不同海拔关山草原群落的结构特征
2013-03-13胡远彬
胡远彬,郭 婷,陈 俊
(西北农林科技大学动物科技学院,陕西 杨凌 712100)
草地资源作为一种可再生的农业自然资源,在发展畜牧业、维持生物多样性、保持水土和维护生态系统平衡方面有着重大作用[1]。目前草地退化已成为全世界草地所面临的普遍问题而受到广泛关注,我国90%的草地处于不同程度的退化状态[2],虽然采取了“退耕还林”、“退牧还草”等重大举措,但远不足以遏制草原急速退化的趋势[3]。草地退化是由人为活动和不利自然因素等引起的[4],其中前者是草地退化的主要原因[5]。这些因素的干扰可引起空间异质性,影响植物种群结构及物种多样性[6-7],从而直接影响生态系统的演变过程[8]。空间异质性是生态系统的一个主要属性,也是产生空间格局的主要原因[9],异质性分析已成为生态学者研究不同尺度生态系统功能和过程的重要手段[10],可用于确定人类活动或自然事件对生态格局的影响范围,对受损生态系统恢复和重建给予理论指导[11]。因此,对草地生态系统空间异质性的定量研究显得尤为重要。
生态旅游是一种发展迅速的新兴旅游形式,是21世纪一个极为重要的旅游经济增长点[12]。关山草原地处关山山脉腹地,是陕西所拥有的唯一成片山地草原,拥有独特的旅游资源和旅游环境,但近年来由于不合理的开发和过度利用,使草原退化日益加剧。因此,本研究以不同海拔梯度下关山高山草甸草原作为研究对象,用样方法调查植物种类组成及其出现次数,通过幂乘方法则进行数据解析,探索不同海拔梯度下草地的物种数、物种多样性及空间分布的基本规律,为草地的科学管理及合理利用、退化草地的恢复提供可靠的依据。
1 材料与方法
1.1调查地概况 调查样区位于陕西省陇县西南部关山草原风景区(106°34′23″ E,34°43′17″ N),属于高山草甸草原,景区为石质山地,黄土覆盖较薄,海拔高度差异较大,平均在2 100 m左右,谷坡较缓,多在20°~35°。年均气温5.4 ℃,月均最高、最低气温分别为17 ℃和-10 ℃,≥10 ℃年积温不足2 000 ℃·d,年均降水量650~700 mm,气候湿润,拥有天然草地2.3万hm2,天然林6.9万hm2。
1.2调查时间和方法 野外调查于2010年7月2日至5日在关山草原进行,所选草地为阳坡,坡度约45°。在同一坡面的坡下(海拔2 092 m)、坡中(海拔2 112 m)、坡上(海拔2 132 m)草地群落中分别拉一条长40 m的样线,每条样线连续摆放80个50 cm×50 cm的铁制样方框,称之为大样方(以下简称L样方),将L样方等分成4个25 cm×25 cm的小样方(简称S样方)。调查时只统计草地群落内各植物种在L样方内的出现次数,如果某植物种在L样方内的4个S样方中都没有出现,为0次,只在1个S样方内出现记作1次,以此类推,4个S样方中都出现则记作4次[13-14]。此调查方法简单,对草地没有破坏性。
1.3数据解析方法
1.3.1幂乘方法则 本研究的数据解析运用幂乘方法则进行[15-16],幂乘方法则就是分别把xi和yi作为横坐标与纵坐标,对群落内出现的所有植物种作散点图时,yi可以用xi的线性回归方程式yi=α+βxi+εi(α、β为回归系数,εi为植物种i的回归残差)来表示的经验法则。
其中:
xi=log[Pi(1-Pi)/n],
yi=log(vi/n2) .
式中,Pi为植物种i在S样方内出现的频率;n为L样方内的S样方数,本调查中,n=4;Pi(1-Pi)/n为植物种i在随机分布时出现频率的方差;vi为植物种i在L样方内出现次数的方差;vi/n2则为植物种i实际出现频率的方差。
1.3.2植物群落中各植物种的空间异质性指数 群落中各植物种空间异质性程度的大小,用异质性指数δi来表示[15-17]:
δi=log(vi/n2)- log[Pi(1-Pi)/n] .
各植物种的空间异质性程度,通过计算δi值来判定:若δi=0,则植物种处于随机分布状态;δi>0时,植物种比随机分布具有更高的空间异质性(即团块分布或集中分布);δi<0时,植物种i具有比随机分布小的空间异质性(即规则性分布或均匀分布)。
1.3.3植物群落整体的空间异质性指数 植物群落整体的空间异质性指数[14-15,17-19],是在综合考虑组成群落的所有植物种的出现频率及其对它们异质性程度的大小进行加权平均得出的:
式中,s为群落内出现的物种数,Pi、δi分别为植物种i的出现频率及异质性指数。δc值对空间异质性程度的判断基准与δi值相同。
1.3.4物种多样性指数及均匀度指数 多样性指数采用了Shannon-Wiener的H′指数,用下式计算:
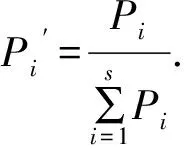
均匀度指数采用了Pielou[20]的均匀度指数(J′):
式中,s为群落内出现的物种数。
2 结果与分析
2.1幂乘方法则的吻合性 坡下、坡中和坡上草地群落中,以组成群落的各个植物种随机分布时出现频率方差的对数值{xi=log[Pi(1-Pi)/n]}为横坐标,以它们实际出现频率方差的对数值[yi=log(vi/n2)]为纵坐标作散点图时,得到回归直线的决定系数R2值都在0.96以上,表明3个不同海拔梯度下植物群落中各植物种及其植物群落整体均与幂乘方法则非常吻合,且幂乘方法则的回归直线整体上都位于y=x直线的上方,表明不同海拔梯度下植物群落整体都具有比随机分布更高的空间异质性(图1)。因此,随后研究中关于群落空间异质性的数据解析将利用幂乘方法则进行。
2.2植物群落的整体特征随海拔梯度的变化 在3个不同的海拔梯度下,各自的20 m2(0.5 m×0.5 m×80)调查样地内,坡上出现的总物种数有66种,比坡中多8种,比坡下多17种;物种多样性指数(H′)随海拔的升高而增加,坡下(3.278)<坡中(3.450)<坡上(3.510);L样方内的平均物种数、平均物种多样性指数均表现为坡下<坡中<坡上,但它们之间的差异不显著 (P>0.05);均匀度指数(J′)随海拔梯度的升高变化不明显;群落整体的空间异质性指数(δc)表现为坡下(0.237)<坡中(0.260)<坡上(0.311)的趋势(表1)。
2.3不同海拔梯度下群落中出现的所有植物种及其空间异质性指数 坡下草地群落中,蛇莓(Duchesneaindica)、毛茛(Ranunculusjaponicus)的出现频率非常高(大于98%),成为第1、第2主要的优势种,二者是草地退化的标志性植物;坡中最主要的优势种为紫羊茅(Festucarubra)、火绒草(Leontopodiumleontopodioides),其中紫羊茅的出现频率为100%,火绒草也是草地退化的标志性植物,蛇莓、毛茛的出现频率大幅度下降;坡上草地中,第1、第2主要的优势种为紫羊茅和膨囊苔草(Carexlehmanii),其出现频率在93%以上,同时第3优势种豆科的天蓝苜蓿(Medicagolupulina)出现频率高于坡中和坡下,蛇莓的出现频率明显低于坡中、坡下,毛茛已经不属于优势种之列,禾本科的雀麦(Bromusjaponicus)出现频率也超过了50%(图2)。结果表明,随着海拔梯度的升高,植物群落的主要物种组成发生了变化,禾本科、豆科植物种的出现频率呈上升趋势,退化草地的标志性植物明显降低,即草地的退化趋势随海拔的升高而减弱。
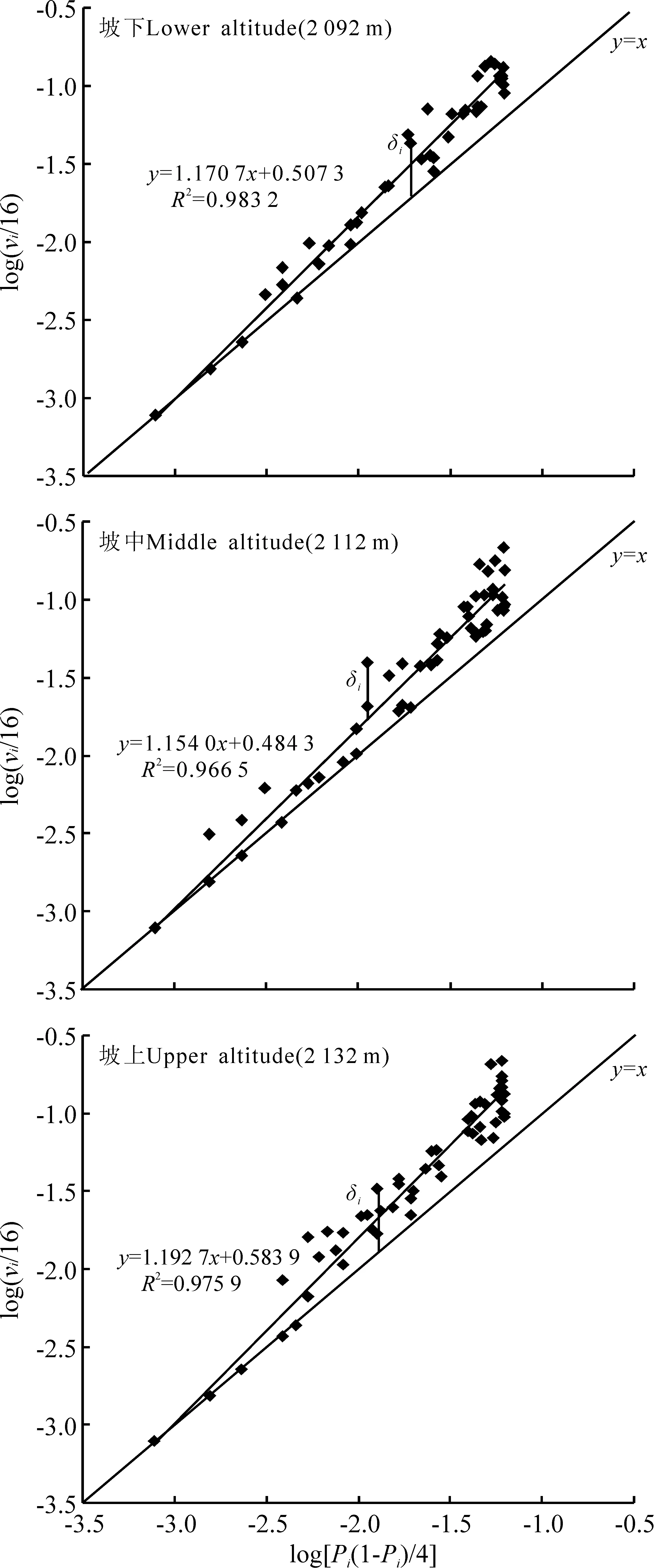
图1 不同海拔梯度下植物种及群落整体对幂乘方法则的吻合性Fig.1 An application of power law to constructing species and entire community at different altitudes
坡下草地中,紫羊茅、膨囊苔草、莎草(Cyperussp.)、火绒草、獐牙菜(Swertiabimaculata)和小花草玉梅(Anemonerivularis)6种植物的出现频率大于50%,且空间异质性较高(大于0.25),它们对于提高群落整体的空间异质性发挥了较大的作用;蛇莓、披碱草(Elymusdahuricus)、毛茛、天蓝苜蓿、龙牙草(Agrimoniapilosa)、小米草(Euphrasiapectinata)和卷耳(Cerastiumarvense)7种植物,出现频率也在50%以上,但空间异质性指数较低(其中蛇莓的出现频率高达99%,异质性指数趋于0),它们大大降低了群落整体的异质性。坡中草地,能大幅度提高群落整体空间异质性指数的有披碱草、蛇莓、膨囊苔草、委陵菜(Potentillachinensis)和毛茛5种植物(出现频率及异质性指数均较高);降低群落整体空间异质性的植物种较坡下少,有紫羊茅、小米草、火绒草和天蓝苜蓿4种(出现频率较高且异质性指数较低)。坡上,天蓝苜蓿、小米草、委陵菜、蛇莓和雀麦5种植物的出现频率及其异质性指数都很高,在很大程度上提高了群落整体的异质性;同时出现频率较高的膨囊苔草、披碱草、铁杆蒿(Artemisiasacrorum)和一年蓬(Erigeronannuus)4种植物降低了群落整体的异质性。
另外,坡下、坡中、坡上草地群落中,出现频率在20%以下的植物都较多,虽然它们的异质性有高有低,但对群落整体的影响较小。出现频率在20%~50%的植物种群,它们异质性程度的高、低在一定程度上提高或降低了群落整体的异质性。
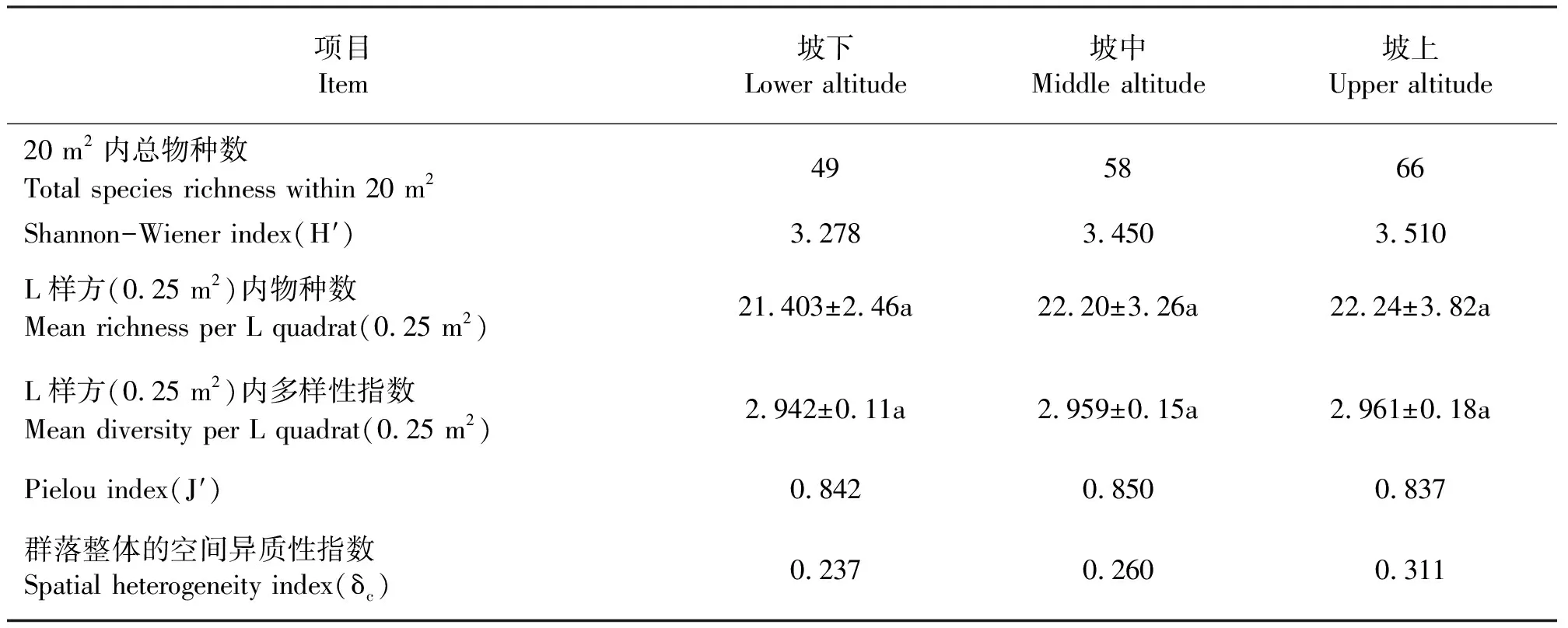
表1 群落总体特征Table 1 Characteristics of communities
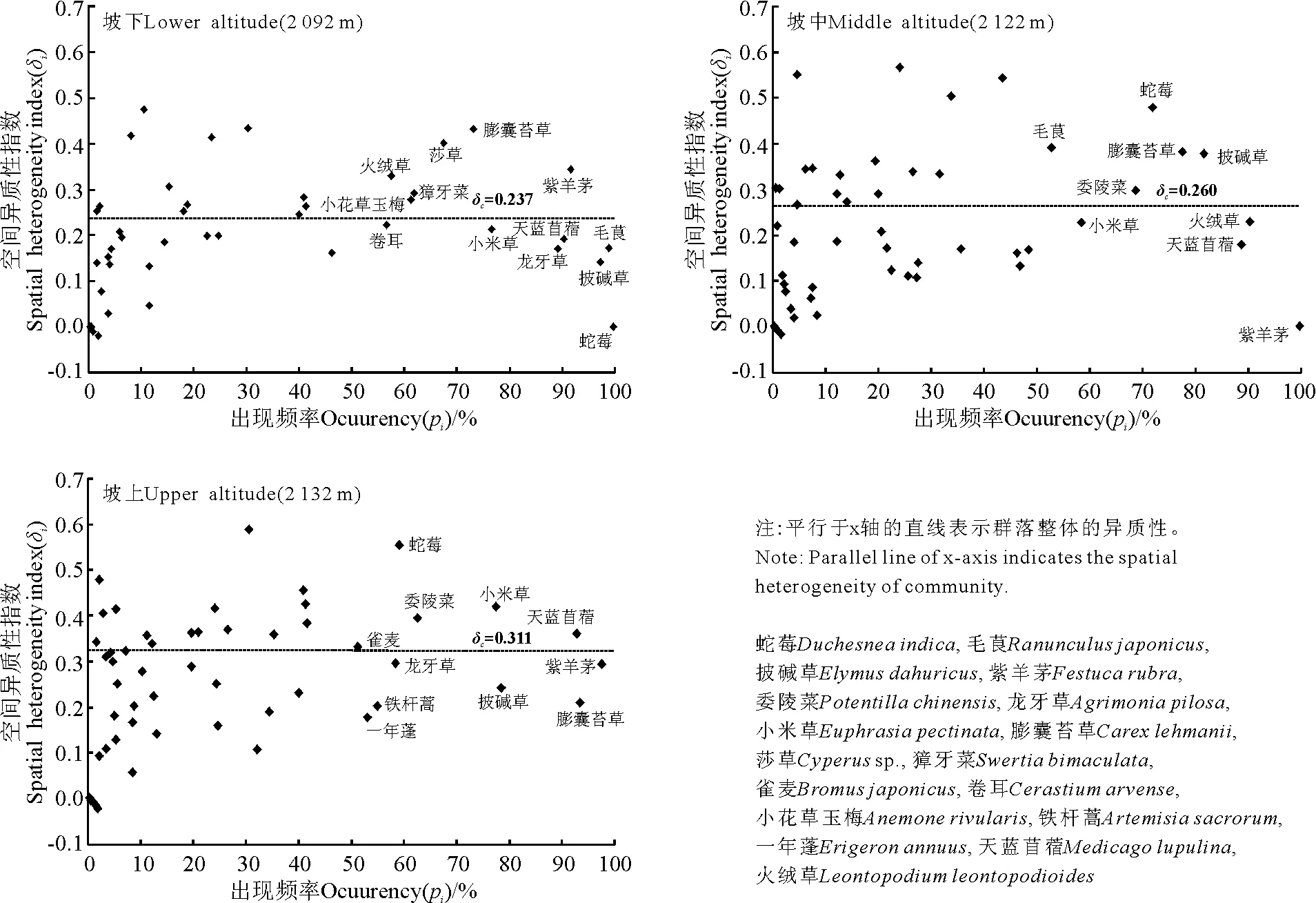
图2 出现频率(Pi)及其空间异质性指数(δi)Fig.2 Occurrence (Pi) and spatial heterogeneity index (δi)
3 讨论与结论
本研究将幂乘方法则与野外植被调查方法相结合,来解析关山草原不同海拔梯度下植被群落的物种组成、物种多样性及其空间分布特性。幂乘方法则早在1960年由昆虫学家Taylor博士在研究昆虫种群分布中发现,现仍然在很多昆虫生态学研究中被应用。1992年,通过Ohio州立大学的Madden和Edinburgh大学的Hudges的研究,将其应用于植物病理学中植物的“发病”与“健康”数据的解析。在日本,大量的研究[14-16]证实该方法可以在草地植被分析中应用。植被调查方法对草地没有破坏性而且省时省力,并且从幂乘方法则的回归直线图形中能够很直观地看出群落中各植物种及群落整体的空间异质性程度[21]。Chen等[17]对内蒙古5个不同景观(依据水分条件进行划分)植被的空间异质性和多样性进行研究表明,幂乘方法则可用于景观植被的分析。吕杰和陈俊[22]利用幂乘方法则对不同利用方式下额尔古纳草原群落结构特征的研究表明,割草地和放牧地的构成植物种及植物群落整体对幂乘方法则具有很好的吻合性。本试验旨在验证幂乘方法则能否应用于不同海拔梯度下草地群落结构特征的研究,依结果可知,3个不同海拔梯度草地的植物种及群落整体对幂乘方法则具有很好的吻合性,所以可在不同海拔梯度草地的空间异质性的定量研究中应用。
关山草原群落的总物种数随着海拔的升高而增加,草地群落的物种组成也发生了变化,其中,坡下草地退化的标志性植物蛇莓、毛茛成为主要的优势种,且它们降低了群落整体的异质性,禾草比例大大下降,杂类草比例上升,草地出现退化演替迹象。其原因是关山草原作为中国西北内陆地区的以高山草甸为主体的具有欧式风情的省级风景名胜区,其旅游业的发展非常迅速,游客的践踏和放牧的干扰(每年5月1日至10月10日),特别是马、牛、羊的混牧是造成草地严重退化的直接原因。坡下草地,距离公路较近,受上述因素干扰的影响较大,而坡中、坡上草地受到的干扰相对较小。同时不同海拔高度之间土壤养分、水分条件、光照、气温等自然因素存在一定的差异,这些因素通过长期、持久的相互作用影响草地植被,使得植物种类和植被随着海拔的上升呈明显的垂直分布[23]。但对于关山草原而言,其自然因素的影响远不及人类活动的影响大。所以建议关山草原在管理上实行划区轮牧,对草地践踏比较严重的马采取圈养,旅游区应合理规划,设置游客行走通道,减少对草地的过度践踏,从而促进关山草原旅游业及畜牧业的持续、稳定、健康发展。
山地植物群落物种多样性随海拔高度的变化规律一直是生态学家感兴趣的问题[24]。关山草原地区气候湿润,植物群落物种多样性指数表现为坡下<坡中<坡上,应属于贺金生和陈伟烈[25]分类模式的第4种,即随海拔高度的升高,植物群落物种多样性增加。在3个海拔梯度上,幂乘方法则的回归直线整体上位于y=x随机分布直线的上方,种群呈现团块分布的特点,这意味着群落整体比随机分布具有强的空间异质性,且群落整体的空间异质性指数也表现为坡下<坡中<坡上,因为坡下草地受放牧和游人践踏等人为过度干扰,使得植被组成较单一,异质性降低。
[1] 王绍强,周成虎,罗盛文.中国陆地自然植被碳量空间分布特征探讨[J].地理科学进展,1999,18(3):238-244.
[2] 郭健,郎侠.西部草地生态环境现状及其功能恢复措施[J].家畜生态学报,2005,26(5):1-3.
[3] 李晓宇,林震.退牧还草政策执行过程中的问题及建议[J].内蒙古农业科技,2011(1):1-2.
[4] 徐剑波,陈进发,胡月明,等.青海省玛多县草地退化现状及动态变化研究[J].草业科学,2011,28(3):359-364.
[5] 张文海,杨韫.草地退化的因素和退化草地的恢复及其改良[J].北方环境,2011,23(8):40-44.
[6] 施济普,朱华.三种干扰方式对西双版纳热带森林群落植物多样性的影响[J].广西植物,2002,22(2):129-135.
[7] 张金屯,阎耀川,金义兴.植物群落与异质性[J].武汉植物研究,1995,13(2):329-336.
[8] 陈宝瑞,杨桂霞,张宏斌,等.不同干扰类型下羊草种群的空间格局[J].生态学报,2010,30(21):5868-5874.
[9] 许清涛,黄宁,巴雷.不同放牧强度下草地植物格局特征的变化[J].中国草地学报,2007,29(2):7-12.
[10] 胡旭,王海涛,卢建国,等.干旱和半干旱区油蒿对土壤空间异质性的响应[J].中国沙漠,2007,27(4):588-592.
[11] Legendre P.Spatial auto correlation:Trouble or new paradigm[J].Ecology,1993,74:1659-1673.
[12] 李兆国.浅谈生态旅游[J].当代生态农业,2011(3-4):150-155.
[13] 塩見正衛,安田泰輔.草地の新しい植生調査法(1):ベータ二項分布を利用した植生調査[J].畜産の研究,2003,57(1):171-175.
[14] 陳俊,堀良通,塩見正衛,等.半自然草地および森林化した長期放任草地の群集構造[J].日本草地学会誌,2005,51(2):143-151.
[15] Shiyomi M,Takahashi S,Yoshimura T,etal.Spatial heterogeneity in a grassland community:Use of power law[J].Ecological Research,2001,16(3):487-495.
[16] 塩見正衛,安田泰輔.草地の新しい植生調査法(2)——べき乗則による解析[J].畜産の研究,2003,57(2):303-306.
[17] Chen J,Huang D M,Shiyomi M,etal.Spatial heterogeneity and diversity of vegetation at the landscape level in Inner Mongolia,China,with special reference to water resources[J].Landscape and Urban Planning,2007,82(4):222-232.
[18] Yasuda T,Shiyomi M,Takahashi S.Differences in spatial heterogeneity at the species and community levels in semi-natural grasslands under different grazing intensities [J].Grassland Science,2003,49(2):101-108.
[19] Song Z Y,Huang D M,Shiyomi M,etal.Spatial heterogeneity and variability of a large-scale vegetation community using a power-law model[J].Tsinghua Science and Technology,2005,10(4):469-477.
[20] Pielou E C.Mathematical Ecology[M].New York:John Wiley & Sons.,1977:224-266.
[21] 盐见正卫,安田泰铺,陈俊.关于放牧草地植被的调查方法[J].草地学报,2005,13(2):149-158.
[22] 吕杰,陈俊.不同利用方式下额尔古纳草原群落结构特征的研究——幂乘方法则的应用[J].草地学报,2011,19(3):388-394.
[23] 韩建国.草地学[M].第3版.北京:中国农业出版社,2007:47.
[24] 段敏杰,高清竹,郭亚奇,等.藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局[J].草业科学,2011,28(10):1845-1850.
[25] 贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征[J].生态学报,1997,17(1):91-99.
