基于TRIPLEX模型的湖南省杉木林生产力模拟及预测
2012-12-27项文化赵梅芳邓湘雯彭长辉
王 灿 ,项文化 ,赵梅芳 ,邓湘雯 ,彭长辉
(1. 湖南会同杉木林生态系统国家野外科学观测研究站,湖南 会同 438107;2. 中南林业科技大学 生命科学与技术学院,湖南 长沙 410004;3. 加拿大魁北克大学 环境科学研究所,加拿大 蒙特利尔 H3C 3P8)
基于TRIPLEX模型的湖南省杉木林生产力模拟及预测
王 灿1,2,项文化1,2,赵梅芳1,2,邓湘雯1,2,彭长辉1,3
(1. 湖南会同杉木林生态系统国家野外科学观测研究站,湖南 会同 438107;2. 中南林业科技大学 生命科学与技术学院,湖南 长沙 410004;3. 加拿大魁北克大学 环境科学研究所,加拿大 蒙特利尔 H3C 3P8)
杉木Cunnighamia lanceolata是我国优良速生用材树种,其面积达921万hm2,占全国人工用材林面积的30.4%。用机理性模型在区域尺度上研究杉木林生产量可为估算杉木林固碳潜力、预测其对气候变化的响应和提高经营管理水平提供科学依据。利用气象、水文、土壤及植物等方面数据,对TRIPLEX 1.6 模型进行参数化,用森林资源调查样地数据对模型进行验证,模拟湖南省杉木林1991~2040年生物量与生产力动态变化、空间分布格局。结果表明:模拟值与实测值之间拟合度高(p=0.995),TRIPLEX1.6 模型可应用于模拟亚热带杉木林生长和生产量的动态变化。2010~2018年湖南省杉木林生产力(NPP)为迅速增长期,此后NPP缓慢下降,2008年所有样地杉木林平均年龄为23 a,单位面积平均NPP达到最大值(8.09 t·hm-2a-1),2010~2060年间平均值为6.96 t·hm-2a-1。
杉木林;生物量动态;TRIPLEX 模型;空间分异;气候变化
生物量和生产力反映森林生态系统中有机物质积累总量和速率,是整个生态系统运行的能量基础和营养物质来源[1]。由于森林生物量与森林碳贮量紧密相关,随着近年来人们对森林固碳研究的重视,有关区域或国家尺度森林生物量估算以及动态变化等成为研究的热点[2-6]。20世纪70年代末期,国内在林分尺度上开展了杉木、马尾松等森林生物量和生产力的研究[7-9]。杉木是我国南方集体林区栽培历史悠久、栽培最大的用材树种,有关杉木林生物量和生产力研究数据较丰富,多来源于收获法,还有基于森林资源清查数据以及遥感资料的推算以及林分模型的应用[10-12]。我国杉木林生物量宏观尺度的空间分异格局研究方面,有一些学者对其进行了分析[11-13],但由于数据的限制、研究侧重点不同以及区域性差异等,难以预测杉木林的未来变化及其对气候变化响应。
机理性模型为模拟森林生产力及对气候变化响应提供了有效途径。20世纪80年代末森林生态系统模型相继开发,主要分经验模型和过程模型[14-15],但是混合模型弥补两类模型的不足。从20世纪90年代开始,混合模型被广泛应用于生态系统结构和功能研究[16-23]。TRIPLEX模型是一种混合模型,包括3-PG模型的森林产量子模型[24]、TREEDYN3.0模型的光合辐射(PAR)子模型以及树木增长和产量的子模型[25]以及CENTURY4.0模型中的土壤碳和分解子模型[26],在世界范围内获得较好的检验[19-22]。
湖南省地处亚热带,水热条件优越,是杉木产区中心产区之一,目前对区域尺度上杉木林产量和固碳潜力及全球气候变化响应缺乏深入研究。本研究利用2009年森林资源清查样地数据,验证TRIPLEX模型对湖南省杉木林的模拟,分析湖南省杉木林生产量从1990~2040年间动态变化规律,为湖南省森林生产力、碳贮量和大面积森林固碳潜力估算提供科学依据。
1 研究区概况
湖南省位于我国长江中游,地处东经108°47′~ 114°15′,北纬 24°38′~ 30°08′之间。地貌东南西三面环山,中部、北部低平,形成向北开口的马蹄形盆地,境内山地约占总面积一半,平原、盆地、丘陵、水面约占一半;土壤为红壤、黄壤地带(图1)。气候为亚热带大陆性季风湿润气候,全年总辐射量为360~456 kJ·cm-2,年平均气温16℃~18.5℃,年均降水量1 300~1 700 mm。由于光、热、水资源丰富,气候在年际和季节上的多变性以及气候类型的多样性等特征,对森林分布及生长发育产生重要影响。

图1 湖南省的地貌、海拔和土壤特征的空间格局Fig. 1 Spatial distribution pattern of topography, altitude (above mean sea level), and soil in Hunan
2 模型应用方法和验证
2.1 数据来源和模型参数化
本研究用TRIPLEX1.6模型对湖南省杉木林进行模拟,模型结构、原理及应用情况可参考文献[19,22]。TRIPLEX 模型需要输入的数据包括气候、土壤、林分类型、树龄和树种等,输出结果有林分的胸径、树高和生物量等。其中,输入的林分数据来自于湖南省2009年森林资源调查数据,包括702个杉木林样地(图2),2009年调查的杉木林样地林龄在1~47 a之间,平均林龄为15 a。
TRIPLEX模型模拟需要输入的气候数据包括每月降雨量(mm)、每月平均气温(℃)、每月平均水汽蒸汽压差(DVPD)和月平均有霜天数,本研究利用2000~2010年所有样地点的气象资料,计算出10年间各月降雨量(mm)、每月平均气温(℃)、每月月最高和最低气温 (Tmax,Tmin)。通过Tmax和Tmin来计算DVPD(mbar)(公式1)。用于计算GPP的有霜比率 (IFrostdays/month) 取决于月最低气温 (Tmin)(公式2),数据结果见图3和图4。


图2 湖南省杉木林林分初始化参数(2009年森林资源清查资料)Fig. 2 Parameters related to initial status of C. lanceolata forest recorded in 2009 in Hunan

图3 湖南省杉木林2000~2010年间的月平均降雨量、气温、霜冻天数以及水汽压差Fig. 3 The monthly average records of precipitation,temperature, frost days, and DVPD from 2000 to 2010 for C. lanceolata forest sites in Hunan
模型的参数化主要是确定气候因子、研究地点基本情况、外界营养输入、土壤有机质初始含量、森林植物有机质初始含量、土壤矿物质初始含量、土壤水分状况等多方面的参数[19],这些参数主要根据文献中数据,各参数的含义和确定方法可参阅TRIPLEX1.6模型说明。
2.2 模型验证和参数调整
模型验证主要是检验模拟值与实测值之间的吻合程度,我们用702个样地中的351个样地数据作为输入数据进行模拟,剩下的351个样地数据进行验证,采用独立样本t检验方法,在统计软件SPSS13.0进行统计分析,检验模拟结果,调整模型标准参数,直至模拟值与观测值之间不存在显著性差异。

图4 湖南省杉木林2000~2010年间的年平均降雨量、气温、霜冻天数以及水汽压差的空间分异格局Fig. 4 Annual average records of precipitation, temperature, frost days, and DVPD from 2000 to 2010 for all C. lanceolata forest sites in Hunan
3 结果与分析
3.1 模型验证和可靠性分析
利用2009年森林资源调查的样地数据,对杉木林胸径和树高进行模拟和比较,模拟值与实测值均非常接近(R2>0.8),实测值与模拟值的双尾t检验概率P>0.05(PH=0.128 13,PDBH=0.586 06),说明实测值与模拟值之间没有显著差异 (图6),表明TRIPLEX1.6模型可用来模拟杉木林生长过程及生产量。
3.2 杉木林林分生物量空间格局及其动态

图5 湖南省杉木林林分平均胸径(a)及树高(b)的模拟值和观察值的比较Fig. 5 Comparison between simulated and observed stand average tree (a) diameter at breast height (DBH, cm) and (b)height (H, m) of C. lanceolata forests in Hunan
湖南省702块杉木林样地生物量在2010~2060年间空间格局(图6)及变化情况(表1)表明,2010年生物量范围为0.16~137.74 t·hm-2,均值30.19 t·hm-2,其中80%的样地生物量低于50 t·hm-2。2020年生物量范围为16~146 t·hm-2之间,均值为73.27 t·hm-2,其中30%的样地生物量低于50 t·hm-2,约40%样地生物量为100~150 t·hm-2。2030年生物量范围为53~183 t·hm-2之间,均值为114.66 t·hm-2,其中20%的样地生物量低于100 t·hm-2,约50%的样地生物量为100~150 t·hm-2。2040年生物量范围为79~200 t·hm-2,均值为150.09 t·hm-2,其中60%的样地生物量高于150 t·hm-2,约20%的样地生物量为100~150 t·hm-2。2050年杉木林生物量为96~249 t·hm-2,均值 181.02 t·hm-2,其中 20% 的样地生物量高于200 t·hm-2,约60%的样地生物量为150~200 t·hm-2。2060年杉木林生物量为96~ 249 t·hm-2,均值为 209.19 t·hm-2,其中 5%的样地生物量高于250 t·hm-2,约40%的样地生物量为200~250 t·hm-2,35%小于200 t·hm-2。
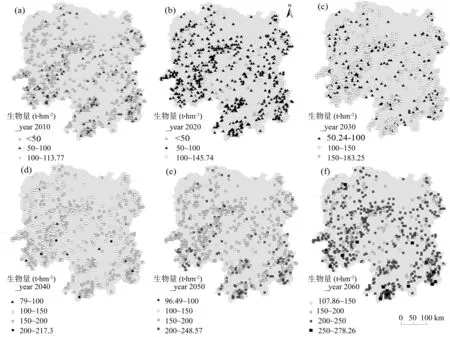
图6 湖南省杉木林分布点生物量现存量(t·hm-2)的空间动态的预测Fig. 6 Simulated spatial pattern potential of total biomass (t·hm-2) of C. lanceolata forest s in Hunan
从全省范围来看,生物量分布整体呈现出自北部中山高海拔地带向到中东南部丘陵沿海低海拔地区逐渐增大的趋势,西南和东南地带生物量密度最高。随着时间的推移,2020年杉木林生物量比2010年增加了142.68%, 2030年比2020年增加了56.49%,2040年比2030年增加了30.90%,2050年比2040年增加了20.61%,2060年比2050年增加了15.57%,近60年间的增加幅度是逐渐变小的。

表1 湖南省杉木林生物量(t·hm-2)60年动态变化Table 1 Changes in total biomass of C. lanceolata forests in recent 60 years in Hunan t·hm-2
3.3 净初级生产力(NPP)与总生物量动态变化预测
根据湖南省702个杉木林样地林分平均净初级生产力(t·hm-2a-1)与生物量( t·hm-2)年际变化模拟结果,乘以湖南省杉木林总面积2.519 035×106hm2[11],即计算出湖南省杉木林总NPP(106t·a-1)和生物量(106t)的年际变化(图7)。

图7 湖南省杉木林初级生产力及生物量模拟动态及预测Fig. 7 The simulated NPP and total biomass of C. lanceolata forest s in Hunan
湖南省杉木林平均NPP和总NPP在2010~2060年间的均值分别为6.96 t·hm-2a-1和17.53×106t·a-1。2010年,湖南省杉木林样地平均林龄为16 a,平均NPP为6.28 t·hm-2a-1,总NPP为15.81×106t·a-1。随着林龄增加NPP迅速增大,2010年杉木林样地平均林龄为23年,平均NPP和总NPP分别为8.09 t·hm-2a-1和20.38×106t·a-1,达到最大值,此后平均NPP和总NPP随着林龄增加而缓慢减少;到2060年,杉木林样地平均林龄为74 a,平均NPP和总NPP达到最小值,分别为 5.87 t·hm-2a-1和14.793×106t·a-1。湖南省杉木林平均生物量和总生物量在2010~2060年间的平均值分别为145.24 t·hm-2和365.87×106t,最小值分别为2010年的28.97 t·hm-2和72.99×106t,最大值分别为2060年的235.66 t·hm-2和593 63×106t。
4 结 论
模型是基于特定目的对真实系统组成、结构及功能关系简化或抽象的表述,本研究用TRIPLEX模型较准确地模拟和预测湖南省杉木林生物量与生产力变化。2010~2060年湖南省杉木林平均NPP为 6.96 t·hm-2a-1,高于我国森林平均值(约4.8~6.22 t·hm-2a-1)[27],表明杉木林作为速生树种,其生产力及固碳潜力大。模拟的2010~2015年间湖南省杉木林生物量为72.99~127.48×106t,与焦秀梅等利用湖南省森林资源数据(1990~1995年)估算的2005年的结果(103.175×106t)相似[11]。本研究应用TRIPLEX模型模拟和预测湖南省杉木林生物量与生产力是基于系列前提条件或预定方案进行的,如大气CO2浓度及N沉降维持在初始化水平,同时由于杉木林结构较简单,没有考虑模拟过程中可能发生的森林结构变化,也没有涉及真实系统中可能发生的采伐、病虫害、风等干扰。但是,与上述假设条件的相关内容都是目前生态学的热点研究内容,本研究为进一步模拟森林生态系统对气候变化响应、干扰及生态学效应、生态系统结构与功能相互作用提供验证和基础参数。
[1] 冯宗炜, 王效科, 吴 刚. 中国森林生态系统的生物量和生产力[M]. 北京:科学出版社, 1999.
[2] Dixon R K, Brown S, Houghton R A, et al. Carbon pools and flux of global forest ecosystems[J]. Science, 1994,263:185-190.
[3] 方精云, 刘国华, 徐嵩龄. 我国森林植被的生物量和净生产量[J]. 生态学报, 1996, 16(5): 497-508.
[4] 周玉荣, 于振良, 赵士洞. 我国主要森林生态系统碳贮量和碳平衡[J]. 植物生态学报, 2000, 24 (5):518-522.
[5] 刘国华, 傅伯杰, 方精云. 中国森林碳动态及其对全球碳平衡的贡献[J]. 生态学报, 2000,20(5):733- 740.
[6] 于贵瑞. 全球变化与陆地生态系统碳循环和碳蓄积[M]. 北京:气象出版社, 2003.
[7] 冯宗炜, 陈楚莹, 张家武, 等. 不同自然地带杉木林的生物生产力[J]. 植物生态学与地植物学丛刊, 1983, 8 (2) :93 -100.
[8] 叶镜中, 姜志林, 周本琳. 福建省洋口林场杉木林生物量的年变化动态[J]. 南京林学院学报, 1984, (4) : 1 - 9.
[9] 潘维俦, 田大伦, 高正衡. 杉木人工林生态系统中的生物产量及其生产力的研究[J]. 中南林业科技, 1978, (2) : 2 - 14.
[10] 冯宗炜, 陈楚莹, 张家武. 湖南会同地区马尾松林生物量的测定[J]. 林业科学, 1982, 18 (2) : 127 - 134.
[11] 焦秀梅, 项文化, 田大伦. 湖南省森林植被的碳贮量及其地理分布规律[J]. 中南林学院学报, 2005, 25(1): 4-9.
[12] Zhao M F, Xiang W H, Peng CH, et al. Simulating age-related changes in carbon storage and allocation in a Chinese fir plantation growing in southern China using the 3-PG model [J].Forest Ecology and Management, 2009, 257: 1520-1531.
[13] Fang J Y, Piao S L, Field C B, et al. Increasing net primary production in China from 1982 to 1999 [J]. Frontiers in Ecology and the Environment, 2003, 1: 293–297.
[14] Kimmins J P. Community organization: methods of study and prediction of the productivity and yield of forest ecosystems[J].Canadian Journal of Botany, 1988, 66: 2654-2672.
[15] Kimmins J P. Modeling the sustainability of forest production and yield for a changing and uncertain[J]. Forestry Chronicle,1990, 66: 271-280.
[16] Kimmins J P, Mailly D, Seely B. Modeling forest ecosystem net primary production: the hybrid simulation approach used in Forcast[J]. Ecological Modelling, 1999, 122: 195-224.
[17] Peng C H. Understanding the role of forest simulation models in sustainable forest management[J]. Environmental Impact Assessment and Review, 2000a, 20: 481-501.
[18] Peng C H. Growth and yield models for uneven-aged stands: past present and future[J]. Forest Ecology and Management, 2000b,132: 259-279.
[19] Peng C H, Liu J, Dang Q, et al. TRIPLEX: a generic hybrid model for predicting forest growth and carbon and nitrogen dynamics[J]. Ecological Modelling, 2002, 153: 109-130.
[20] Zhang J, Ge Y, Chang J, et al. Carbon storage by ecological service forests in Zhejiang Province, subtropical China[J]. Forest Ecology and Management, 2007, 245: 64–75.
[21] Zhao M, Zhou G S. Estimation of biomass and net primary productivity of major planted forests in China based on forest inventory data[J]. Forest Ecology and Management, 2005, 207:295–313.
[22] Zhou X L, Peng C H, Dang Q L. Assessing the generality and accuracy of the TRIPLEX model using in situ data of boreal forests in central Canada[J]. Environmental Modelling &Software, 2004, 19: 35–46.
[23] Zhou X L, Peng C H, Dang Q L, et al. Predicting forest growth and yield in northeastern Ontario using the process-based carbon dynamic model of TRIPLEX1.0[J]. Canadian Journal of Forest Research, 2005, 35: 2268–2280.
[24] Landsberg J J, Running R H. A generalized model of forest productivity using simplified concepts of radiation-use efficiency,carbon balance and partitioning[J]. Ecological Modelling, 1997,95: 209-228.
[25] Bossel H. TREEDYN3 Forest Simulation Model[J]. Ecological Modelling, 1996, 90: 187-227.
[26] Parton W J, Scurlock J M, Ojima D S, et al. Observations and modelling of biomass and soil organic matter dynamics for the grassland biome worldwide. Global Biogeochemical Cycles,1993, 7: 785- 809.
[27] 方精云. 我国森林植被的生物量和净生产量. 生态学报,1996, 16(5): 497-508.
Simulation and prediction of biomass spatial distribution of Cunninghamia lanceolata forests in Hunan, China based on TRIPLEX model
WANG Can1,2, XIANG Wen-hua1,2, ZHAO Mei-fang1,2, DENG Xiang-wen1,2, PENG Chang-hui1,3
(1.Huitong National Field station for Scientific Observation and Research of Chinese Fir Plantations Ecosystem in Hunan, Huitong 438107, Hunan, China; 2.School of Life Science and Technology, Central South University of Forestry and Technology, Changsha 410004, Hunan, China; 3. Institute of Environmental Sciences, University of Quebec at Montreal, Montreal H3C 3P8, Canada)
∶ Chinese fir is an excellent fast-growing timber tree species, having a planting area of 921× 104 hm2and accounting for 30.4% of plantation timber-forest area in China. Study on Chinese fir biomass with the mechanism model by regional scale can provide a scientific basis for estimating carbon sequestration potential, forecasting the effects of Chinese fir biomass on climate change, and raising the managerial skills. Therefore, the TRIPLEX model was parameterized by using the dataset of climate, hydrology, soil and vegetation, the model was validated with the simulation values of DBH and tree height collected from 702 permanent plots of Chinese fir stands, and the dynamic changes of biomass and productivity, spatial distribution pattern for Hunan in 1991~2040 were simulated.The results show that the fitting degree of simulation value and the measured value were very high(0.995), thus TRIPLEX 1.6 model can be applied to simulate the dynamic changes of growth and productivity of Chinese fir productivity is a rapid-increase stage, after that the productivity will decrease; the mean-age of all Chinese fir stands are 23-year-old, the average productivity per unit area reached the maximum value in 2008(8.09 t·hm-2a-1), the average value in 2010 ~ 2060 will reach 6.96 t·hm-2a-1.
∶ Cunninghamia lanceolata(Chinese fir); biomass dynamics; TRIPLEX model; spatial distribution pattern; climate changes
S791.27;S718.55+6
A
1673-923X(2012)06-0104-06
2012-03-11
国家林业局948项目(2010-4-03)和湖南省“芙蓉学者计划”项目资助
王 灿(1983—),湖南沅江人,硕士研究生,研究方向:森林生态学; E-mail: chancezx@163.com
项文化(1967—),湖北麻城人,教授,博导,主要从事森林生态学研究; E-mail: xiangwh2005@163.com
[本文编校:吴 彬]
