转基因甘蓝型油菜品系W-4 高油酸性状遗传分析
2013-02-23申爱娟周晓婴戚存扣
申爱娟, 陈 松, 周晓婴, 戚存扣
(1.南京农业大学农学院,江苏 南京210095;2.国家油料作物改良中心南京油菜分中心,江苏 南京210014;3.农业部长江下游棉花与油菜重点实验室,江苏 南京210014)
油料作物种子中多含有油酸、亚油酸和亚麻酸等不饱和脂肪酸[1]。油酸(C18∶ 1)属单不饱和脂肪酸,它能够降低人体血液中低密度脂蛋白质含量,降低心血管疾病发病几率。同时油酸的化学稳定性优于多不饱和脂肪酸如亚油酸和亚麻酸,因此,高油酸植物油已被认为是氢化植物油的理想替代品[2-3]。目前,提高油料作物种子中的油酸含量,降低多不饱和脂肪酸含量已成为油料作物品质育种的重要目标之一[4-7]。
据报道植物高油酸种质主要通过诱变育种和基因工程育种两条途径实现。一是对油菜种子或小孢子进行化学诱变[8-9],或物理诱变[10],选择高油酸含量突变体,并应用系统选育与杂交育种技术获得高油酸含量油菜材料。二是利用共抑制(Co-suppression)[11]、反义RNA[5]或RNAi 基因沉默[4,12]等基因工程技术,特异性抑制油菜种子中fad2 基因表达,阻碍油酸进一步脱氢生成亚油酸,而获得高油酸含量的甘蓝型油菜种质。
已有报道,应用植物数量性状主基因+多基因的多世代联合分析方法,对甘蓝型油菜高油酸与低油酸组合后代群体油酸含量进行遗传分析,结果显示,油酸含量受2 对加性-显性-上位性主基因+加性-显性-上位性多基因控制遗传[13-15],并以主基因遗传为主;控制油酸含量的基因效应以加性和上位性效应为主,显性效应较小,没有细胞质遗传效应[13-15]。对诱变获得的甘蓝型油菜高油酸性状的遗传研究结果表明,高油酸性状的遗传受1 对主基因和多个微效多基因控制[16]。有关甘蓝型油菜高油酸性状遗传的结果不尽相同,而且试验材料多为诱变或系统选育获得的高油酸材料,而对转基因获得的高油酸材料的油酸含量的遗传规律鲜有报道。
W-4 是通过农杆菌介导获得的转基因高油酸甘蓝型油菜品系[12]。分子检测结果显示,转基因甘蓝型油菜品系W-4 只含有一个转基因拷贝[17]。有文献报道转基因的拷贝数低(1 或2 个)往往有较好的表达[18],有利于育种上应用。为了明确W-4 高油酸性状的遗传特性,以便更好地将其应用于甘蓝型油菜高油酸育种计划,本试验对转基因甘蓝型油菜高油酸品系W-4 与野生型亲本Westar 杂交后代群体油酸含量进行研究。
1 材料与方法
1.1 材料
甘蓝型油菜转基因品系W-4 和Westar 均由江苏省农业科学院经济作物研究所油菜研究室提供。2009 年秋季,在人工隔离网室内种植W-4(P1)和Westar(P2)2 个亲本。2010 年,于油菜开花期人工配置正、反交组合P1× P2(F1)、P2×P1(RF1),获得正交F1和反交RF1种子。2010 年秋季在人工隔离网室内种植P1、P2、F1、RF1。2011 年花期人工配置回交组合:B1(F1×P1)、B2(F1×P2);RB1(RF1×P1)、RB2(RF1×P2)。植株成熟时获得亲本P1、P2、F2、RF2及与双亲回交组合B1、B2、RB1和RB2种子。2011 年秋在人工隔离网室进行播种,用于苗期Kan 检测。
1.2 方法
1.2.1 种子脂肪酸组成测定 油菜种子的脂肪酸组成受胚基因型控制。因此种子脂肪酸组成分析直接随机选取当代成熟种子进行。其中亲本P1、P2各取10 粒、F2和RF2各取200 粒,其他世代群体分别取100 粒。种子脂肪酸含量在Agilent 6890-GC 气相色谱仪上进行定量检测。测定方法参照GB/T 17376-2008[19]。
油酸含量分级参照Schietholt 等[16]分级方法:以亲本Westar 种子中油酸含量为基础,当种子油酸含量≥Westar+2 倍标准差(SD)时的个体为高油酸,种子油酸含量≤Westar+2 倍标准差(SD)为低油酸。
1.2.2 油菜幼苗Kan 抗性检测 当幼苗长至3 ~4片真叶时,用1 g/L Kan 液喷洒植株,10 d 后调查抗性反应。叶片出现黄色斑点的植株为Kan 敏感型植株,叶片无黄斑的植株为Kan 抗性植株。
1.2.3 适合度测定 对正反交各世代群体高油酸与低油酸含量分离比进行适合性卡方(χ2)测验。
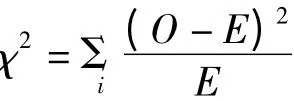
式中O 为实测值,E 为预期值。
2 结果
2.1 转基因油菜W-4 和野生型Westar 不饱和脂肪酸含量的比较
转基因油菜种子中不饱和脂肪酸含量测定结果(图1)显示,W-4 种子中油酸含量为(85.10 ±0.73)%,亚油酸和亚麻酸含量分别为(3.47 ±1.60)%和(3.38 ±0.54)%;Westar 种子中油酸含量为(62.16 ±3.68)%,亚油酸和亚麻酸含量分别为(19.17 ±2.20)%和(7.08 ±0.89)%。
W-4 种子中油酸含量比野生型亲本Westar 高22.94 个百分点,而亚油酸、亚麻酸含量分别比Westar 下降15.70 和3.70 个百分点。这与转基因甘蓝型油菜品系W-4 种子中表达了油酸脱饱和酶(fad2)基因的反向重复序列及诱导了fad2 基因的转录后基因沉默有关[20]。
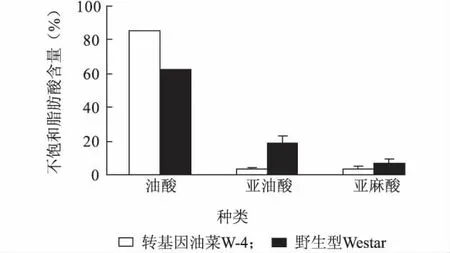
图1 转基因油菜W-4 和野生型Westar 种子中不饱和脂肪酸含量Fig.1 Unsaturated fatty acid contents in the seeds of transgenic oilseed rap W-4 and wild type Westar
2.2 正反交F1 种子中油酸含量
正反交(F1、RF1)种子脂肪酸含量测定结果显示,F1油酸含量为(77.55 ±1.30)%,RF1油酸含量为(77.34 ± 2.46 )%,均 高 于 双 亲 中 间 值(73.63%),与高油酸亲本W-4 值接近。表明,W-4高油酸性状为显性性状,无明显的细胞质遗传效应。
2.3 F2 代和回交后代种子中油酸含量
F2、RF2、B1、RB1、B2和RB2世代群体油酸含量结果(表1)显示,F2群体中144 个个体表现为高油酸含量(≥69.50%),56 个个体表现为低油酸含量(<69.50%);RF2群体148 个个体表现为高油酸含量,52 个表现为低油酸含量,经检验F2和RF2群体均符合3∶ 1 分离(χ2=0.96,χ2=0.11);B2、RB2高油酸与低油酸含量个体呈1∶ 1 分离(χ2=1.0,χ2=0.16)。B1、RB1群体所有个体均表现为高油酸含量。上述结果表明,转基因高油酸性状受1 对显性基因控制遗传。
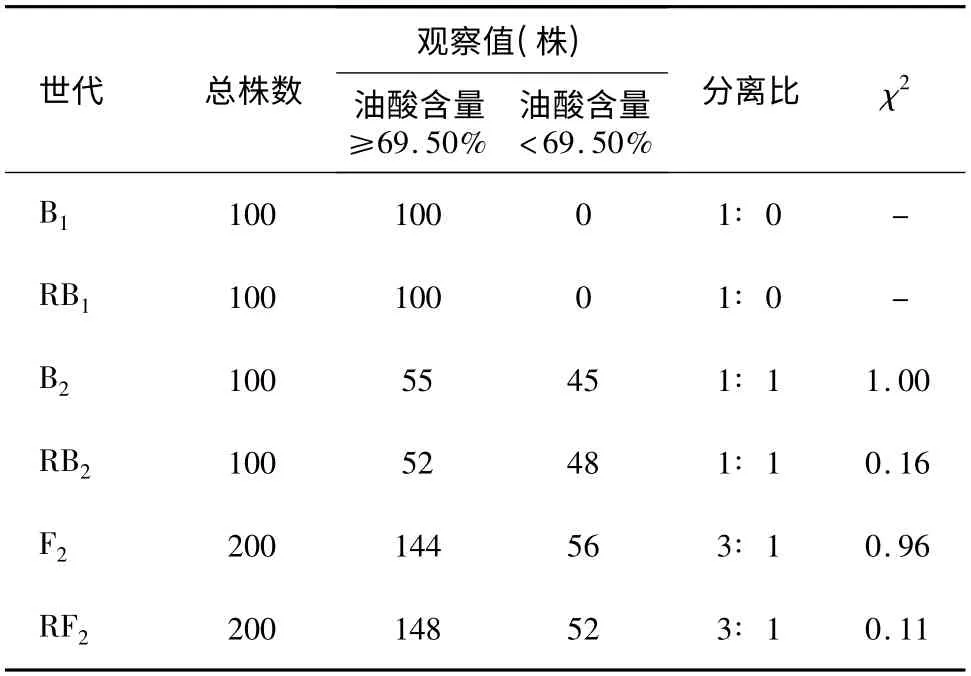
表1 F2、RF2、B1、RB1、B2和RB2世代群体的油酸含量Table 1 The oleic acid contents in the seeds of generations F2,RF2,B1,RB1,B2 and RB2
2.4 Kan 抗性检测
各个世代群体苗期Kan 抗性检测结果(表2)显示,P1(W-4)、F1、RF1、B1、RB1群体植株对Kan 反应均表现为抗性;而P2(Westar)群体植株对Kan 反应均表现为敏感;与Westar 回交世代B2、RB2群体植株对Kan 反应表现分离,抗、感比分别为47∶ 39 和48∶ 42,均呈1∶ 1 分离,F2、RF2群体植株间对Kan反应的抗、感比均呈3∶ 1 分离。结果显示,转基因油菜W-4 的Kan 抗性受1 对显性基因控制。上述结果表明,转基因油菜W-4 基因组中含有1 个拷贝的卡那霉素抗性基因,与转基因拷贝数分析结果一致[17]。
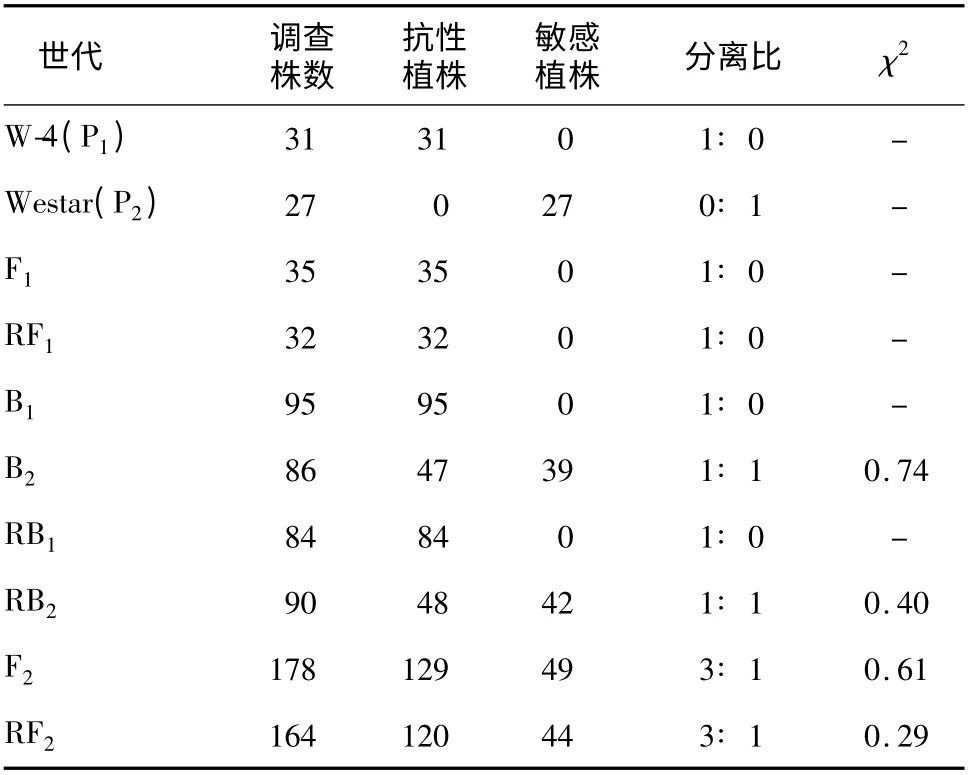
表2 各个世代群体苗期Kan 抗性反应Table 2 Resistance of seedling of each population to kanamycin
3 讨论
油菜种子中油酸含量的遗传一直是油菜品质性状研究的热点之一。Schietholt 等对甘蓝型油菜高油酸突变体油酸含量遗传分析发现,高油酸性状是由2 个位点的突变所致,其中一个位点HO1 主要在种子中表达,所有突变被认为是该位点的等位基因;另一个突变位点HO2,既增加种子中油酸含量,也提高叶片和根中油酸含量,这2 个位点表现出加性效应,并观察到不显著的显性效应,无上位效应和母性效应[16]。索文龙等、张洁夫等应用植物数量性状主基因+多基因多世代联合分析法,对油酸含量分别为65.33%和10.91%的甘蓝型油菜杂交组合的油酸含量进行遗传分析,结果表明油酸含量受2 对加性-显性-上位性主基因+加性显性上位性多基因控制,基因效应以加性效应为主,显性效应较小[13-14]。费维新等对低油酸与高油酸突变体的杂交后代F1、RF1、B1、RB1、B2、RB2、F2和RF2的高油酸性状进行了数量性状遗传分析,认为高油酸性状由2 对主效基因控制,其狭义遗传力为63.16%,正反交试验表明,高油酸的遗传没有细胞质遗传效应[15]。然而,官梅等以高油酸突变体与常规低芥酸油菜为材料,研究油菜油酸含量的遗传,结果显示油菜油酸性状属数量性状,杂种1 代表现为中间类型,并受母本影响,支配油酸性状的基因数为1 对主基因,油酸含量遗传力为69.00% ~71.00%,并表现出对环境敏感[21]。有关甘蓝型油菜油酸含量的遗传结果不尽相同,这可能与试验材料的不同有关。
然而,对转基因获得的高油酸油菜油酸含量的遗传特点尚未见报道。本试验材料高油酸油菜品系W-4 是利用RNAi 基因沉默技术获得的高油酸油菜新种质。W-4 的fad2 基因的反向重复序列是在油菜napin 启动子控制下表达的。有研究显示该启动子在开花后21 d 的种胚中开始表达[20,22],理论上功能正常的napin 启动子控制的性状应表现为显性性状。本研究以高油酸油菜品系W-4 与其野生型非转基因对照Westar 杂交获得的衍生世代为材料,分析各世代单株种子中油酸含量,结果表明转基因油菜W-4 高油酸性状受1 对显性基因控制,无明显的细胞质遗传效应;此外通过对各世代植株对卡那霉素抗性分析,结果显示转基因油菜W-4 对卡那霉素的抗性也受1 对显性基因控制。这些结果与陈松等[7]对转基因油菜W-4 的T-DNA 拷贝数的分析结果一致;核酸杂交试验结果显示W-4 整合了一个拷贝的T-DNA[18]。当外源基因以单拷贝方式插入染色体的某一位点时,整合的外源基因拷贝可作为一个独立的遗传单位共同分离,即表现孟德尔遗传。综合多方面的数据可认为W-4 的高油酸性状受1对显性单基因控制遗传,这正是W-4 高油酸性状能够稳定遗传的基础。结合应用Kan 检测,W-4 的高油酸性状更容易整合到育种材料中,使得高油酸品种(系)筛选更加简单易行。可见W-4 作为一个高油酸新种质,在油菜高油酸育种上的应用前景十分广阔。
[1] 李成磊,付三雄,戚存扣. 甘蓝型油菜种子发育过程中主要脂肪酸的积累及相关分析[J]. 江苏农业学报,2011,27(2):258-263.
[2] ORTHOEFER F T. Performance of trans-free vegetable oils in shortenings and deep-fat frying[J]. Lipid Technology,2005,17(5):101-106.
[3] NAPIER J A. The production of unusual fatty acids in transgenic plants[J]. Annual Review of Plant Biology,2007,58:295-319.
[4] YIN D M,DENG S Z,ZHAN K H,et al. High-oleic peanut oils produced by hpRNA-mediated gene silencing of oleate desaturase[J]. Plant Molecular Biology Reporter,2007,25(3-4):154-163.
[5] SIVARAMAN I,ARAMUGAM N,SODHI Y S,et al. Development of high oleic and low linoleic acid transgenic in a zero erucic acid Brassica juncea L. (Indian mustard)line by antisense suppression of the fad2 gene[J]. Molecular Breeding,2004,13(4):365-375.
[6] LACOMBE S,SOUYRIS I,BERVILLÉ A J. An insertion of oleate desaturase homologous sequence silencing via siRNA:the functional gene leading to high oleic acid content in sunflower seed oil[J]. Mol Genet Genomics,2009,281(1):43-54.
[7] LIU Q,SINGH S P,GREEN A G. High-stearic and high-oleic cotton-seed oils produced by hairpin RNA- mediated post-transcriptional gene silencing[J]. Plant Physiology,2002,129(4):1732-1743.
[8] 张育军,肖 钢,谭太龙,等. EMS 处理甘蓝型油菜(Brassica napus L.)获得高油酸材料[J]. 中国农业科学,2008,41(2):4016-4022.
[9] 和江明,王敬乔,陈 嶶,等.用EMS 诱变和小孢子培养快速获得甘蓝型油菜高油酸种质材料的研究[J]. 西南农业学报,2003,16(2):34-36.
[10] 官春云,刘春林,陈社员,等. 辐射育种获得油菜高油酸材料[J].作物学报,2006,32(11):1625-1629.
[11] STOUTJESDIJK P A,HURLSTONE C,SINGH S P,et al. Higholeic acid Australia Brassica napus and B . juncea varieties produced by co-suppression of endogenous delta-12 desaturases[J].Biochemical Soc Trans,2000,28:938-940.
[12] 陈 松,浦惠明,张洁夫,等. 转基因高油酸甘蓝型油菜新种质的获得[J]. 江苏农业学报,2009,25(6):1234-1237.
[13] 索文龙,戚存扣.甘蓝型油菜油酸含量的主基因+多基因遗传分析[J].江苏农业学报,2007,23(5):396-400.
[14] 张洁夫,戚存扣,浦惠明,等. 甘蓝型油菜主要脂肪酸的主基因+多基因遗传分析[J]. 中国油料作物学报,2007(4):359-364.
[15] 费维新,吴新杰,李强生,等.甘蓝型油菜高油酸材料的遗传分析[J].中国农学通报,2012,28(1):176-180.
[16] SCHIETHOLT A,BECKER H C. Environmental variability and heritability of high oleic acid content in winter oilseed rape[J].Plant Breeding,2001,120(1):63-66.
[17] 陈 松,张洁夫,浦惠明,等. 转基因高油酸油菜T-DNA 插入拷贝数及整合位点分析[J]. 分子植物育种,2011,9(1):17-24.
[18] STOUTJESDIJK P A,SINGH S P,LIU Q,et al. Hp-RNA-mediated targeting of the Arabidosis FAD2 gene gives highly efficient and stable silencing [J]. Plant Physiology,2002,129(4):1723-1731.
[19] GB/T 17376-2008 动植物油脂脂肪酸甲酯制备[S].
[20] 周晓婴,申爱娟,张洁夫,等.RNAi 沉默转基因油菜fad2 基因表达的种子特异性分析[J]. 分子植物育种,2012,10(3):305-310.
[21] 官 梅,李 栒.油菜(Brassica napus)油酸性状的遗传规律研究[J].生命科学研究,2009,13(2):152-157.
[22] HÖGLUND A S,RÖDIN J,LARSSON E,et al. Distribution of napin and cruciferin in developing rapeseed embryos[J]. Plant Physiol,1992,98:509-515.
