青藏高原高寒草甸生态系统CO2通量研究进展
2012-12-08乔春连李婧梅王基恒葛世栋徐世晓
乔春连,李婧梅,王基恒,葛世栋,赵 亮,徐世晓
(1.中科院西北高原生物研究所,青海 西宁 810008; 2.中国科学院研究生院,北京 100049; 3.中国科学院高原生物适应与进化重点实验室,青海 西宁 810008)
青藏高原高寒草甸生态系统CO2通量研究进展
乔春连1,2,3,李婧梅1,2,王基恒1,2,葛世栋1,2,赵 亮1,徐世晓1
(1.中科院西北高原生物研究所,青海 西宁 810008; 2.中国科学院研究生院,北京 100049; 3.中国科学院高原生物适应与进化重点实验室,青海 西宁 810008)
高寒草甸是广布于青藏高原的主要植被类型,是青藏高原大气与地面之间生物地球化学循环的重要构成部分,在区域碳平衡中起着极为重要的作用。本研究首先系统回顾了青藏高原高寒草甸生态系统CO2通量日、季、年等不同时间尺度的变化特征,以及温度、光合有效辐射、降水等主要环境因子对高寒草甸生态系统CO2通量的影响;其次比较了青藏高原3种典型高寒草甸生态系统类型源汇效应和Q10值;最后针对青藏高原高寒草甸生态系统CO2通量研究现状,分析了当前存在的一些不确定性,展望了未来工作的重点。
高寒草甸;CO2通量;净生态系统CO2交换量;温度
草地是陆地生态系统的重要组成部分,草地生态系统中碳素贮量的全球估计成为草地碳循环研究的关键之一[1-3]。以往对草地生态系统碳循环的研究大多集中于低海拔地区,近年来,高寒草甸因面积广大和分布的特殊性,成为研究生态系统碳循环、全球变化领域关注的热点地区之一。青藏高原隆升过程所形成的特殊自然环境,形成了草原化嵩草草甸(Steppe Kobresia meadow)、典型嵩草草甸(Type Kobresia meadow或True Kobresia meadow)和沼泽化嵩草草甸(Swamp Kobresia meadow)3种主要的嵩草草甸类型[4]。目前,关于天然高寒草甸生态系统碳循环的研究主要基于此3种类型进行。
高海拔生态系统较低海拔地区对全球变化的反应更敏感,且青藏高原地区自20世纪50年代以来变暖趋势已经超过北半球及同纬度地区[5],青藏高原昼夜温差大,在牧草生长季节夜间温度(10 ℃以下)明显低于白昼(20 ℃ 以上),夜间低温降低了呼吸消耗,利于牧草白天光合作用所固定的碳的积累;因此,青藏高原草地生态系统在区域生态系统碳平衡中起着极为重要的作用,可能对区域甚至全球水平的植被和大气界面间的CO2交换有显著贡献。本研究主要对近年来青藏高原高寒草地生态系统CO2通量的研究情况进行评述,将该区域碳源汇潜能和土壤呼吸的温度敏感性与世界不同草地类型相比较,提出目前研究可能存在和值得继续深入研究的问题,试为以后的研究提供借鉴。
1 高寒草地生态系统CO2净交换量的变化特征
高寒草地生态系统CO2交换过程包括植被通过光合、呼吸作用从大气吸收、释放CO2(大气-植被),植物生物量进入土壤分解、碳素再分配(植被-土壤)以及土壤中的有机物通过微生物分解作用释放CO2到大气中(土壤-大气)的过程[6]。在不考虑人为因素与动物活动影响的情况下,陆地与大气之间的净生态系统碳交换量(Net Ecosystem Exchange,NEE)相当于植物光合作用固定的碳通量与生态系统呼吸释放碳量(包括植物的自养呼吸和土壤微生物分解有机物和凋落物的呼吸)的差值[7],即NEE=GPP-Reco(GPP表示生态系统固定的总碳量,Reco表示生态系统呼吸释放的碳量)。由于青藏高原特殊的自然环境和气候特征,该区CO2循环呈现出自身特殊的变化特征。
1.1NEE日变化规律 净生态系统CO2交换量是指生态系统与大气圈之间的净碳交换量,是生态系统与大气间交换量的直接测度。高寒草甸和灌丛生态系统6-9月生长季中CO2通量呈现明显的日变化规律,表现为单峰型曲线,白天吸收大于晚间 排放,总体均表现为碳汇[8-11]。CO2通量从08:00左右通过零值后开始上升,表现为净吸收,最大吸收值一般出现在11:00-12:00,光合效率达到最高;20:00-07:00为CO2的净排放阶段。非生长季中,NEE日变化振幅较小,仅在白天11:00-18:00出现少量的净排放,其他时段CO2通量接近于零[8-9,11-12]。与温带草原相比,在低温抑制下的高寒灌丛生态系统非生长季的日平均CO2释放率较低[10]。高寒湿地生长季白天NEE的变化与高寒草甸和灌丛相似,表现为单峰型日变化,但总体上表现为明显的CO2排放源[13]。
1.2NEE季节变化规律 青藏高原植被生产力与雨热分布的密切关系,使NEE变化具有明显的随生长季和非生长季变化的趋势,生长季的8月一般是全年CO2吸收最高的月份,4月是全年CO2净排放最高的月份[8,12]。高寒灌丛生态系统在整个生长季CO2净吸收的总量达583 g·m-2。非生长季节(10-4月)CO2通量变化振幅极小,CO2净排放量为356 g·m-2[8,12]。生长季中,6-9月为CO2净吸收阶段,至10月初生长季结束,植物枯黄后转变为碳排放[8-9,11,14]。在非生长季各月NEE均较低,但差异明显,4和10月CO2净释放量较大,而1和12月CO2净释放量较小[10,12]。由于青藏高原东北部平均气温高于青藏高原腹地,该区域高寒灌丛具有相对较高的叶面积指数,致使其在短暂的生长季节具有较高CO2净吸收量。非生长季较低的土壤温度下,土壤微生物活动较弱,植被凋落物分解速率缓慢,在土壤表层堆积形成较厚的腐殖质层,导致大量碳素储存在土壤中。
1.3NEE年变化规律 矮嵩草草甸和灌丛草甸年吸收CO2量分别为282和227 g·m-2[8,15], 而沼泽化草甸年排放CO2约为478 g·m-2。表明青藏高原嵩草草甸和灌丛草甸可能具有一个相对较低的CO2碳汇和碳源潜能,而沼泽化草甸具有一个较高的排放源。海北高寒湿地生态系统生长季从大气吸收CO2230.16 g·m-2,非生长季向大气中排放CO2546.18 g·m-2,全年CO2释放量为316.02 g·m-2,较Zhao等[15]对沼泽化草地的测定值478 g·m-2高,高寒湿地生态系统均表现为碳源,有较高的排放潜能[13,15]。青藏高原不同植被类型的碳源汇功能存在明显的差异,这主要是由植物光合能力不同和土壤呼吸差异所引起的[15]。
2 影响高寒草地生态系统CO2通量变化的因素
任何影响植物生长发育的自然因素(如光强、温度、降水等)和人为因素(放牧、干扰、火烧等)均可能影响到高寒草地生态系统CO2通量,其中有些因素贡献较大,有些较小。
2.1光合有效辐射、叶面积指数对NEE的影响 光合有效辐射是影响光合作用的主要因子之一,青藏高原地区太阳辐射强烈,是我国日照时间最长的地区之一,强烈的太阳辐射有利于高寒草地植被光合作用。在整个生态系统呼吸量一定的条件下,生态系统的总初级生产力(Gross Primary Productivity,GPP)越大,固定的碳越多,生态系统NEE越大。光量子通量密度(Photosynthetic Photon Flux Density,PPFD)作为决定光合速率的重要因素之一,对生长季CO2通量的日变化有直接影响[16]。NEE随PPFD的增加而增加,当PPFD达到一定强度,生长季的叶面积指数(Leaf Area Index,LAI)增大也能导致NEE不断增加[17]。徐玲玲等[9]和石培礼等[18]也认为,生长季末白天生态系统的NEE主要受到光合有效辐射(PAR)变化的控制,PAR与LAI产生交互作用,共同调节光合速率和光合效率的强度,从而对GPP产生影响,最终影响生态系统的NEE。
2.2温度、昼夜温差与NEE的关系 青藏高原净初级生产力的分布决定于水热条件的共同作用[19]。温度是决定高寒草甸生态系统碳交换的最主要因素,对生态系统碳收支有重要影响[16]。草地生态系统GPP与气温变化呈正相关,由于青藏高原地区平均气温较低,植物生长期较短,GPP和地上生物量低于同纬度的低海拔地区。冷季NEE与温度密切相关,由于温度很低,微生物代谢活动受到抑制,CO2通量接近于零,暖季NEE与夜间温度变化呈显著的正相关[2,8,20]。一般情况,生态系统CO2日吸收量与昼夜温差变化呈正相关,生长季昼夜温差大有利于生态系统形成碳汇[21]。石培礼等[18]也认为,昼夜温差大不利于生态系统形成碳汇,生长季的温度波动导致较大的日较差,提高了呼吸日总量;生长季末植被的LAI降低,对CO2的吸收明显减弱,夜晚呼吸作用未显著减弱,表现为温差较大碳获取反而减少。温度从整体上影响植被生产力,决定了生态系统的固碳潜能,通过影响土壤的呼吸作用,控制生态系统碳的排放,可见温度是影响生态系统CO2交换量的重要因素。在全球变暖的形势下,对增温可能引起的草地生态系统碳循环的变化进行模拟也是当前国内外的研究热点。
2.3土壤温度、土壤水分和降水对NEE的影响 温度和湿度是影响高寒草地CO2通量的主要环境因子。Kato等[22]研究表明,矮嵩草(Kobresiahumilis)草甸土壤5 cm温度是影响CO2通量的主要因素,主要表现在土壤表层的融冻对CO2通量的影响。不考虑土壤水分条件时,土壤呼吸与土壤温度呈正相关,当水分条件和植被类型不同时,土壤呼吸则发生变化。土壤水分影响土壤呼吸,并对不同植被类型影响程度不同,土壤温度和水分是影响高寒草甸生态系统CO2释放量的主要因素[23]。王俊峰等[24-25]也发现,5 cm土壤温度和水分含量与CO2排放通量呈正相关,土壤温度与CO2排放通量显著相关,是控制CO2排放的主要环境因子。降水是高寒草地生长季的经常性事件[26],会对土壤温度和土壤含水量产生影响,从而影响生态系统的CO2通量。闫巍等[14]用涡度相关技术对高寒草甸生态系统CO2通量进行观测发现,生长季净CO2吸收量与降水量呈正相关。降水使草地生态系统的土壤微生物和植被根系呼吸增强,导致整个生态系统的CO2通量增加[27]。生长季初和生长季末的脉冲性降水会促进生态系统的碳排放,成为碳收支的决定因素[18]。积雪不但对青藏高原大气CO2浓度有显著影响,对其日变化也有明显的影响[28]。生态系统的碳源汇功能很大程度上受到年降水量、强度和季节分配的影响。
2.4叶面积指数、植被反射率对CO2通量的影响 生态系统光合能力与LAI呈显著的正相关,LAI高的生态系统有较高的CO2吸收速率和光合能力,在PFFD相同的条件下,生长季内CO2的净吸收量与LAI有相同的变化趋势。高寒草甸生态系统的LAI高于高寒灌丛,能吸收较多的光照,其固定CO2的能力高于灌丛[29]。青藏高原腹地生长期植被LAI较低,生长季内最高LAI为1.9,植物具有较低的光合能力和光合效率,致使生态系统的碳吸收能力低于世界大多数草地生态系统[18]。东北部地区高寒灌丛LAI约为2.6,生态系统净CO2吸收量也相对较高[8]。
青藏高原夏季主要地面的反射率为16%~28%,高于中纬度其他地区,冬季大面积的积雪会导致地面反射率增加,这样形成夏季巨大的热源作用和冬季巨大的冷源作用,通过辐散气流对毗邻地区及整个北半球和全球气候产生影响[26]。高寒灌丛和矮嵩草草甸CO2通量与植被反照率存在负相关关系,并随不同植被类型导致的群落结构变化而产生差异;而湿地NEE与气温、地表反射率等环境因素呈现相似的相关性,与地上生物量和群落叶面积指数呈线性负相关[21]。
2.5放牧和草地退化对高寒草地CO2通量的影响 放牧是高寒草地最重要的利用方式之一,合理的放牧利用有助于土壤碳的积累[30],而过度放牧是导致土壤有机碳释放量增加的原因之一[31]。放牧活动对碳平衡的影响主要表现为对地上生物量的直接采食和对根系生长的间接影响[32-33]。放牧直接减少地上生物量,通过影响地上碳进入土壤中再循环的速率,影响植被种类的分布和凋落物在土壤中的积累,从而影响碳在整个生态系统中的分布[23,34-36]。放牧导致植被和凋落物的变化可以改变土壤温度和湿度,土壤微生物数量增加[34,37-38]。放牧通过对草地土壤的物理结构与化学成分的影响,进而影响土壤呼吸[39]。对内蒙古锡林河流域羊草草原40年的研究发现,过度放牧使表层0~20 cm土壤的碳储量降低12.4%[40],可见,长期过度放牧加速了土壤有机碳向大气中的释放。另外,对草地的开垦也是影响草原土壤碳储量最剧烈的人类活动因素[41],长期对草地的过度利用可能使草地变成一个巨大的碳源[7,25,42]。而张金霞和曹广民[43]认为,高寒草甸CO2排放量总体上随着退化程度的加剧而逐渐减小。高寒湿地生态系统CO2排放强度也与其退化程度相关,整个生长季中,退化越严重,CO2排放通量越低[24];说明草地退化影响高寒草地的碳收支,严重的退化可能影响高寒草地生态系统的源汇潜能的变化。Cao等[44]对青藏高原东北部矮嵩草草甸进行放牧试验发现,重度放牧处理的土壤呼吸Q10值小于轻度放牧处理,即随着放牧程度的加剧,土壤呼吸对温度的升高响应变小。短期的放牧活动能够改变植物群落的物种组成,长期会导致生态系统碳分布格局的变化,放牧不仅直接改变土壤的呼吸速率,对土壤温度的影响也能间接地对土壤呼吸产生影响,从而影响整个系统的碳交换。
3 各类型草地的生态系统碳源汇潜能比较
在同样无放牧条件下,美洲北方大草原CO2净吸收量为125~209 g·m-2·a-1,长白山高山冻原为145 g·m-2·a-1,内蒙古羊草草原为126~195 g·m-2·a-1,而青藏高原高寒草地生态系统全年吸收量可达到227和282 g·m-2·a-1,表现为显著的碳汇(表1)。究其原因,植被生长季夜间温度明显低于白昼,昼夜温差大可能是青藏高原高寒灌丛有较高CO2净吸收的重要因素之一[8]。藏嵩草沼泽化草甸年CO2净排放量478 g·m-2,高于其他草地生态系统,表现为较明显的碳源。草原化嵩草草甸表现为较弱的碳汇,在降水减少的年份则表现为较弱的碳源[18]。
4 全球气候变暖对高寒草地生态系统CO2通量可能的影响
在全球变暖作用下,土壤呼吸加剧可能是对温室效应的正反馈作用[45]。Q10表示温度每升高10 ℃,土壤呼吸速率增加的倍数,在土壤呼吸测定中,常被用来表示土壤呼吸对温度变化的响应程度。一般认为,生态系统呼吸的Q10的平均值为2.4[45],即温度每升高10 ℃,土壤呼吸速率增加2.4倍;陆地生态系统土壤Q10平均幅度范围1.3~5.6[45-46]。随温度的升高,Q10逐渐减小,青藏高原高寒草地生态系统土壤的Q10也符合这一规律。由于青藏高原地区的年均温较低,其Q10较低海拔地区高(表2)。在低温地域,全球变暖对土壤有机质分解的加速作用 明显[47],因此,这一区域土壤呼吸对温度升高更加敏感。与全球其他地区相比,土壤呼吸释放大量CO2的潜力更大,对全球变暖的响应更加迅速和强烈。

表1 不同地区草地生态系统CO2年吸收量比较Table 1 Comparison of annual CO2 absorbance of grassland ecosystems in different regions
注:“-”表示净排放。
Note:“-”,net emission.
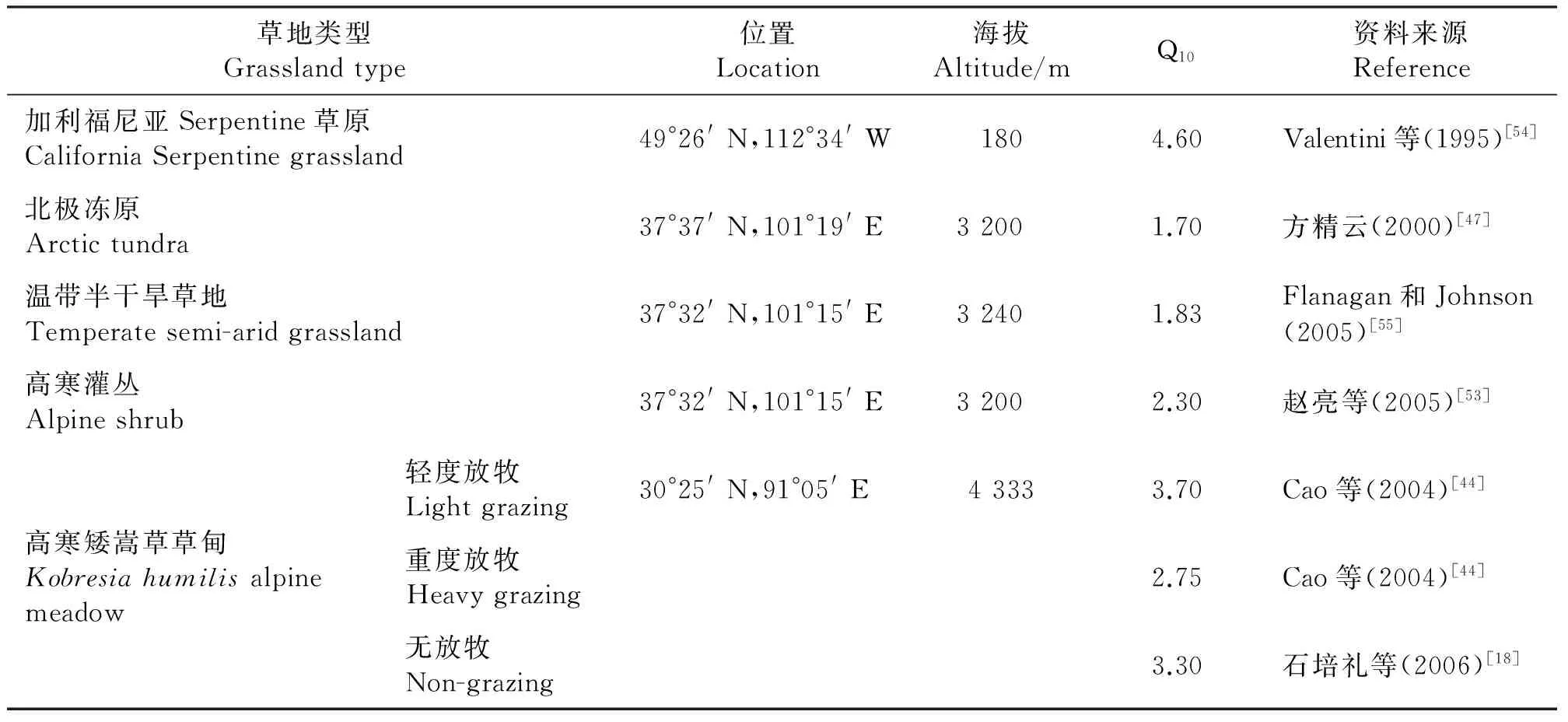
表2 各种草地生态系统对气候变暖的敏感性的比较Table 2 Comparison of sensitivity to climate warming of different grassland ecosystems
青藏高原各种草地类型有不同的碳源汇潜能,张永强等[56]的模拟结果显示,在不考虑土地利用变化,中等放牧强度的条件下,青藏高原草地土壤有机碳(0~20 cm)主要贮存于高寒草甸中。在全球变暖的背景下,高寒草地有较高的Q10,因此具有大量释放所储存的碳的潜能,可能减弱原有的碳汇潜能,而沼泽化草甸可能会转化为更加明显的碳源。方精云和郭兆迪[57]认为,近年来中国的灌丛生物量总体上呈现恢复的趋势,草地生物量碳库也在增加,因此,推断草地生态系统的土壤碳储量也在增加。
5 小结
目前,青藏高原各区域的高寒草地进行CO2通量研究已经积累了大量的结果,陆地生态系统碳循环过程中大气CO2源/汇强度及其时空分布特征的研究必须是在长期连续监测的基础上,尤其是在降水、温度等气候因素年际差异显著的青藏高原地区。所以,青海高寒草地生态系统CO2通量时间尺度上的长期延续研究非常必要。
当前,青藏高原碳储量的估算几乎都是建立在草地-土壤界面基础上,忽略了放牧家畜这一重要组成部分在草地生态系统碳循环中的作用。在草地-土壤界面系统框架下的草地生态系统碳循环研究不断深入的同时,考虑动物采食生物量,将放牧家畜的采食纳入草地生态系统碳循环研究才能全面、系统地诠释高寒草甸生态系统碳循环。
[1] Scurlock J M O,Hall D O.The global carbon sink: A grassland perspective[J].Global Change Biology,1998,4(2):229-233.
[2] Zhao L,Li Y,Gu S,etal.Carbon dioxide exchange between the atmosphere and an alpine shrubland meadow during the growing season on the Qinghai-Tibetan Plateau[J].Journal of Integrative Plant Biology,2005,47(3):271-282.
[3] Kato T,Tang Y,Gu S,etal.Carbon dioxide exchange between the atmosphere and an alpine meadow ecosystem on the Qinghai-Tibetan Plateau,China[J].Agricultural and forest meteorology,2004,124(1/2):121-134.
[4] 周兴民.中国嵩草草甸[M].北京:科学出版社,2001:17-21.
[5] 郑度,林振耀,张雪芹.青藏高原与全球环境变化研究进展[J].地学前缘,2002,9(1):95-102.
[6] 赵亮,古松,徐世晓,等.青藏高原高寒草甸生态系统碳通量特征及其控制因子[J].西北植物学报,2007,27(5):1054-1060.
[7] 于贵瑞,孙晓敏.陆地生态系统通量观测的原理与方法[M].北京:高等教育出版社,2006:10-12.
[8] Xu S,Zhao X,Li Y,etal.Diurnal and monthly variations of carbon dioxide flux in an alpine shrub on the inghai-Tibet Plateau[J].Chinese Science Bulletin,2005,50(6):539-543.
[9] 徐玲玲,张宪洲,石培礼,等.青藏高原高寒草甸生态系统净二氧化碳交换量特征[J].生态学报,2005,25(8):1948-1952.
[10] 徐世晓,赵亮,赵新全,等.青藏高原高寒灌丛非生长季节 CO2通量特征[J].西北植物学报,2006,26(12):2528-2532.
[11] 朱志鹍,马耀明,李茂善,等.珠穆朗玛峰北坡高寒草甸生态系统 CO2通量日变化与月变化特征[J].高原气象,2007,26(6):1300-1304.
[12] 徐世晓,赵新全,李英年,等.青藏高原高寒灌丛生长季和非生长季 CO2通量分析[J].中国科学:D 辑,2004,34(A02):118-124.
[13] 张法伟,刘安花,李英年,等.青藏高原高寒湿地生态系统 CO2通量[J].生态学报,2008,28(2):453-462.
[14] 闫巍,张宪洲,石培礼,等.青藏高原高寒草甸生态系统 CO2通量及其水分利用效率特征[J].自然资源学报,2006,21(5):756-767.
[15] Zhao L,Li Y,Zhao X,etal.Comparative study of the net exchange of CO2in 3 types of vegetation ecosystems on the Qinghai-Tibetan Plateau[J].Chinese Science Bulletin,2005,50(16):1767-1774.
[16] Saito M,Kato T,Tang Y.Temperature controls ecosystem CO2exchange of an alpine meadow on the northeastern Tibetan Plateau[J].Global Change Biology,2009,15(1):221-228.
[17] Zhao L,Li Y,Zhou H,etal.Diurnal,seasonal and annual variation in net ecosystem CO2exchange of an alpine shrubland on Qinghai-Tibetan plateau[J].Global Change Biology,2006,12(10):1940-1953.
[18] 石培礼,孙晓敏,徐玲玲,等.西藏高原草原化嵩草草甸生态系统 CO2净交换及其影响因子[J].中国科学:D 辑,2006,36(A01):194-203.
[19] 吕建华,季劲钧.青藏高原大气-植被相互作用的模拟试验Ⅱ.植被叶面积指数和净初级生产力[J].大气科学,2002,26(2):255-262.
[20] 徐世晓,赵亮,李英年,等.温度对青藏高原高寒灌丛 CO2通量日变化的影响[J].冰川冻土,2007,29(5):717-721.
[21] Gu S,Tang Y,Du M,etal.Short-term variation of CO2flux in relation to environmental controls in an alpine meadow on the Qinghai-Tibetan Plateau[J]. Journal of Geophysical Research-Atmospheres,2003,108(D21):4670.
[22] Kato T,Mitsuru H,Tang Y,etal.Strong temperature dependence and no moss photosynthesis in winter CO2flux for a Kobresia meadow on the Qinghai-Tibetan plateau[J].Soil Biology amp; Biochemistry,2005,37:1966-1969.
[23] 赵亮,徐世晓,李英年,等.青藏高原矮嵩草草甸和金露梅灌丛草甸 CO2通量变化与环境因子的关系[J].西北植物学报,2006,26(1):133-142.
[24] 王俊峰,王根绪,吴青柏.青藏高原腹地不同退化程度高寒沼泽草甸生长季节 CO2排放通量及其主要环境控制因子研究[J].冰川冻土,2008,30(3):408-414.
[25] 王俊峰,王根绪,王一博,等.青藏高原沼泽与高寒草甸草地退化对生长期 CO2排放的影响[J].科学通报,2007,52(13):1554-1560.
[26] 孙鸿烈,郑度.青藏高原形成演化与发展[M].上海:上海科学技术出版社,1996:264-266.
[27] 徐世晓,赵亮,李英年,等.降水对青藏高原高寒灌丛冷季 CO2通量的影响[J].水土保持学报,2007,21(3):193-195.
[28] 赵亮,徐世晓,伏玉玲,等.积雪对藏北高寒草甸CO2和水汽通量的影响[J].草地学报,2005,13(3):242-247.
[29] 李英年,王勤学,古松,等.高寒植被类型及其植物生产力的监测[J].地理学报,2004,59(1):40-48.
[30] 孔玉华,姚风军,鹏爽,等.不同利用方式下草地土壤碳积累及汇/源功能转换特征研究[J].草业学报,2010,27(4):40-45.
[31] 王根绪,程国栋.青藏高原草地土壤有机碳库及其全球意义[J].冰川冻土,2002,24(6):693-700.
[32] 李凌浩.锡林河流域一个放牧羊草群落中碳素平衡的初步估计[J].植物生态学报,2004,28(3):312-317.
[33] 宋智芳,安沙舟,孙宗玖.刈割和放牧条件下伊犁绢蒿生物量分配特点[J].草业科学,2009,26(12):118-123.
[34] Naeth M,Bailey A,Chanasyk D,etal.Water holding capacity of litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta[J].Journal of Range Management,1991,44(1):13-17.
[35] Haferkamp M R,Macneil M D.Grazing effects on carbon dynamics in the northern mixed-grass prairie[J].Environmental Management,2004,33:S462-S474.
[36] Li S G,Asanuma J,Eugster W,etal.Net ecosystem carbon dioxide exchange over grazed steppe in central Mongolia[J].Global Change Biology,2005,11(11):1941-1955.
[37] Hamilton III E,Frank D.Can plants stimulate soil microbes and their own nutrient supply? Evidence from a grazing tolerant grass[J].Ecology,2001,82(9):2397-2402.
[38] Lecain D,Morgan J,Schuman G,etal.Carbon exchange rates in grazed and ungrazed pastures of Wyoming[J].Journal of Range Management,2000,53(2):199-206.
[39] 郭明英,卫智军,运向军,等.放牧对草原土壤呼吸的影响[J].草业科学,2011,28(5):729-736.
[40] Li L,Chen Z,Wang Q.Changes in soil carbon storage due to over-grazing inLeymuschinensisSteppe in the Xilin River Basin of Inner Mogolia[J].Journal of Environment Sciences,1997,9(4):486-490.
[41] 周萍,刘国彬,薛萐.草地生态系统土壤呼吸及其影响因素研究进展[J].草业学报,2009,18(2):184-193.
[42] 钟华平,樊江文,于贵瑞,等.草地生态系统碳蓄积的研究进展[J].草业科学,2005,22(1):4-11.
[43] 张金霞,曹广民.放牧强度对高寒灌丛草甸土壤 CO2释速率的影响[J].草地学报,2001,9(3):183-190.
[44] Cao G,Tang Y,Mo W,etal.Grazing intensity alters soil respiration in an alpine meadow on the Tibetan plateau[J].Soil Biology and Biochemistry,2004,36(2):237-243.
[45] Raich J,Schlesinger W.The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate[J].Tellus,1992,44(2):81-99.
[46] Tjoelker M,Oleksyn J,Reich P.Modelling respiration of vegetation:Evidence for a general temperature-dependent Q10[J].Global Change Biology,2001,7(2):223-230.
[47] 方精云.全球生态学:气候变化与生态响应[M].北京:高等教育出版社,2000:156-160.
[48] 吴钢,邵国凡.长白山高山冻原生态系统碳循环及与北极对比研究[J].中国科学:D辑,2001,31(12):1039-1045.
[49] 陈佐忠,汪诗平.中国典型草原生态系统[M].北京:科学出版社,2000:112-118.
[50] 方精云,位梦华.北极陆地生态系统的碳循环与全球温暖化[J].贵州科学,1998,18(2):113-121.
[51] Frank A,Dugas W.Carbon dioxide fluxes over a northern,semiarid,mixed-grass prairie[J].Agricultural and Forest Meteorology,2001,108(4):317-326.
[52] Frank A.Carbon dioxide fluxes over a grazed prairie and seeded pasture in the Northern Great Plains[J].Environmental Pollution,2002,116(3):397-403.
[53] 赵亮,李英年,赵新全,等.青藏高原3种植被类型净生态系统CO2交换量的比较[J].科学通报,2005,50(9):926-932.
[54] Valentini R,Gamon J,Field C.Ecosystem gas exchange in a California grassland:Seasonal patterns and implications for scaling[J].Ecology,1995,76(6):1940-1952.
[55] Flanagan L,Johnson B.Interacting effects of temperature,soil moisture and plant biomass production on ecosystem respiration in a northern temperate grassland[J].Agricultural and forest meteorology,2005, 130(3/4):237-253.
[56] 张永强,唐艳鸿,姜杰.青藏高原草地生态系统土壤有机碳动态特征[J].中国科学:D辑,2006,36(12):1140-1147.
[57] 方精云,郭兆迪.寻找失去的陆地碳汇[J].自然杂志,2007,29(1):1-6.
AdvancesoncarbondioxidefluxesofalpinemeadowecosystemsontheTibetanPlateau
QIAO Chun-lian1,2,3, LI Jing-mei1,2, WANG Ji-heng1,2, GE Shi-dong1,2, ZHAO Liang1,3, XU Shi-xiao1,3
(1.Northwest Plateau Institute of Biology, CAS, Xining 810001, China; 2.Graduate School of Chinese Academic of Science, Beijing 100049, China; 3.Key Laboratory of Adaptation and Evolution of Plateau Biota, Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810008, China)
Alpine meadow is the key vegetation widely distributed on the Tibetan Plateau. Alpine ecosystem is a great important part in biogeochemical cycle between air and the ground surface, and it plays an extremely important role in carbon balance in this area. Advances on CO2fluxes of alpine ecosystem were reviewed. The variations of carbon dioxide fluxes of alpine meadow ecosystem and its effecting factors were summarized on the Tibetan Plateau. Furthermore, the potential of source/sink and value of Q10were compared between typical alpine meadow ecosystems and others. More specifically, some unsolved questions which need to be answered in the future were raised in this review.
Tibetan Plateau; CO2flux; net ecosystem CO2exchange; temperature
XU Shi-xiao E-mail:xushixiaoqh@126.com
S812.2
A
1001-0629(2012)02-0204-07
2011-05-25 接受日期:2011-07-24
国家自然基金面上项目(30970519、30770419);中国科学院西部行动计划项目(KZCX2-XB2-06)
乔春连(1985-),女,山东青岛人,在读硕士生,主要从事区域碳循环研究。E-mail:qiaochunlian2008@126.com
徐世晓 E-mail:xushixiaoqh@126.com
