我国竹类植物种质保护体系的适度理念研究
2012-12-06何亚平张翠微高志民李本祥费世民蒋俊明
何亚平 张翠微 高志民 李本祥 费世民 蒋俊明
(1 四川林业科学研究院 成都 610081 2 国际竹藤网络中心 北京100102 3 长宁县林业局 四川长宁 614000)
竹类是禾本科(Gr a min ea e)竹亚科(Bambusoideae)植物的总称,由于其形态、结构和生长发育等方面的特殊性,成为植物分类系统中较为复杂的一大类群,也是生物多样性体系中的重要环节与谱系分支。从构型上考虑,植物存在连续分支与重复性生长骨架与易变的连接骨架2类结构,前者是一般被子植物类群,后者是禾本科植物[1]。某些禾本科植物还具有基部多重分蘖的习性,从土壤基质或地表构建个体构型,形成一年生、多年生或木本构型。全世界共有竹类植物107个属1 300多种,其中木本竹类植物79个属1 200多种,草本竹类植物28个属180余种。据《中国植物志(第九卷第一分册)》记载,中国有竹类植物39个属(其中新属及中国新分布属共19个)509种,属数和种数分别占世界总数的36%和39%。全球共有竹林面积2 000 万hm2以上,约占世界森林总面积的10%,中国的竹林面积约500 万hm2,约占全国森林总面积的4%,占全球竹林面积的25%,是世界上竹类资源最丰富的地区之一[2,3]。
1 竹类植物的分布与繁殖特性
1.1 竹类植物的分布特性
竹类起源于热带但仍有很强的适应性,从热带至温带都有分布,大多数种类集中于热带和亚热带湿润而温暖地区,包括中国在内的东南亚季风区是竹类植物的中心分布区。海南竹类区系具特有成分丰富,热带性明显,分布异质,原始古老等特点[4]。
竹类资源受季风气候影响,主要分布在水热条件较好的热带、亚热带地区,少数种类可生长在温带或亚寒带地区,其水平分布南到拉丁美洲的阿根廷南部(47°S),北至亚洲库页岛(49°N) ;垂直分布,亚洲的高山竹种可达海拔3 800 m,拉丁美洲的高山竹种分布到海拔4 000 m,有少数种类可达雪线或雪线以上,最高记录达4 500 m,成为世界竹类植物分布海拔最高的地区。
从世界范围来看,竹类的地理分布可分为3大区域,即亚太竹区、美洲竹区和非洲竹区。根据狄尔斯的世界植物区划原则,世界竹类分布分为4大区系,即泛北极竹区、古热带竹区、新热带竹区和澳洲竹区。中国的竹类植物种质资源自然分布以云南最为丰富,有26个属220余种,东南沿海的广东、浙江、福建、台湾等省区也较多,内地则以四川、江西、湖南、广西、贵州等地为主。中国竹林水平分布的北界在北纬40°的辽东半岛、北京一线,华中和华东及其以北地区主要以单轴散生竹类植物为主,华南和西南地区则以合轴丛生竹类植物为主。竹林的垂直分布可以从沿海至海拔4 000 m以上的高山[5]。
中国竹类植物从分布上看也具有明显的地带性和区域性,可划分为北方散生竹区、江南混合竹区、西南高山竹区、南方丛生竹区和琼滇攀援竹区[6]。从竹林经营的角度来研究竹林的区划,根据影响竹子分布的降水、气温、季风等主导生态因子,中国竹林可划分为如下类型:亚高山竹林带、散生竹林带(降水性散生竹林带和灌溉性散生竹林带)、混生竹林带、丛生竹林带(东南季风丛生竹林带和西南季风丛生竹林带)[7]。我国竹类分布不但在水平分布上具有明显的南北差异,而且在垂直分布上也表现出明显的不同[8]。云南、四川、浙江、贵州、广东等省竹类资源丰富多样且分布区类型较为复杂,都形成了各行政区划背景下的竹林的生态地理区划[9-13]。
竹子是一类亚热带起源的植物,对气候变化非常敏感,是研究气候变迁的指示物种。在竹类植物分布的历史上,随着气候变化与人为干扰,其分布范围呈现萎缩状态。300万年前的三叠纪,竹类植物在欧洲也有分布,但由于冰期而萎缩[14]。唐代以前,黄河流域有较多的竹类分布,北界可达北纬39°~40°,而今天为北纬35°,往南退缩了4°~5°;中国历史上,年均气温每下降1 ℃,竹类分布北界则南退约1N,反映了历史上气候的变化;且中国“竹子文明的国度”与历史分布更为广泛有关[15]。
1.2 竹类无性系种群特点
竹类植物分支格局通过地下茎构件发育来实现,并构建明显空间格局的无性系种群。竹类植物按其地下茎结构及竹秆群集程度分为合轴丛生、单轴散生和复轴混生3大类,反映了不同的构型特征与觅养行为,其生长格局主要对应于扩展觅养型、稳定觅养型与稳定扩展复合觅养型3种类型:①扩展觅养型的隔离者(地下茎或竹鞭)长度较长,向外扩展能力很强,子代无性系植株与父代无性系植株在空间尺度上相距较远,生态位重叠度偏低,能充分利用空间资源,无性系种群规模形成速度快,生态适应性较强,生理特性为喜阳植物,无性系种群呈成片状均匀分布;②稳定觅养型地下茎长度很短,仅向母竹周围扩展,子代与父代植株在空间上相距较近,生态位重叠度较高,生态适应性偏低,生理特性属耐荫植物,种群规模形成速度慢,种群间断聚集分布利用空间那资源不够充分;③稳定扩展觅养复合型隔离者既具有扩展觅养型的长竹鞭,又生长着稳定扩展格局的段根茎,在种群形成的初期,主要为稳定扩展格局;当达到一定生长年龄阶段(指基株种群年龄)后,分化形成部分具有扩展觅养特征的地下茎[16]。
合轴丛生型竹类植物较喜暖热,而单轴散生型竹类植物则较耐寒,复轴混生型竹类植物对热量的需求介于2者之间。在世界范围内,印度、泰国、印度尼西亚、菲律宾等国家及南美诸国主要分布合轴丛生竹;而日本、朝鲜等国家及北美洲主要分布单轴散生竹;与中国的热带、亚热带同纬度的区域,往西为北非及中亚荒漠。往东则为太平洋海域,只有在中国合轴丛生、单轴散生和复轴混生3大类兼有分布。从竹类热带起源来看,散生竹类由于是较为进化的类型,适应能力强,对不良气候及环境的抗性强,因而其分布范围也更广泛。
竹类构型特性与异质性资源的可利用性有关,克隆格局形成的适应假说能解释类型差异。竹类克隆生长格局与生态存在协同进化关系,同时受制于植物遗传机制的影响,其隔离者的数量特征(长度、基径)是无性系植物种群觅食行为的重要参数,决定者该种群的生长格局与资源利用潜力。随着生境可利用性减少,隔离者长度则可能增加,隔离者数目可能减少;相应地,克隆构型也发生从丛生型向散生型过渡,生境水热协调性减弱,间隔子延长。这种克隆构型间演化关系,需要从生长格局对有性生殖过程逐渐调控中实现的,并经历了复杂而漫长的自然选择。
对许多多年生克隆植物来说,大量的研究表明,光是限制因子时,随着立地密度的不断增加,克隆分株的出生率逐渐减小、死亡率逐渐增加。由于竹类克隆生长的生理整合功能,随着立竹度增加,出笋数、活笋数等明显呈现增加趋势,竹笋生长过程很大程度上不需要光照[17]。这种情形发生在母竹光能利用充足的条件下,在密度制约显著时,影响了母竹的光合作用与自我营养能力时,直接会影响基株资源配置格局,会发生从低龄向大龄偏移,分株枝叶减少且基株克隆分株或发笋数、存活数减少。自然条件下,慈竹的密度与土壤厚度有关,高度、密度与海拔呈正相关,笋逐日生长量与湿度、温度、降雨量有密切关系,出笋数与土厚有关,退笋数受到母竹高度和出笋数制约[18]。无性系形态可塑性反映与能量投资对策对水分适应性明显,总生物量、分株数量分别随着水分供应呈现增大与减小趋势,间隔子长度与分株大小都呈现显著差异,低水分能量投资于根茎伸长而高水分时能量主要投资于分株的生长[19]。
空间分布格局是无性系植物适应环境和分株之间相互作用的结果。随着时间的推移,无性系不断产生新的分株.其分布格局必然是一个动态的过程,目前主要用扩散模型和相关随机走动模型来描述无性系的克隆生长过程。斑苦竹无性系采用了“倒逐龄级累加法”(RAA)度量生长格局动态与分株生长型的动态趋势,种群随着时间发育表现为逐渐降低的聚集性分布格局[20]。生境不同,斑苦竹表现出不同的适应特征,自然林斑苦竹无性系种群类似于k-对策种群,成为A型;人工林斑苦竹无性系种群类似于r-对策种群,称为B型,不稳定多变生境系统利于枝叶发展[21]。
1.3 竹类植物有性生殖特点
竹类植物生长达到花熟状态,营养叶的发生受到抑制,小枝上的顶芽或侧芽从营养生长锥转变为生殖生长锥,成为花的分生组织,继而发育成花芽、花穗,直至花的开放。竹类植物花芽的形成到开花所需时间因竹种而异:一些竹种需要跨越2个年度,有的竹种则当年形成花芽并开花,历时仅需2~3个月时间。竹类植物的花属于颖花,雄蕊的伸出作为花开放的特征。竹类植物的花期因竹种而异,可分为长花期和短花期。长花期是指一年有2次或2次以上的盛花期,全年均有零星不规则开花的竹种;短花期是指一年1次盛花期,花期相对较集中的竹种。因为花期受环境因子,特别是受温湿度的影响较大,即使有的长花期竹种也会有开花休止的时候。开花竹分为有叶型与无叶型,有叶型花穗基部带绿叶,是由枝端的分生组织演变而成,无叶型花穗则是由节间分生组织发育而成。日本有的学者则将半面开花以及部分不开花的秆看作一种嵌合体。
绝大多数的竹类在经历一个周期(约30~120年),一生只开1次花,结实后枯死,而后又以种子萌发成幼苗,经过无性繁殖阶段,再进入有性繁殖期,使种族更新繁衍。秦自生等 (1993)对冷箭竹进行过较为详细的研究,冷箭竹的开花周期约为45~55年,开花期一般为5~6月,花开后持续6天时间而枯萎,由于开花期内植食性昆虫和草食动物的破坏,箭竹种子产量较低,种子相对较重,常落在开花后的母株下以保持原来的领地,母株开花结实后枯死有利于种群的更新(秦自生等,1995)。观测表明,竹子的开花结实现象可以分为全体成片开花、零星开花;开花至死、开花不死、两者并存;开花后结实、开花后不结实等类型,竹子的开花、结实类型与竹种是否为野生种或栽培种密切相关,也与属级分类群紧密相关[22]。大多数竹子靠风媒传粉,仅热带几种草本竹类是靠昆虫传粉;最近发现,蜜蜂在一些竹类进行风媒传粉的过程中起到间接的帮助作用,表明传粉的有效性在试图解释竹类的大面积开花是可以考虑的(Huang等,2002)。对于开花原因的假说主要有周期学说、环境诱因学说、地震学说等(王微等,2006;陈嵘,1984;温太辉,1965;裘福根,1992)。据对安吉竹种园21种开花竹种的观察,认为竹子开花结实是从营养生长转变到生殖生长的生理过程,其历时因竹种不同而有显著差异,且环境对延缓或促进开花具有显著的影响,也可用周期说、环境说共同作用来解释开花现象[24]。余群洲等[25]用周期说、环境说、碳氮比说解释了卧龙箭竹的开花现象。竹子开花内因是关键,是生理成熟后的产物,环境因素只是加速或延迟开花过程[22]。
竹类普遍存在花而不实现象,雄性不育可能是主要原因。竹类植物开花后结实率极低,传粉限制、资源限制、物理伤害、病虫害等都可能解释结实率低下。这些都表现在开花过程中,无鲜艳花瓣、气味,无法吸引传粉昆虫;竹类植物分布在季风气候区,空气湿润,散粉不宜,受精作用不良,结实率低。竹类种群开花、结实机理是当前保护生物学研究中最具争议性和挑战性的议题之一。麻竹结实率低用传粉有效性、雌雄异熟的花器官构造、花粉活力低(5.4%~40.4%)来解释[26]。花药合并黏连型、无花药型,存在扁败型、圆败型、浅染型等花粉败育类型,花粉败育、活力低下、授粉率低与花粉花柱发育不良可能也是五月季竹结实率低的主要原因[27]。
2 竹类物种基因库维持的种群结构度
2.1 竹类种质基因库保存的小种群问题
竹类种质资源多样性及其基因库保持主要表现为野生种质多样性的维持与演化,但往往竹类种质在许多因素的影响下而发生退化,未有适宜该种类物种基因库保存的基因维持条件。中国有39属500多个竹种,丰富的竹类植物种质资源多样性形成了丰富的遗传多样性,同时决定了不同竹种具有各异的性状[5],即丰富的竹类植物性状多样性,也决定了不同竹种具有不同的生态功能和不同的外部景观,形成种群、群落与生态系统多样性。
生命系统是一个等级系统,包括多个层次或水平(基因、细胞、组织、器官、构件、种群、群落、生态系统),但有性生殖的种群单元系统,即集合种群才是基因库的保存、维持与演化载体。单一局域居群容易灭绝,物种往往以多个空间尺度下的异质性种群的形式存在,并伴随着生态系统内部斑块格局演变而发生时间变化与空间迁移。但受到人为干扰影响,竹类种质赖以生存的生境改变、破碎化导致很多竹种种群个体数量急剧减少,进而发生近交与近交衰退,逐渐丧失原有基因库,形成小种群。小种群除了易受环境、种群统计学因素影响而增大灭绝风险外,小种群遗传多样性丧失也影响到后代的适应性;小种群的形成可能与适宜生境的缺乏、生境的承载力有限、生境处于非稳定的状态(如演替中),以及种群入侵新生境(未扩张)有关[28]。
物种维持可能存在一个必备的集合种群系统,需要局域种群保持一定的物种数量,以利于基因交流,种质基因库传递与适宜种群数量的维持与扩展。适宜种群规模,种群与集合种群数量可能决定着物种不同的演化命运,但这些种命运从现实种群数量与分布能反映出来(图1)。竹类种质基因库赖以保存的个体数量、生殖能力、种群更新能力与现实种群的维持能力都直接影响着种群与物种的命运,在多种因素都促成生存适合度下降时,小种群成为必然归宿与趋势,随之而来的基因交流受限、近交发生与种群稳定遗传潜能下降而逐渐减少基因库,且长期后处于病危与趋于灭绝状态。
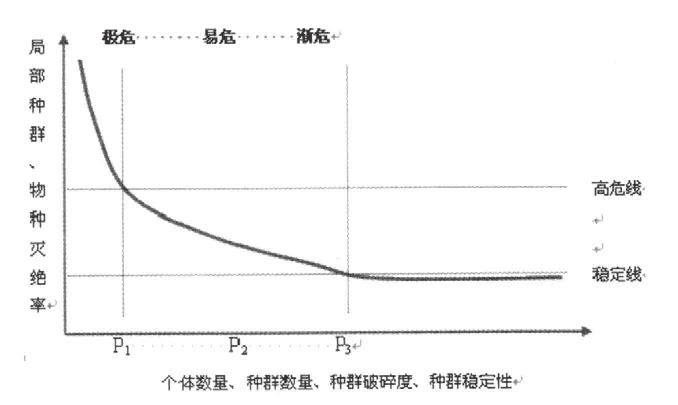
图1 个体数量居群规模与分布和物种灭绝的关系
2.2 竹类种质基因库的种群结构效应
图1描述了不同种群数量、个体数量、种群破碎度与种群稳定性条件下,局部种群与物种灭绝概率。显然,在种群个体数量、种群数量、种群离散性与种群稳定性趋近于或低于一定阈值时(P3、P2、P1),物种或局部种群处于渐危、易危与濒危状态,灭绝风险逐渐从稳定线上升至高危险。物种稳定维持与演化需要一个适宜的种群结构,包括种群规模、种群分布、种群数量与离散性,以及种群自身的稳定性,在上述这些因素都趋于高值时,以集合种群形式存在的物种才能稳定维持与演化,其本身固有的基因库能得到保存。物种存在一个适宜的种群结构度,物种基因库存在一个基因交流与传递的种群结构度,并且这种种群大于某一结构阈值时利于物种的维持,否则逐渐从渐危进入易危、极危后逐渐不利于基因库的保存。
实际上,基因库以存活个体形式保存,种群数量直接影响到基因交流与杂合性、种群扩散与维持演化能力,现实种群状况也形成了基因库的间接表征指标。往往一个种群的现实结构特征也是群落环境状况与立地条件综合作用的产物,野生濒危竹类种群的群落学及其人为干扰可能是判断种群结构度状况与维持性的基本方法。小种群近交衰退研究支持了竹类物种基因库保存与维持存在一个适宜的种群结构度。小种群遗传变异较小,稀有物种的种群结构度明显较低,遗传变异性要低于广布物种。濒危竹类类似于岛屿物种,岛屿物种遗传多样性低于大陆物种。物种近交衰退与生殖世代之间呈现指数增长:F=1-[1-1/(2Ne)]t,表明有效生殖个体数量越小,F值越明显,当F小于1而大于0时,表示和随机交配相比,近交产生更多的纯合个体,越大,种群近交程度越高;当其值在-1与0之间时,表示和随机交配性比,远交产生更多的杂合个体;其中Ne为有效种群大小,t为世代数量[28]。
长期近交,有2种效果预测,一种是显性假说(Dominance hypothesis),即近交增加了有害等位基因纯合度几率,导致了大多数有害等位基因因纯合而淘汰,种群进一步近交将不会导致适合度的进一步下降(约50个世代);超显性假说(Overdorminance hypothesis),认为种群的进一步近交将导致杂合度的进一步降低,还将导致种群适应性进一步下降[28]。应对多变环境,竹类种质遗传多样性较高时能增加适应性,提高生存适合度,单纯的纯合基因能保持瞬间生存适合度,但对于物种对长期多变环境的适应可塑性则不能保持。近交对竹类种质长期存活的风险极为明显,不同种群大小与世代条件下,一对等位基因交配后的近交系数变化如图2所示。显然,在种群数量从1 000向10个的种群衰退过程中,随着交配世代的延伸,线性斜率明显增加,表示近交系数增长逐渐加剧,种质适应未来多变生存环境的潜能极度下降,且F趋近于1的交配世代提前,暗示物种遭受随机环境灭绝的几率大大增加。图1试图从物种层面上揭示灭绝的结构性因素,增加了种群结构以外的个体数量、种群数量、种群破碎度和种群稳定性,来统一表示集合种群形式存在物种的局域种群数量、分布特性,来增加物种灭绝与基因库丢失的种群或物种结构度描述的普遍性。

图2 一对等位基因在不同种群大小和世代下的近交系数(引自王峥峰等,2005)
2.3 竹类种质种群结构度的影响因素
诸多因素都会影响到竹类种群的数量与分布特征。竹类物种都为依附性生物体,未能构建顶级群落,都是以小乔木、灌木或草本形式构建乔木优势种类的亚层,且光照不充分,水分相对充足,长期利用散射光来完成存活与无性系种群建立所需要的光合同化作用。但竹类植物长期稳定演化,其机理可能归结为水分对光照的补偿,生理整合的克隆繁殖,并在优势乔木丧失优势或林窗形成时借助强光照与林内干燥进行有性生殖,并完成基因交流与基因库的种群替代。
现有人类活动与土地垦殖条件下,竹类植物遭受的种群威胁主要源自于以下几点:①土地经营活动的调整,从保护走向开发或从竹林走向建设用地、农业用地;②水热条件良好的竹类分布区的土地利用格局调整引发生境破碎化,导致了竹林从规模化走向片段化与小种群,完全阻断了局域种群间基因交流与广泛随机交配的发生;③竹林经营活动直接增强了无性系种群生存的营养性,而杜绝或降低了有性生殖的概率,使得竹林长期处于一种克隆维持状态,而积累有害基因而种质得不到基因补充改良;④人工引种栽培改变了野生竹林的种群环境,离散而大量的园林用途使得竹类名义上保存了物种基因库,但长期维持风险依然受到美学欣赏的改变而增加;⑤竹类植物本身的华而不实而多花少实的生殖限制特性也是基因库保存的不利因素,但这种局部开花灭绝现象之所以危害大根源于总体该竹类种群规模小而种群少有关,是大尺度上竹林赖以发育的生境生态系统发生了蜕变所致,是集合种群结构度退化的产物;⑥竹类种质分布区无序牧业、无序采挖竹笋、竹材,以及森林破坏导致竹类赖以生存的生境遭受损毁,干扰了竹类长期维持与演化的生活史条件,森林自然演替格局打乱,竹类生活史发生响应而不利于种群扩张与生存维持。上述各类因素在现有生态发展条件下将长期存在,竹类种群基因库保存需要建立和完善现有野生与人工栽培状态下的竹类种质群体,建立依照物种基因库结构度的适宜性来确保竹类种质保存的管理体系,以保障竹类特殊种质基因库在保障经济社会发展背景下的竹类种质稳定维持与竹类多样性的持续演化,保护竹类资源持续利用的种质基因库。
3 竹类种质保护的适度思想
竹类分布广泛,历史上分布范围变动明显,竹类种质赖以生存的自然生态系统受到人口剧增的影响,而发生了剧烈的破碎化,不但使得竹类种群分布范围狭窄化,还使得种群规模数量发生衰减,因近交衰退而使得基因库发生减少而丧失长期适应性。随着交配世代延长,竹类种质及其赖以生存的生态系统都发生了类似的物种基因库衰减,从而总体竹林区域基因库都发生大幅度不可逆减少,进而丧失了区域全面、协调与可持续发展的生态支撑力。总体看来,竹林区域生态系统结构现状与经济可持续发展密切相关,也表明竹类种质保存现状与总体生态系统背景现状休戚与共、协同共生。
3.1 开展竹类多样性保存原因
竹类植物是野生动物,特别是珍稀动物的食物来源和栖息地,其种质资源也是遗传改良的基本材料,是人类安居乐业所依赖的重要种质资源之一。珍稀竹种及其保护研究是竹类多样性研究的重要内容,此方面的研究目前显得十分薄弱,表现在:“中国珍稀濒危保护植物名录”(1985)、“国家重点保护野生植物名录(第一批)”(1999)列出了国家级珍稀濒危保护植物389种,其中仅包括竹亚科4种。据我国“生物多样性国情研究报告”,濒危及稀有竹类植物达12种,但实际上特产中国的竹类种质资源以及稀有、濒危竹类植物有上百种,更多的处于濒危状况而经济价值极高的竹种尚未被认识。刚竹属竹类植物基本上原产中国,植物志上列出50种,33变种、变型,近年来发表的新种达26种,变种、变型24个,全部为中国特产,其中有具重要观赏价值的黄纹竹、黄秆乌哺鸡、花秆早竹、黄棺斑竹和黄秆京竹等,有具重要材用或笋用价值的紫蒲头灰竹、尖头育、安吉金竹、摘竹、巨县红壳竹、芽竹以及云和哺鸡竹等都分布极为有限,处于濒危状态。特产中国的俊形竹具有很高观赏价值,现已几近灭绝,仅安吉竹种园早年有引种得以保存。
在生境压力持续并逐渐加强的情况下,诸多竹种的小种群呈现破碎化分布,并与农户承包经营农林业用地灵活性相关,保护区设立受到林地权属、种群维持历史与土地破碎现状的限制,其保护作用受到严重影响,竹类野生种群长期处于恢复微弱而威胁持续的状态。竹类种质利用依靠野生种质人工繁育而栽培,或者直接改造野生资源为人工经营林地,往往由于人口分布范围宽,密度大,土地垦殖强度大,除了高山峡谷地带外,几乎都为人为土地利用所覆盖。热带、亚热带丛生竹大量以四旁分布为主体,一些成块野生竹林都被改造为纯林高产经营,长期处于强度砍伐而营养生长状态,是无法自然延续的基因库,仅仅是种质抢救性保存,种群维持完全依靠人为栽植与经营活动。
群体开花死亡对珍稀竹种种质资源的威胁、不良经营方式对珍稀竹种造成的破坏值得注意。研究的重点应放在发掘尚存的种质资源并及时加以引种保存,从保护生物学的角度搞清其生物学特性、生态适应性及生殖发育机制,采用组织培养等手段进行扩大繁殖及开发利用等。建立自然保护区开展就地保护,能使丰富的竹类种质资源在其天然生长地得到有效保护,保护区内因人为因素导致对珍稀竹种资源的威胁已得到有效控制,而因自然因素导致的威胁尚缺乏深入研究和有效控制。
3.2 竹类种质保护的适度思想
保护适度强调的是依照竹类野生种质保存现状来确定保护的对策,具体说就是依据野生集合种群数量来划分物种趋于灭绝的程度,进而确定合理的资源利用与保护途径。这种思路与传统珍稀濒危竹类种质保护不同之处在于,依照竹林生态系统景观多样性与竹类种质的集合种群现状来确定合理的保护对策。例如,有些竹类种质的种群数量相对较多,但是分布零散,集合种群数量多而局域种群沦为小种群或低密度种群;有些竹类种质的局域种群发达,种群数量大,但集合种群数量少,竹林生态系统的景观多样性较低。前者总体多数沦为灭绝的风险大,而后者局域种群因灾变而沦为灭绝的风险较大,但前者在局域种群密度增加,后者在于局域种群数量增加时,整体物种灭绝风险降低。用物种分布与种群数量同时进行濒危等级划分,进而确定保护区域与保护种类优先性,是保护生物学研究的基本思路。
通过构建单一物种集合种群结构与竹类种质基因库的定性关系模型发现,一个物种基因库是由很多集合种群的基因库构成的,而集合种群则是由很多局域种群基因库构成的。遗传多样性研究表明:遗传多样性贡献可划分为个体内、居群内、居群间占优3种类型,分别表明个体、种群与集合种群在物种基因库中的贡献份额高低,个体占优表示一个个体能代表绝大多数基因库基因份额,居群则表示居群对物种基因库的代表性,居群间则表明集合种群才能代表物种基因库。并随着集合种群数量增加,则整个物种基因库的组成增加而基因总量也随之增加。图3表明,在集合或局域种群数量从P0增加到P1、P2、P3、P4时,物种基因库分别从G1增加到G2、G3、G4;显然,对于一个物种而言,科学的保护则需要多个集合种群,且每个集合种群都由若干个局域种群组成,每个局域种群都处于小种群以上的种群结构。若以物种整个基因保护为目标,则大部分G4基因库水平的P4种群结构度需要满足,那就是所有分布区的大部分野生种群;但往往部分种群就能代表物种基因库的95%以上,但这些种群分布与数量可能很难量化,一些起源性质的,往往很萎缩,而演化状态的往往很发达,且起源地基因库也在丢失当中,很多树种沦为低密度种群。竹类种质依照区域生态系统来划定珍稀濒危等级是合理的,科学区划要以地理气候与自然演化格局作为依据。从单一竹种基因库与种群结构度模型可知,我们需要一个适宜大小的、气候决定的竹林生态系统地理范围,在决定了该范围内的竹类种质稀有程度与种群结构度,进而确定保护方案是科学合理的,即确立一个生态系统结构度上限。在适宜上限基础上,确定适宜的集合种群结构度、局域种群结构度,进而采取科学的保护策略。
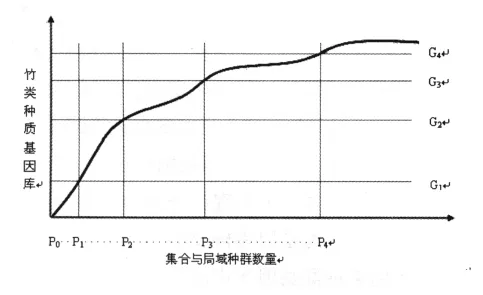
图3 单一竹类物种在集合与局域种群结构度增加过程中的基因库变化
3.3 基于生态系统结构度的竹类适度保护
基于集合种群与物种基因库关系,划定一个气候决定的生态系统结构度上限,进而作为确定物种珍稀濒危的范围。但这一生态系统结构度是有若干个子系统构成的,子系统的结构大小与数量分布直接影响了总体的生态系统结构。竹类物种稳定性与生态系统结构度增减存在明显的联系,这种关系可用图4定性模型来表述。

图4 基于生态系统结构度的竹类物种稳定性模型
当区域生态系统作为一个封闭系统时,其竹类种质稳定性受到生态系统的影响;对于生态系统而言,越大越稳定。用单一竹类物种集合种群来描述区域生态系统结构度内的物种稳定性与结构度大小的变化关系。这一区域物种总体或亚物种基因库是所有种群或一个集合种群基因库组成的,局域种群稳定性随着局域种群规模增加而增加,局域种群规模受到生态系统结构度增加的促进,即转化为生态系统结构度增加时,竹类物种稳定性增加。图4表明,在生态系统大小从Ea0增加到Ea1、Ea2、Ea3、Ea4时,局域种群承载力越大则基因库发生从0、S1、S2、S3到S4,物种稳定性呈现增加趋势。在自然无干扰的封闭生态系统中,物种稳定性则依据种群数量分布,物种呈现恢复状态,则期稳定性增强,而呈现退化状态在,则其稳定性减弱。在集合种群尺度,生态系统背景下,种群表述直接依照景观多样性来计量,将竹类种群分布斑块作为景观斑块单元来划分,确立一个集合种群度量的方法。例如区域内,竹类种质斑块可划分为20块,则有20个局域种群,共同组成一个集合种群,往往生态系统结构度从1到20的增加过程中,竹类种质稳定性则从S1增加到S4。显然,生态系统结构度低于某一阈值,则集合种群数量不足,基因库代表性与物种稳定性不保,保护效果是值得商榷的。
基于生态系统,则竹类种质适度保护主要从局域种群斑块数量来作为保护的局域种群单位,并对局域种群结构数量进行适宜优化评估。在局域种群数量少时,生态系统自身的内部循环演替会导致局域种群灭绝的概率增加,整体集合种群存留率降低,区域物种灭绝风险增加;而在局域种群数量增加时,能提高区域物种保存率与稳定性,在局域种群增加超过一定的度时,局域物种基因库代表性上限达到了,多余局域种群数量显得冗余。生物多样性有冗余假说,这种物种冗余实际上在种下居群水平也有体现。尽管如此,但不能用区域冗余来放弃另外其余的同类竹类种质的保护,物种基因库是基于地理气候种源基础上的遗传结构的长期演化所致,具有地理独特性,这种独特性需要多个区域生态系统共同保护才能实现物种基因库的保护需求。对于生态系统尺度上的集合种群保护而言,适度就是要依照局域种群数量与区域种质基因库代表性之间关系来确定局域种群的适宜下限,即要达到若干个局域种群的规模才能有效保护基本的基因库。
从竹类种质适度保护思想出发,则区域竹类生态系统内局域种群则不作保护上的数量要求,仅作集合种群数量上的要求。这是基于群落物种呈现共生状态,在竹类种质与其生态系统组分相互依存,在竹类种质居群数量增加时,其他物种种群的数量就会呈现下降趋势,在多个濒危物种同时存在时,一味增加单一种质种群数量是不理智的,且不具有可持续性。单一竹类种质具体保护策略还需要研究濒危机制,很多濒危物种是基于有性生殖衰退、生态位退化、种质退化而长期局域狭窄分布,或者依靠无性繁殖勉强维持种群,往往都是基于局域种群结构度、集合种群结构度、生态系统结构度低于物种基因库保存所需要的最小阈值所致。这种情况下,除了生态系统结构度恢复外,还需要因时因地制宜来依照濒危机制来确定合理的保护策略。
4 濒危竹种优先保护的策略
竹类种质适度保护首先需要确立合理的种群结构度,明确每一竹种的分布与种群数量特征,单纯的竹类竹种编目不是单纯的生物多样性调查,而是建立在全分布区基础上的多样性编目,建立竹类种质分布与生存档案,确定灭绝等级,依据优先保护等级来确定抢救性保护的策略与措施。总体上看,竹类种质基因库保护是基于完善的竹林生态系统结构度与功能的保护,生态系统途径是保护生物学的重要策略(Primack,2000),完全是基于物种自身对集合种群、生态系统的依赖性原理,生态系统是物种维持的基本单位。
4.1 加强基于集合种群与种群生态学的竹类种质多样编目
依照小种群与集合种群适宜的结构度,来确定竹类种质多样性的物种生存状况,划分每一竹种的存活等级。并识别和筛选出一批需要特别关注的处于濒危和渐危状态的竹类植物种类,并对那些濒危和渐危的竹类植物种类的生活史进行深入仔细的研究, 找出其致危环节的机制[5]。研究的重点应放在发掘尚存的种质资源并及时加以引种保存,从保护生物学的角度搞清其生物学特性、生态适应性及生殖发育机制[17]。掌握相关科属野生竹类种质资源的动态信息,并及时依照制定的适宜方案,参照保护体系进行相关方案的落实工作,区域生态系统物种编目与种群格局是必须掌握的关于竹类种质多样性国情信息。
4.2 加强各类森林生态系统保护区建设,完善就地保护
我国以保护竹类植物为主要对象的自然保护区有由川、陕、甘3省交界地区由卧龙自然保护区、王朗自然保护区、唐家河自然保护区、佛坪自然保护区、白水江自然保护区等20 多个自然保护区组成的大熊猫自然保护区网[5]。截止2007年,我国已经建立了自然保护区2 543个,覆盖了竹类分布的所有范围,尤其是竹类广泛分布的西南地区、华南地区、华东地区、华中地区、西北地区,以及黄河、淮河、三江流域的潜在分布区。众多的森林生态系统类型自然保护区中, 竹类植物是其中重要的组成物种,如西双版纳自然保护区的黄竹、野龙竹(Dendrocalamus semiscandens)、香糯竹(Cephlostalchyum pergracile) 和武夷山自然保护区的武夷山苦竹(Pleioblastus wuyishanensis)、长鞘茶杆竹(Pseudosasa tongivaglnata)、武夷玉山竹(Yushania wuyishanensis) 等[5]。这些保护区面积广大,几乎覆盖了所有珍稀特有竹种分布区,为竹类的就地保护创造了良好条件。大面积自然保护区的建立、管理机构的不断健全、一系列政策法规的贯彻实施,使丰富的竹类种质资源在其天然生长地得到了有效保护(辉朝茂等,2003)。
完善我国现有自然保护区建设与管理,在现有保护区基础上通过合理的林地或土地流转来调整保护区与生境廊道格局,形成利于竹类种质保护的保护区结构与网络,彻底构建我国竹类野生种质的有效就地保护体系。自然保护区通过全民所有经营权控制,能有效遏制由于土地垦殖、牧业生产、狩猎、砍伐、采集等导致的竹类种质濒危或灭绝,在人为干扰导致的物种濒危条件下保护成效显著。
4.3 加强我国4大区域竹类种质保存圃的建立和完善
对珍稀竹种种质资源的保护还可以采取就地保护和迁地保护的方法,建立大面积的自然保护区,建成种质资源引种保存和实验研究基地,不断地完善管理机构,推进一系列政策法规的实施[17]。为尽快切实有效地保护、抢救、研究和利用珍稀种质资源,辉朝茂等(2003)也曾建议以现有珍稀竹种园为基础,加大力度给予专项资金在全国建成3大种质资源引种保存和试验研究基地,东亚珍稀竹种园、东南亚珍稀竹种园与喜马拉雅珍稀竹种园。建立和完善热带、亚热带、温带与寒温带4大区域竹类种质保存圃,形成旅游、种苗繁育、珍稀资源保存、良种培育、科学研究、科普示范、观赏为一体的种质研发、生产与加工一体的实体,种类涵盖所有竹类分布区,在野生竹类种质基因库与人工保存基因库之间建立迁地保护与遗传改良的基地与平台,保护竹类种质与基因库。基地建立后可通过开发生态文化旅游来以园养园(辉朝茂等,2003),升级传统竹种园,提升规模,完善管理,增加科技含量,增加覆盖面,强化生物种质产业在新兴产业中的地位,构建竹类种质的人工保存体系,间接强化野生种质的保存安全。
4.4 加强竹类种质资源定向培育
竹资源定向培育需要建立“三种保障体系”,即技术和人才保障体系、政策和资金保障体系以及资源与环境保障体系[29]。竹类资源定向培育需要建立种质良种化、繁育壮苗化、栽培高效化、经营集约化、采收生态环保化,从种质资源培育、种苗供应与栽培经营等多层次上构建竹林资源培育技术模式,带动竹类种质保存圃、繁育圃与生产圃建设,并发挥种质保护的产学研价值。竹类多样性保护和竹产业可持续发展,必须确立以保护生态环境为基础、经济效益与生态效益并重的竹业发展方针[30]。竹林具有发达的鞭根系统和很强的固土能力,涵养水源、保持水土的功能超过其它人工林;竹林植被也是许多珍稀野生动物的食物来源和栖息场所,如珍稀动物大熊猫和金丝猴就依赖竹林生存,共同形成良好的生态关系。通过培育竹资源,发展竹产业,可以在较短的时间内辐射带动若干相关产业,同时更好地突出生态环境建设。
竹类分布地带坡度大,岩石出露,降水量大,是流水侵蚀区,土壤肥力容易遭受水土流失的危害。加强坡度大于25°以上私人经营土地竹林生态系统的保护,完善以混交林为主体的竹林结构,保育竹林可持续经营的群落优化结构,加强大坡度竹林的生态功能维护,为该地带竹类种质持续发育提供现实的生态承载力。从小流域竹林生态系统结构与功能出发,研究竹林-土壤-大气连续体、水文活动与水土保蓄、地力恢复的关键技术,从生态系统管理与可持续经营的高度建立竹林经营管理制度,并寻求技术落实的法制途径。突出竹类种质在园林观赏、生态环保、水土保持、竹材生产、森林蔬菜生产等原料供应中的重要价值,形成竹类栽培基地与原料生产基地的管理与检测,尤其是珍稀濒危竹种,与种质保存圃、繁育基地形成竹类种群人工保护与定向培育的动态数据库,为竹类种质长效保护提供数据支撑。
[1] Moul i a B, Cout and C, Lenne C,. Post ur e cont r ol and skel et al mechanical accl imat ion in t er r est r ial pl ant s: impl icat ions f or mechanical model ing of pl ant archit ect ur e.Amer ican Jour nal of Bot any, 2006, 93(10):1477-1489.
[2] 江泽慧.世界竹藤[M].沈阳:辽宁科学技术出版社,2004.
[3] 费世民,等.竹林生态研究[M].北京:中国林业出版社,2012.
[4] 单家林.海南竹类的地理分布及区系特征[J].植物研究,1997,17(4):403-408.
[5] 李睿,章笕,章珠娥.中国竹类植物生物多样性的价值及保护进展[J].竹子研究汇刊,2003,22(4):7-17.
[6] 周芳纯.20世纪竹业的回顾和21世纪的展望[J].竹子研究汇刊,1999,18(4):1-4.
[7] 粱泰然.中国竹林类型与地理分布特征[J].竹子研究汇刊,1990,9(4):1-6.
[8] 辉朝茂. 怒江竹类[M]. 北京:中国农业科技出版社,1994: 17-122.
[9] 杨宇明,辉朝茂.云南竹亚科植物地理分布区划研究[J].竹子研究汇刊,1999,18(2):20-25.
[10] 易同培.四川竹林自然分区[J].竹子研究汇刊,1997,16(3):5-22.
[11] 林万涛.广东竹林自然区划[J].竹子研究汇刊,1992,11(4):1-6.
[12] 张喜.贵州竹林区划[J].竹子研究汇刊,1994,13(2):1-l 4.
[13] 李睿.浙江省竹类植物的区系地理特点[J].竹子研究汇刊,2001,20(4):10~17
[14] Liese W. Advances in Bamboo Research[J]. 南京林业大学学报(自然科学版),2001,25(4): 1-6.
[15] 樊宝敏,李智勇.黄河流域竹类资源历史分布状况研究[J].林业科学,2005,41(3):75-81.
[16] 周世强.竹类的几种觅养生长格局[J].四川林勘设计,2002(4):28.
[17] 李华,辉朝茂.中国竹类多样性特征及其保护研究进展[J].云南大学学报(自然科学版),2008,30(S1):443-445.
[18] 钟章成,李睿.慈竹的数量生态研究[J].植物生态学与地植物学报,1990,14(1):63-68.
[19] 刘庆,钟章成.斑苦竹无性系生长与水分供应及其适应对策的研究[J].植物生态学报,1996,20(3):245-254.
[20] 刘庆,钟章成.斑苦竹无性系种群克隆生长格局动态的研究[J].应用生态学报,1996,7(3):240-244.
[21] 刘庆,钟章成,何海.斑苦竹无性系种群在自然林和人工林中的生态对策[J].重庆师范学院学报(自然科学版),1996,13(2):16-21.
[22] 杜凡,薛嘉榕,杨宇明,等.15年来云南竹子的开花现象及其类型研究[J].林业科学,2000,36(6):57-68。
[23] 王微,陶建平,宋利霞,等.竹类植物种群生态学研究进展与展望[J].广西植物,2006,26(4):412-417.
[24] 张文燕,马乃训.竹类植物花期生物学特性[J].林业科学研究,1989,2(6):596-600.
[25] 余群洲,吴萌,赵木虎,等.大熊猫主食竹开花习性的初步研究[J].四川林业科技,1994,8(1):49-54.
[26] 邢新婷,傅懋毅,肖贤坦. 麻竹开花生物学特性观察及控制授粉的初步研究[J]. 北京林业大学学报, 2005, 27(6): 103-107.
[27] 陈光才,马乃训.竹子遗传育种研究进展[J]. 林业科学研究,2005,18(6):749-754.
[28] 王峥峰,彭少麟,任海.小种群的遗传变异和近交衰退[J].植物遗传资源学报,2005,6(1):101-107.
[29] 熊文愈等.中国农业百科全书-林业卷(竹类条目)[M].北京:农业出版社,1988.
[30] 辉朝茂,胡冀珍,张国学,等.中国竹类多样性及其可持续利用研究现状和展望[J].世界林业研究,2004,17(1):50-54.
(参考文献略)
