新疆伊犁两种生境无芒雀麦种群的年龄结构
2012-08-20贾娜尔阿汗李海燕杨允菲李建东
张 维,贾娜尔·阿汗,张 娜,李海燕,杨允菲,李建东
(1.伊犁师范学院化学与生物科学学院,新疆 伊宁835000;2.东北师范大学草地科学研究所植被生态科学教育部重点实验室,吉林 长春130024)
无性系植物(克隆植物)普遍具有营养繁殖的特性[1],其通过营养繁殖来增加数量和水平扩散[2]。种群年龄结构不仅反映种群的不同年龄个体的配置情况、现存种群的状态,也反映种群的数量动态及其发展趋势。1990年开始,国内外对植物种群年龄结构的研究主要集中在木本植物[3-5],随着无性系草本植物年龄结构划分方法上的突破,国内在该领域相继开展了一系列研究工作[6-10]。
无芒雀麦(Bromus inermis)是短根茎型多年生禾草,是典型的无性系植物[11],其具有生态适应性强、耐践踏、产草量大、营养价值高、适口性好等刈牧兼用的优良特性。因此,国内外对无芒雀麦的抗逆性、遗传多样性、生物学特征、生理特性、生长规律、种间竞争、产量[12-19]等方面开展了较多研究,但对无芒雀麦种群年龄结构的研究尚未见报道。本研究从种群水平对两种生境条件下无芒雀麦自然种群分蘖株、根茎及潜在种群的年龄结构进行了分析,旨为揭示无芒雀麦种群在不同生境条件下的繁殖潜力,以期为牧草的管理和生产实践提供科学依据。
1 材料与方法
1.1 试验地概况 本试验在新疆伊犁州昭苏县进行,地理坐标为80°10′~80°30′E,42°38′~43°15′N,海拔1 323~6 995m,属温带山区半湿润半干旱、冷凉型气候。年平均气温2.7℃,1月平均气温-11.7℃,7月平均气温14.6℃,极端最高温33.5℃,最低温-34.1 ℃;≥10 ℃年积温1 328.1℃·d;无霜期85~100d;年均降水量570mm;年蒸发量1 340mm;积雪期158d,积雪厚度20~60 cm;平均日照时间2 540h[20]。生境1(H1)在夏塔乡河岸边湿地,为草甸生境,土壤为黑钙土,无芒雀麦为优势种,伴生植物主要为草原苔草(Carex liparocarpos)、草地早熟禾(Poa pratensis)、问荆(Equisetum arvense)、野苜蓿(Medicago falcata)等;生境2(H2)在昭苏镇公路两侧,土壤层薄,主要为沙砾,难保水,为干旱生境,无芒雀麦为伴生种,主要植物有 羊 茅 (Festuca ovina)、问 荆、羊 草 (Leymus chinensis)、沙生针茅(Stipaglareosa)和北艾(Artemisia vulgaris)等。
1.2 取样方法 2010年8月下旬,选取植被未受干扰的地段取样。生境1取样面积为25cm×25 cm,生境2由于无芒雀麦较稀疏,取样面积为50cm×50cm,土柱深度均为25cm,5次重复。取样时,尽量保持土柱的完整性,用样品袋分装并标记。带回室内用水轻轻冲洗干净,注意保持无芒雀麦地上分蘖株与地下根茎、活分蘖株与枯死分蘖株、芽与分蘖株和根茎的自然联系,以便于鉴别与计数。对无芒雀麦种群年龄结构的研究方法与羊草、赖草(L.secalinus)等其他无性系植物相同[6-9]。
1.3 分蘖株、根茎、冬性苗及芽年龄的划分和数据处理 根据形态特征,以根茎分蘖节营养繁殖存活的世代数来确定无芒雀麦分蘖株、根茎的年龄,按分蘖节上残留的株痕鉴别,具体划分方法见文献[21]。由根茎顶端和根茎节间向上生长的绿芽和未拔节的分蘖株一并称为冬性苗。芽的年龄划分标准为:各龄分蘖节横向生长的根茎顶端芽和根茎节间芽为0龄芽,由1龄分蘖节产生的向上生长的芽为1龄芽,由2龄分蘖节产生的向上生长的芽为2龄芽,依次类推。分别计数样方中各龄级分蘖株、冬性苗和芽的数量,测量各龄根茎的长度。将样方取样所调查测定的各种数量指标均换算成1m×1m的常规单位面积数量指标,用SPSS 13.0统计软件分析和作图。
2 结果与分析
2.1 分蘖株的年龄结构及其年龄谱 两个生境样地中,无芒雀麦种群的分蘖株均由营养分蘖株和生殖分蘖株组成,营养分蘖株有4个龄级,生殖分蘖株有3个龄级;营养分蘖株和生殖分蘖株数量均为草甸生境>干旱生境,草甸生境的分蘖株数是干旱生境的1.8倍(表1)。营养分蘖株数为1龄>2龄>3龄>4龄;生殖分蘖株数量在草甸生境为1龄>2龄>3龄,在干旱生境为1龄>2龄=3龄;分蘖株数在部分龄级间差异显著。各龄分蘖株的标准差(SD)均较大,有的接近或大于其平均数(M),其中3龄或4龄变化更大,表明无芒雀麦无性系种群分蘖植株数量在各样方内存在着较大的差异。由相同龄级营养分蘖株和生殖分蘖株数之和与全体分蘖株数的百分比构成无芒雀麦种群分蘖株的年龄谱(图1)。不同生境样地,分蘖株均以1龄比例最大,以1龄和2龄占绝对优势,在草甸生境中,1龄和2龄分蘖株占93.6%;在干旱生境中,1龄和2龄分蘖株占85.7%。从分蘖株数量的年龄谱反映出两种生境无芒雀麦种群均呈增长型年龄结构。
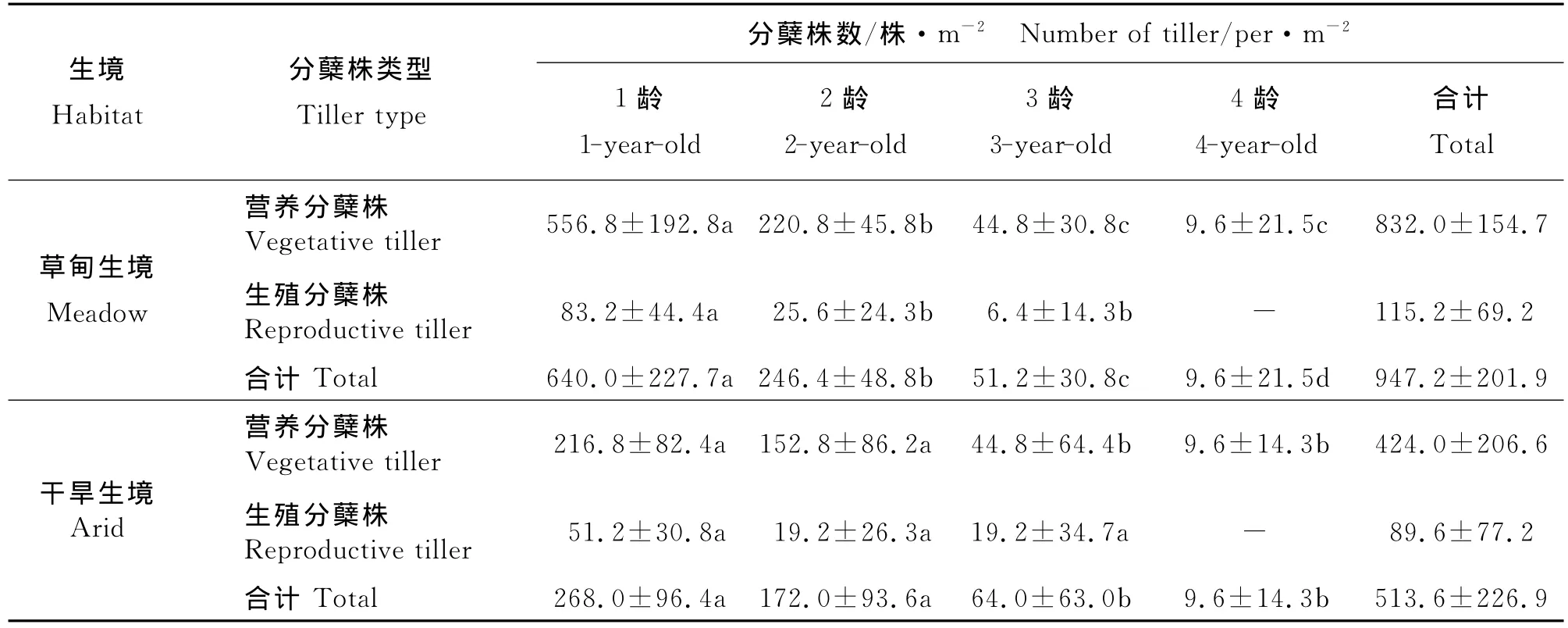
表1 两种生境无芒雀麦种群不同龄级分蘖株的数量组成Table 1 Age class distribution of tillers in Bromus inermis populations in two habitats
2.2 根茎的年龄结构及其年龄谱 在两种生境中,无芒雀麦种群根茎均为4个龄级,根茎累积长度为草甸生境>干旱生境(表2)。在草甸生境中,根茎长度为1龄>2龄>3龄>4龄,即随龄级增加,根茎长度减小,其中1龄根茎长度是4龄根茎的21.1倍;1、2龄与3、4龄差异显著(P<0.05)。在干旱生境中,根茎长度为2龄>1龄>3龄>4龄。
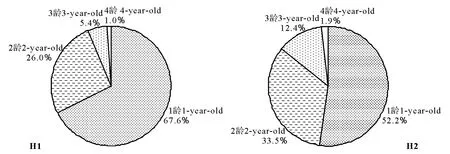
图1 两种生境无芒雀麦种群分蘖株的年龄谱Fig.1 The age spectrum of tillers in Bromus inermis populations in two habitats

表2 两种生境无芒雀麦种群不同龄级根茎长度及其多重比较Table 2 The length and multiple comparisons on rhizomes of different age classes in Bromus inermis populations in two habitats
由各龄根茎长度与全体根茎长度的百分比构成无芒雀麦种群根茎的年龄谱(图2)。在草甸生境中,1龄根茎比例最大,2龄根茎次之,1龄和2龄合计占根茎长度的88.5%,3龄根茎比例急剧减小,4龄根茎仅为2.7%;在干旱生境中,1龄和2龄根茎比例差异不大,1龄和2龄合计占根茎长度的76.9%。根茎是无芒雀麦重要的繁殖器官,根茎的长度反映了其地下部分的扩展,由此反映出无芒雀麦种群地下部分的扩展主要由1龄和2龄根茎贡献。2个生境均以低龄级的根茎占优势,从根茎年龄谱也反映出无芒雀麦种群呈增长型年龄结构。
2.3 潜在种群的年龄结构及其年龄谱 在生长季末期,无芒雀麦无性系潜在种群均由冬性苗、根茎顶芽和各龄分蘖节芽组成(表3)。芽有3个龄级,冬性苗有4个龄级;芽数量为草甸生境>干旱生境,冬性苗数量为草甸生境<干旱生境。芽数量为0龄>1龄>2龄>3龄,部分龄级间芽数量差异显著(P<0.05);冬性苗数量为1龄>2龄>3龄>4龄,但各龄冬性苗数量差异未达到显著水平(P>0.05)。两个生境样地,冬性苗和芽均以低龄级数量居多,从潜在种群的数量组成也反映出无芒雀麦种群呈增长型年龄结构。
由各龄冬性苗或芽的数量与全体冬性苗或芽的数量百分比构成无芒雀麦潜在种群的年龄谱(图3)。两种生境,冬性苗均以1龄占最大比例,2龄次之,3龄再次之,4龄最少。冬性苗以1龄和2龄占优势。两个生境样地均以0龄和1龄芽占优势。在草甸生境中,0龄和1龄芽占86.3%;在干旱生境中,0龄1龄芽占75.8%。生境不同,各龄芽的配置差异比较大,如在草甸生境中,0龄芽占57.5%,而在干旱生境中0龄芽仅占38.2%。
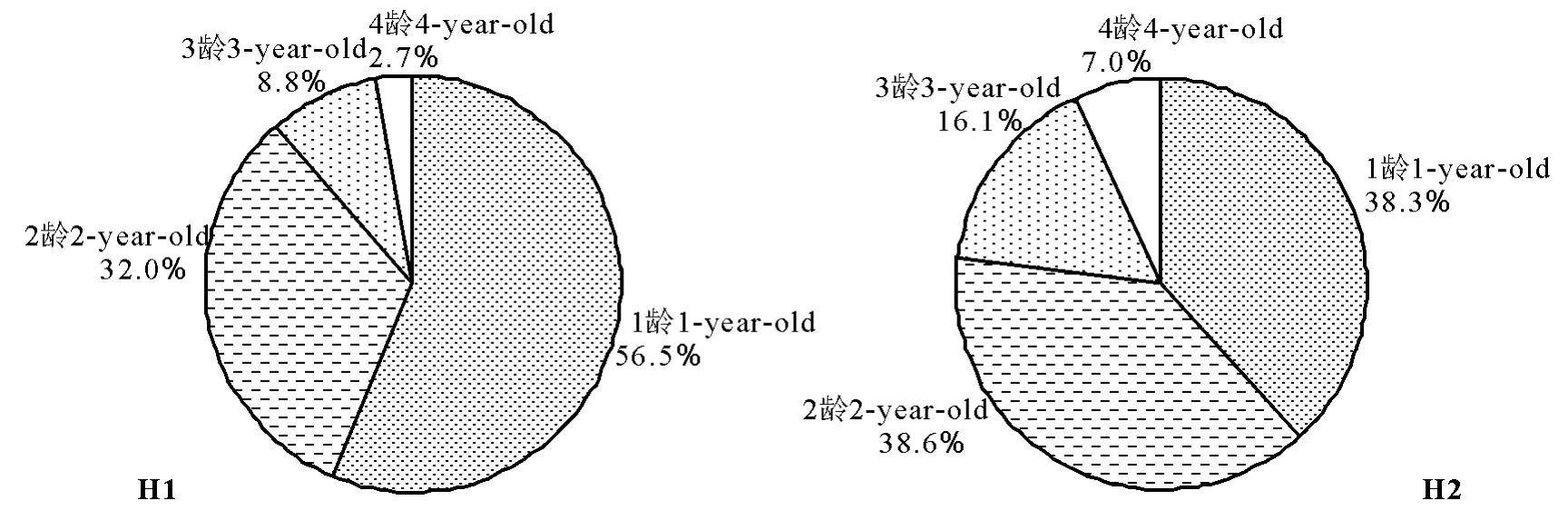
图2 两种生境无芒雀麦种群根茎长度的年龄谱Fig.2 The age spectrum on length of rhizomes in Bromus inermis populations in two habitats
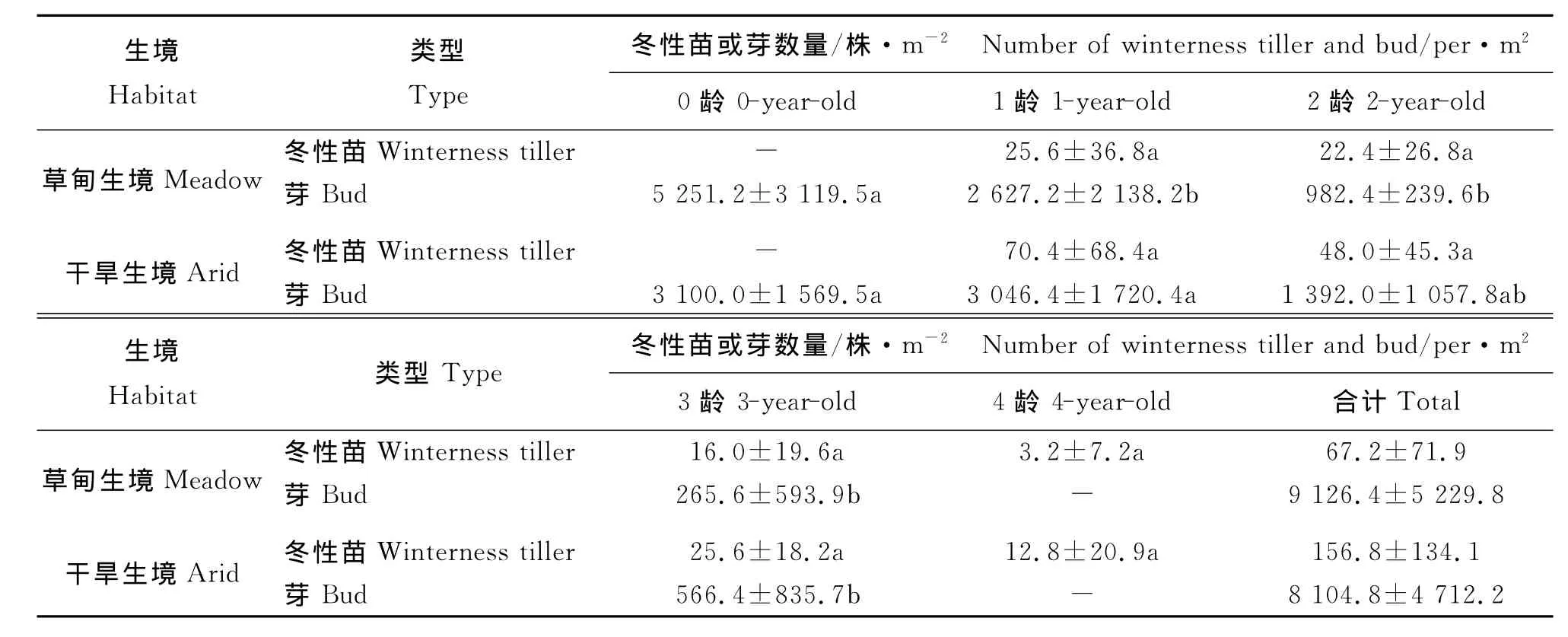
表3 两种生境无芒雀麦潜在种群不同龄级组成数量及其多重比较Table 3 The component number and multiple comparisons on different age classes of Bromus inermis potential populations in two habitats
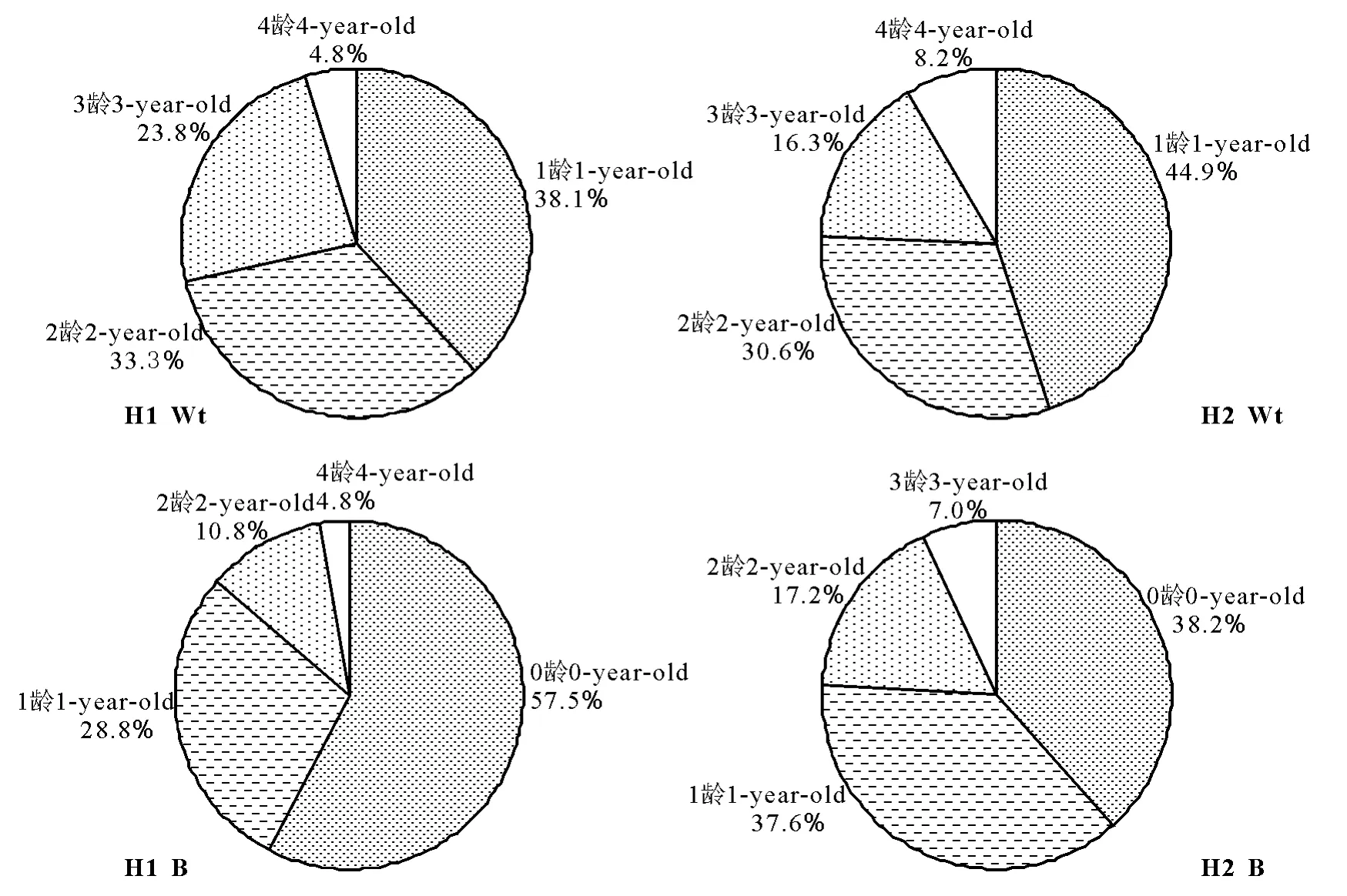
图3 两种生境无芒雀麦潜在种群中冬性苗(Wt)和芽(B)的年龄谱Fig.3 Age spectrum of winterness tillers and buds in Bromus inermis potential populations in two habitats
3 讨论与结论
两种生境,无芒雀麦无性系种群均呈增长型年龄结构;种群生殖分蘖株有3个龄级,营养分蘖株、根茎、冬性苗和芽均有4个龄级;按照杨允菲等[21]的判断方法,无芒雀麦种群的根茎分蘖节最多繁殖4个世代,最多可存活5个生长季。随龄级增加,无芒雀麦的分蘖株、冬性苗、芽的数量急剧减少,根茎累积长度也急剧减小,即低龄级构件占更大的比例,与赖草(Leymus secalimus)[6]有相似的年龄结构特征。由此预示着无芒雀麦无性系种群翌年也将呈增长型的龄级结构。
分蘖株能否进入生殖生长阶段是衡量无性系植物生活力的重要标准之一[7]。无芒雀麦无性系种群均具有营养分蘖株和生殖分蘖株,表明不同生境无芒雀麦种群均可以进入有性生殖阶段,但两个生境均表现为营养分蘖株数量远大于生殖分蘖株,则又反映出无芒雀麦无性系种群的繁殖以营养繁殖为主。分析营养分蘖株数量远大于生殖分蘖株数量的原因,第一,可能为测定时间较晚,新疆昭苏县地处高寒地区,到8月下旬,部分生殖分蘖株已经进入果后营养期,已有部分新营养分蘖株形成;第二,无芒雀麦无性系种群具有充分的生长空间,种群将更多的能量投入到营养生长和营养繁殖以占据现有空间,从而降低了有性生殖的比率。
植物种群由不同生活史阶段的个体组成,而不同生活史阶段个体的数量配置体现着种群的生存与发展,以及种群的调节功能[7]。草甸生境的营养分蘖株和生殖分蘖株数量均大于干旱生境,在草甸生境中,土壤水分充足、营养物质丰富,无芒雀麦分蘖节产生较多的直接向上生长的芽并发育成分蘖株,以补充母株的空间位置。在该生境,无芒雀麦是优势种,种内竞争激烈,无芒雀麦分蘖节产生生殖分蘖株进行有性生殖以扩大种群。在干旱生境中,土壤水分、养分相对贫乏,且无芒雀麦是伴生种,种内的竞争作用相对较小,在有充分扩展空间的条件下,分蘖节将产生较多的横向生长根茎远离母株,再以根茎顶芽发育成1龄分蘖株以实现水平扩散。可见,无芒雀麦无性系种群对生态环境变化具有一定的调节能力,群落微生境是影响无芒雀麦种群分蘖株年龄结构的重要因素。
无芒雀麦的1龄根茎由各龄分蘖节产生且均有生长点。根茎累积长度均以1龄和2龄占优势,不同生境条件下,不同龄级根茎的累积长度具有不同的结构特征,表现出较大的生态可塑性。根茎顶芽为当年形成,源自于1龄根茎并由1龄根茎不断向其供应养分,可以远离母株以有效拓展无性系种群的生态位空间。在新疆昭苏,无芒雀麦生长季末期根茎顶芽将输出为冬性苗。冬性苗形成来年的冬性植株,冬性植株的生长发育状况将直接关系到种群在翌年的有性生殖状况。在生产上,应加强对无芒雀麦生长季后期的管理,以促进翌年的种子生产。无芒雀麦在生长季的中后期,通过营养繁殖为下一年的生存和繁衍准备了8 000株·m-2以上的潜在种群,但是,潜在种群的数量并不意味着翌年的实际种群中的个体数量,无芒雀麦种群将通过种群调节来实现种群的补充和更新。
[1] Harper J L,White J.The demography of plants[J].Annual Review Ecology System,1974,5:419-463.
[2] Silvertown J,Charlesworth D.Introduction to Plant Population Biology[M].Fourth Edition.London:Blackwell Science,2001:11-13.
[3] Antos J A,Parish R,Conley K.Age structure and growth of the tree-seedling bank in subalpine spruce-fir forests of south-central British Columbia[J].American Midland Naturalist,2000,143(2):342-354.
[4] Paynter Q,Downey P O,Sheppard A W.Age structure and growth of the woody legume weed Cytisus scoparius in native and exotic habitats:Implications for control[J].Journal of Applied Ecology,2003,40(3):470-480.
[5] 谢宗强,陈伟烈,路鹏,等.濒危植物银杉的种群统计与年龄结构[J].生态学报,1999,19(4):523-528.
[6] 杨允菲,张宝田.松嫩平原赖草无性系生长及其构件的年龄结 构 [J].应 用 生 态 学 报,2004,15(11):2109-2112.
[7] 杨允菲,刘庚长,张宝田.羊草种群年龄结构及无性繁殖对策的分析[J].植物学报,1995,37(2):147-152.
[8] 赵玉,李海燕,贾娜尔,等.伊犁河谷不同生境假苇拂子茅种群构件组成及其年龄结构[J].草业学报,2009,18(2):89-94.
[9] 张维,贾娜尔·阿汗,赵玉,等.新疆伊犁2种生境大拂子茅种群年龄结构研究[J].草地学报,2011,19(6):898-903.
[10] 孟令宇,李海燕,杨允菲.退化羊草群落恢复演替系列种群构件年龄结构特征[J].草业科学,2011,28(5):807-812.
[11] 杨慧玲,薛瑞丽,叶永忠,等.克隆整合对无芒雀麦在异质性盐分环境中存活和生长的影响[J].生态学报,2009,29(6):2827-2834.
[12] 胡生荣,高永,武飞,等.盐胁迫对两种无芒雀麦种子萌发的影响[J].植物生态学报,2007,31(3):513-520.
[13] 宋旭红,于涛,田青松,等.69份无芒雀麦种质资源遗传多样性分析[J].西北农业学报,2010,19(12):87-93.
[14] 张仁平,于磊,鲁为华.混播比例和刈割期对混播草地产量及品质影响的研究[J].草业科学,2009,26(10):109-112.
[15] 张淑艳,张玉龙,王晓东,等.氮肥对无芒雀麦生理特性影响的初步研究[J].草业科学,2009,26(5):139-143.
[16] 毛培春,孟林,高洪文,等.39份无芒雀麦种质材料苗期抗旱性综合评价[J].草业科学,2010,27(11):82-88.
[17] Trygve S A.Do low,non-freezing night temperatures during anthesis affect seed set in Bromus inermis Leyss.and Poa pratensis L[J].Journal of Applied Seed Production,1995,13:1-9.
[18] 孙铁军,韩建国,赵守强,等.施肥对无芒雀麦种子产量及产量组分的影响[J].草业学报,2005,14(2):84-92.
[19] Stewart A V.Potential value of some Bromus species of the section Ceratochloa[J].New Zealand Journal of Agricultural Research,1996,39:611-618.
[20] 李军保,马存平,刘洪来,等.围封对昭苏马场春秋草场种群生态位的影响[J].中国草地学报,2009,31(2):86-91.
[21] 杨允菲,郑慧莹,李建东.根茎禾草无性系种群年龄结构的研究方法[J].东北师大学报(自然科学版),1998,30(1):49-53.
