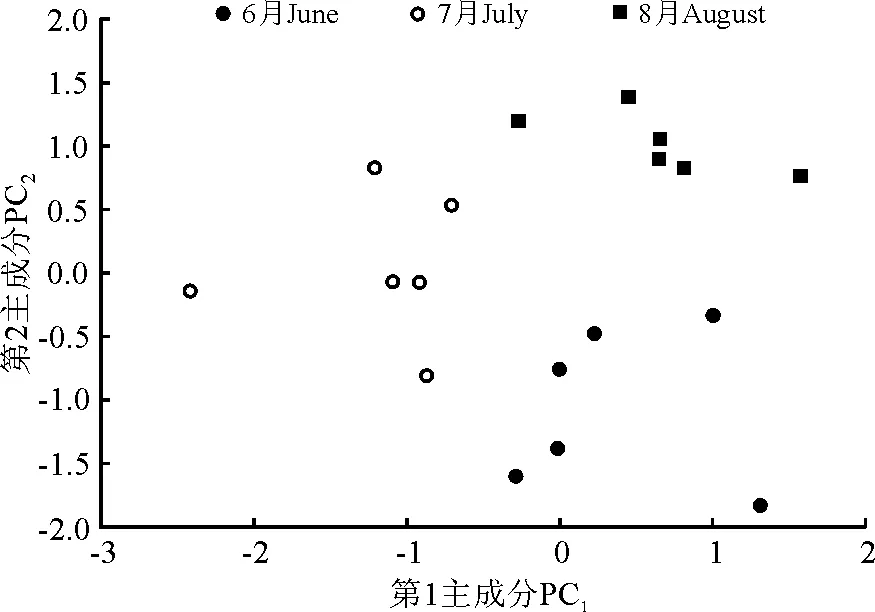
图2 6、7、8月绵羊粪样中饱和链烷、长链醇和长链脂肪酸的主成分分析Fig.2 The principal component analysis of saturated alkanes,long-chain alcohol and fatty acids in sheep faeces in June, July and August
主成分分析显示绵羊在不同季节所采食的植物种类是有差别的,从而导致摄入的饱和链烷、长链醇和长链脂肪酸浓度之间也存在差异。同时,图2中所示的同一季节相对较为分散的点说明绵羊个体之间的采食存在一定程度的差异。
通过对不同月份家畜采食组分的分析,发现家畜采食组分存在明显的季节性差异(表2)。6月,碱蓬是主要的采食组分,其次是羊草和苔草,三者已占该月总采食比例的86.40%;芦苇、滨藜和猪毛蒿采食比例较小,分别为6.68%、4.70%和1.43%。7月,绵羊对羊草的采食出现大幅增加,达到67.18%,而对苔草和碱蓬的采食则急剧下降为14.10%和5.97%;对芦苇的采食有上升趋势但与6月无显著差异;7月绵羊对滨藜和猪毛蒿的采食均出现不同程度的下降。在6、7月均未观测到绵羊对蒲公英和旋复花的采食。8月,绵羊对羊草的采食比例发生大幅下降,仅占总采食比例的24.60%,显著低于7月,但与6月无显著差异;对苔草的采食大幅上升为34.08%,成为该月最主要的采食组分,显著高于前两个月;碱蓬是仅次于苔草的第二大采食组分,相比7月采食比例显著增加,但仍然显著低于6月;对芦苇的采食虽然与7月无显著差异,但绵羊对其采食继续维持增加趋势;8月猪毛蒿仍然是微量采食组分,与7月相比无显著变化,但显著低于6月;8月家畜不再采食滨藜,却首次观测到对蒲公英和旋复花的采食,尽管对两者的采食比例并不高。
2.4择食指数 本试验中所用的择食指数代表家畜采食某种植物的比例与该植物在草地中相对现存量之间的关系。在整个放牧季,家畜对各植物的择食指数具有明显的季节差异,而且对某一植物组分的择食指数没有持续的一致性,其中正值代表家畜采食某植物的比例大于该植物在草地中所占的比例,负值则代表家畜采食某植物的比例小于该植物在草地中所占的比例(图3)。6月,家畜对羊草的采食比例较大,但其择食指数表现为避食性;同样,苔草作为6月主要采食组分其择食指数仅仅表现为轻微喜食,其采食比例与植被在草地中的比例大致相同;虽然对芦苇采食比例较小却对其表现出一定程度的喜食性;对碱蓬和滨藜表现出极强的喜食性;猪毛蒿、蒲公英以及旋复花在6月均表现出较强的避食性。7月,家畜对植物的择食指数发生一定程度的变化,羊草由6月的避食转变为喜食;对苔草的喜食程度进一步增强;芦苇由原来的轻微喜食转变为较强喜食;滨藜在7月继续维持极强的喜食性,而且家畜对其喜食性较6月进一步增强;相反,家畜对碱蓬的喜食性在7月大大降低,仅为微弱喜食;7月家畜对猪毛蒿、蒲公英和旋复花仍然表现出较强的避食性。8月,家畜对植物的择食指数再次发生变化。羊草再次转变为避食,而且避食程度相比6月有所增强;对苔草的喜食性出现大幅增强,表现为极强喜食;对芦苇喜食性降至放牧季最低;而对碱蓬的喜食性与7月差异不大;对猪毛蒿、蒲公英和旋复花仍然表现出较强的避食性,但对蒲公英和旋复花的避食程度较6、7月有明显降低。
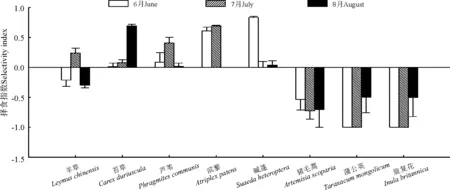
图3 6、7、8月绵羊采食羊草草地对各植物的择食指数Fig.3 Selectivity indices for sheep grazing on Leymus chinensis grassland in June, July and August
3 讨论与结论
6-8月,家畜采食组分具有较大差异。从草地植被结构的季节性变化方面来考虑,家畜采食组分发生变化也是合理的。有关家畜选择性采食[20-21]的研究表明家畜对某植物的采食与其垂直和水平分布密切相关。植被垂直分布主要影响到家畜每口啃食量[22-23];水平分布则影响到家畜遇到相应采食组分的频率,进而影响到其采食率[24-25]。6月,羊草、苔草和碱蓬是该月主要的采食组分,由于该月羊草、苔草无论在水平分布还是垂直分布方面较其他草种要高,因此对两者采食量较大;对碱蓬而言,虽然在6月水平分布和垂直分布均较低,但仍然成为该月采食比例最高的组分,主要是因为碱蓬幼嫩多汁,适口性较好,而且也有利于家畜补充水分,这些特征均有利于家畜的采食。赵钢等[26]也发现家畜倾向于选择性采食那些幼嫩、多汁、营养价值高的植物种类。滨藜虽然也具有幼嫩、多汁的特性,但因其在草地中的组成比例较小,所以导致其采食比例较低。同样,其他草种如芦苇、猪毛蒿、旋复花等由于较低的水平分布和垂直分布,对家畜来说较难获取,家畜很少采食或不采食。7月,家畜对羊草采食明显增加,主要是因为在该月羊草的垂直分布进一步增加,尽管其水平分布有所下降但其仍然维持在较高水平并未限制到家畜对其采食;对苔草的采食出现一定程度的下降,或许是因为羊草的更易获取在一定程度上限制了对苔草的采食,另外,羊草较苔草有好的适口性也是影响其采食的重要因素。 由于在6月家畜对碱蓬的高强度采食使得碱蓬地上生物量大幅下降,从而导致在7月采食较少。家畜对芦苇的采食有一定程度的增加,主要是因为7月芦苇的垂直分布和水平分布均有增加,使其地上组成比例相应提高,增加了家畜采食机会;7月滨藜、猪毛蒿、蒲公英等无论是垂直分布还是水平分布都较低,从而在家畜采食比例中维持较低水平。8月,家畜采食组分发生进一步变化,家畜采食倾向于更加多样化,苔草成为该月采食比例最高的植物,而对羊草的采食出现大幅回落;碱蓬再次成为绵羊重要的采食组分;而对芦苇的采食继续维持上升趋势。通过分析草地植被状况可以看出,8月羊草无论是在垂直分布还是水平分布均出现一定程度的下降,而且由于该月羊草纤维含量明显上升,使得家畜对羊草采食比例出现大幅度的下降,而转向适口性较好的苔草;碱蓬尽管在垂直分布方面有所下降,但其水平分布明显增加,使其地上组成比例大大增加,从而使得家畜对其采食比例显著上升。与此相似,芦苇在8月垂直分布方面有所下降,但水平分布有明显增加,地上组成比例达到放牧季最高,从而也使得家畜对其采食比例达到最高水平。在8月首次观测到家畜对蒲公英和旋复花的采食,尽管两者在垂直分布方面变化不大,但水平分布增加,增大了家畜遇到的频率,使得对两者的采食成为可能。
通过对择食指数的分析来看,家畜采食比例较高的植物择食指数并不高,甚至在有的情况下为避食,如羊草;采食较少的植物反而择食指数较高,如芦苇、滨藜。本研究中,择食指数的计算涉及到家畜采食比例及草地植被组成比例,因此,家畜对某种植物的采食口数较多或采食比例较大并不意味着其相对于草地组成比例较大。另外,择食指数的这种表示方法可以较好地反映家畜采食对草地植被利用的影响。例如,6月,碱蓬和滨藜处于过度采食状态,7月为芦苇和滨藜,8月转向苔草,各月份的其他采食组分则均处于轻度采食或适度采食状态。因此,放牧管理者在不同的放牧季,可以根据家畜对植物择食指数的变化采取相应管理措施,促进某些植被的生长,以实现草畜平衡,在维持草地长久持续可利用的前题下实现畜产品的最大产出。
综上所述,绵羊选择性采食的季节变化与草地植被的垂直分布和水平分布密切相关,家畜对草地植被的选择性采食会影响到植被的分布,同时草地植被分布的变化也会反过来影响绵羊的采食,并且草地植被的适口性在一定程度上也会对绵羊的采食产生一定程度的影响。择食指数是衡量绵羊采食以及绵羊对草地植被影响的重要指标,通过对不同季节绵羊择食指数的测定可以为合理放牧制度的制定提供可靠依据。
[1] Xiao X M,Wang Y F,Jiang S,etal.Interannual variation in the climate and above-ground biomass ofLeymuschinensissteppe andStipagrandissteppe in the Xilin river basin,Inner Mongolia,China[J].Journal of Arid Environments,1995,31:283-299.
[2] Li C H,Zheng X F,Zhao K Y,etal.Basic types of pasture vegetation in the Songnen plain[A].In:Proceedings of XIV international Grassland Congess[C].Lexington,Kentucky,USA.1:Westview Press,1983:432-434.
[3] Beck J L,Peek J M.Diet composition,forage selection,and potential for forage competition among elk,deer,and livestock on aspen-sagebrish summer range[J].Rangeland Ecology and Management,2005,58:135-47.
[4] Burns J C,Sollenberger L E.Grazing behavior of ruminants and daily performance from warm-season grasses[J].Crop Science,2002,42:873-881.
[5] Hunter R F.Hill sheep and their pasture——A study of sheep-grazing in southeast scotland[J].Journal of Ecology,1962,50:651-680.
[6] Fraser M D,Theobald V J,Moorby J M.Determining diet composition on complex swards using n-alkanes and long-chain fatty alcohols[J].Ecological Applications,2006,16:1901-1910.
[7] Narvaez N,Pittroff W.Use of N-alkanes to estimate seasonal intake,digestibility,and diet composition of small ruminants grazing in California chaparral[J].Journal of Animal Science,2007,85:149-152.
[8] Dove H,Mayes R W.The use of plant wax alkanes as marker substances in studies of the nutrition of herbivores:a review[J].Australian Journal of Agricultural Research,1991,42,912-952.
[9] Hameleers A,Mayes R W.The use of n-alkanes to estimate herbage intake and diet composition by dairy cows offered a perennial ryegrass white clover mixture[J].Grass and Forage Science,1998,53:164-169.
[10] Duncan A J,Mayes R W,Lamb C S,etal.The use of naturally occurring and artificially applied n-alkanes as markers for estimation of short-term diet composition and intake in sheep[J].Journal of Agricultural Science,1999,132:233-246.
[11] Osoro K,Olivan M,Celaya R,etal.The effect of Calluna vulgaris cover on the performance and intake of ewes grazing hill pastures in northern Spain[J].Grass and Forage Science,2000,55:300-308.
[12] Dove H,Mayes R W.Plant wax components:A new approach to estimating intake and diet composition in herbivores[J].Journal of Nutrition,1996,126:13-26.
[13] Dove H,Mayes R W.Using n-alkanes and other plant wax components to estimate intake,digestibility and diet composition of grazing/browsing sheep and goats[J].Small Ruminant Research,2005,59:123-139.
[14] Lin L J.The potential use of long-chain alcohols and fatty acids as diet composition markers:factors influencing faecal recovery rates and diet composition estimates in sheep[J].Animal,2009,3:1605-1612.
[15] Mayes R W,Dove H.Measurement of dietary nutrient intake in free-ranging mammalian herbivores[J].Nutrition Research Reviews,2000,13:107-138.
[16] Mayes R W,Lamb C S,Colgrove P M.The use of dosed and herbage n-alkanes as markers for the determination of herbage intake[J].Journal of Agricultural Science,1986,107:161-170.
[17] Olivan M,Osoro K.Effect of temperature on alkane extraction from faeces and herbage[J].Journal of Agricultural Science,1999,132:305-312.
[18] Lin L,Luo H L,Wang H,etal.Evaluation of long-chain alcohols and fatty acids,in combination with alkanes,as markers in the estimation of the composition of four herbages in mixtures[J].Grass and Forage Science,2009,64:19-25.
[19] Jacobs J.Quantitative measurement of food selection-modification of forage ratio and ivlevselectivity index[J].Oecologia,1974,14:413-417.
[20] Penning P D,Parsons A J,Orr R J,etal.Intake and behavior responses by sheep to changes in sward characteristics under continuous stocking[J].Grass and Forage Science,1991,46:15-28.
[21] Dumont B,Meuret M,Prudhon M.Direct observation of biting for studying grazing behavior of goats and llamas on garrigue rangelands[J].Small Ruminant Research,1995,16:27-35.
[22] Edwards G R,Parsons A J,Penning P D,etal.Relationship between vegetation state and bite dimensions of sheep grazing contrasting plant species and its implications for intake rate and diet selection[J].Grass and Forage Science,1995,50:378-388.
[23] Gross J E,Shipley L A,Hobbs N T,etal.Functional-response of herbivores in food-concentrated patches-tests of a mechanistic model[J].Ecology,1993,74:778-791.
[24] Stephens D W,Krebs J R.Foraging Theory[M].New Jersey:Princeton University Press,1986:1-247.
[25] Thornley,J H M,Parsons A J,Newman J,etal.A cost-benefit model of grazing intake and diet selection in a 2-species temperate grassland sward[J].Functional Ecology,1994,8:5-16.
[26] 赵钢,许志信,李德新.沙地放牧系统反刍家畜选择性采食行为的研究[J].内蒙古大学学报,2006,37(3):311-316.
