3种金鸡菊的光合特性比较
2012-05-29郭运雪蒋文伟黄建荣
郭运雪,蒋文伟,戴 锋,黄建荣
(1.浙江农林大学 风景园林与建筑学院,浙江 临安 311300;2.上房园艺有限公司,上海 201114)
金鸡菊属Coreopsis为菊科Compositae多年生宿根草本花卉,原产美洲,是一种观赏植物。可观叶,也可观花。茎直立,多分枝,喜阳光直射,性强健,耐寒,耐干旱瘠薄。春夏之间,花大色艳,常开不绝,适合布置花坛或作花境及背景材料,丛植于山前、篱旁、林中,应用较广泛,亦可作切花。其种类丰富,管理粗放,适应性强,还能自行繁衍,是极好的疏林地被和屋顶绿化材料,应用非常普遍。因其既耐干旱瘠薄又管理粗放,花后修剪可秋季二次开花,提高了观赏质量,而且大大节约了养护成本。另外,金鸡菊在中国长江流域以南表现常绿状态,这样冬季有景可观,弥补了宿根草本冬季落叶后景观较差的缺点。由于具有观赏性及其生长的抗逆性,奥运会、世博会都有金鸡菊靓丽的身影,也被作为公路或城市中的其他地段水土保持的植被[1-3]。目前,对金鸡菊的研究主要集中在栽培技术[4-5],提取物降压降脂的作用[6-7],黄酮类化合物的提取和乙醇提取物对菌类的抑制作用[8-9],植株水浸液的化感作用[10]等方面,对金鸡菊的光合生理特性等生态适应性方面的研究少见报道。一定程度上,光合特征参数的变化能够反映植物对环境因子的适应性[11-12]。在自然状态下,结合环境因子测定该物种的光合作用更能直接反映该植物的光合生理过程[13]。在植物的主要生理生态指标中,光强响应曲线的测定可以明确植物的光饱和点和光补偿点,进而确定植物的光照类型,以便正确地在园林中应用[14]。本研究对金鸡菊的光合特性进行测定,旨在了解它们的光合生理生态特性,为进一步引种、驯化和栽培提供理论依据。
1 材料与方法
1.1 供试材料
试验材料为3种金鸡菊的扦插苗:大花金鸡菊Coreopsis grandiflora,重瓣金鸡菊Coreopsis lanceolata,天堂之门金鸡菊Coreopsis rosea,由上海上房园艺有限公司提供。将扦插苗栽植于直径为18 cm的花盆中,栽培土按园土(m)∶泥炭(m)∶珍珠岩(m) =3∶1∶1 的比例配比,常规管理。
1.2 试验方法
1.2.1 光合作用的光响应曲线的测定 选择晴天9∶00-11∶00,用Li-6400光合测定仪通过控制光照强度测定光响应曲线。样本室内气流速率控制在500 μmol·s-1,温度控制在当时的外界环境温度。在控制条件下,利用Li-6400型光合作用系统和红蓝光源进行设置,设定光照强度在0~2000 μmol·m-2·s-1范围内(由高到低),梯度为 2000,1500,1000,600,300,200,100,80,50,30,20,0 μmol·m-2·s-1,测定相应的净光合速率(Pn)值,每一个光强下有一定的适应时间,待光合测定仪上的数据稳定后再记录,每组梯度重复3次。净光合速率为0时的光强为光补偿点(PLC),净光合速率最大时的光强为光饱和点(PLS)。采用光合助手软件Photosyn Assistant 1.1.2得出光补偿点、光饱和点、最大净光合速率(Pnmax)以及表观量子效率(EAQ)。
1.2.2 净光合速率日变化的测定 采用美国产的Li-6400便携式光合作用测定系统,叶室选用6400-15叶室,选择晴朗无风的天气,在7∶00-19∶00隔2 h测定1次,3个重复,重复记录5个观测值,取其平均值作为该时刻的测定值。选择生长健康的植株,随机选取该植物向阳面的叶片进行测定。待系统稳定后,取叶片瞬时净光合速率值,同时读取蒸腾速率(Tr),气孔导度(Cond),胞间二氧化碳摩尔分数(Ci),光合有效辐射(PAR),气温(Ta),相对湿度(Rh)等生理生态指标。水分利用效率的计算公式为:水分利用效率(EWUE)=Pn/Tr。
1.3 数据处理
使用Microsoft Excel和SPSS 17.0对测得的数据进行处理和分析。
2 结果与分析
2.1 3种金鸡菊的光响应差异
统计结果表明:3种金鸡菊的光响应曲线变化有显著性的差异。从图1可以看出:在0~100 μmol·m-2·s-1的光强范围内,3种金鸡菊的净光合速率几乎呈线性增长,在 100~200 μmol·m-2·s-1,随着光照强度的增加,净光合速率的增长速度逐渐减慢,当光强达到1500 μmol·m-2·s-1以上时,金鸡菊的净光合速率的增长处于极缓慢状态,基本维持稳定。说明植物在一定的光照强度下,叶片的光合作用处于饱和状态,当光照强度达到光饱和点以后,净光合速率甚至有下降趋势。
比较3种金鸡菊之间的光响应曲线,发现3种金鸡菊叶片的净光合速率对光合有效辐射的响应趋势基本一致,均是在低光照下快速上升后逐渐稳定。在0~100 μmol·m-2·s-1时,3种金鸡菊净光合速率相差不大。在光合有效辐射增大到100 μmol·m-2·s-1后,3种金鸡菊间的净光合速率值差异明显变大,重瓣金鸡菊最大,大花金鸡菊次之,天堂之门金鸡菊最低。总体上,金鸡菊的光响应与美国紫菀Aster novae-angliae[15]等草本植物先迅速上升后趋于稳定的光响应规律基本一致。表1的数据说明不同种金鸡菊的光补偿点和光饱和点具有一定的差异。从表1中可以看出:大花金鸡菊的光补偿点和光饱和点最低,能有效地利用弱光进行光合作用;3种金鸡菊中,重瓣金鸡菊具有相对较低的光补偿点,又具有最高的光饱和点和最大的净光合速率,说明它对光的适用范围相对最大,对强光的适应能力最强,光合适应能力也最强;天堂之门金鸡菊的光补偿点最高,而光饱和点相对较低,说明其光适应范围较窄,利用强光的能力低于重瓣金鸡菊。

表1 3种金鸡菊的光合参数Table1 Photosynthetic parameters of three species of Coreopsis
2.2 3种金鸡菊的净光合速率的日变化比较

图1 3种金鸡菊的光响应曲线Figure1 Light-response curve of photosynthesis of three species of Coreopsis

图2 3种金鸡菊净光合速率的日变化Figure2 Diurnal variation of Pnof three species of Coreopsis
植物光合作用随着环境条件的变化而有所不同,一般条件下栽培植物的光合作用日变化呈单峰型或双峰型曲线[16]。由图2可知:3种金鸡菊的净光合速率日变化表现出相似的规律,均为双峰型曲线。在上午9∶00,大花金鸡菊和天堂之门金鸡菊出现第1个峰值,分别为大花金鸡菊 (7.53±0.05)μmol·m-2·s-1,天堂之门金鸡菊为(6.91 ± 0.10)μmol·m-2·s-1,重瓣金鸡菊稍晚,在 11∶00达到第 1个高峰值,为(10.46 ±0.18)μmol·m-2·s-1;之后随着光合有效辐射和气温继续升高,Rh不断降低,净光合速率开始下降,13∶00时跌入低谷,出现光合“午休”现象。
午后随着光合有效辐射的降低,净光合速率又逐渐升高,到15∶00出现另一个高峰值,由大至小依次为重瓣金鸡菊[(7.86 ± 0.10)μmol·m-2·s-1]>大花金鸡菊[(4.77 ± 0.06)μmol·m-2·s-1]>天堂之门金鸡菊[(4.23±0.13) μmol·m-2·s-1],之后都迅速下降,至 19∶00 降至最低。方差分析表明,3 种金鸡菊间的第 1峰值和第2峰值差异均显著(P<0.05),每种金鸡菊的第1峰值与其第2峰值差异也显著(P<0.05)。在光合作用日进程中,3种金鸡菊的第1峰值均高于其第2峰值。
2.3 环境因子日变化特征
植物的光合速率、蒸腾速率和水分利用效率等生理过程受多种环境因子的影响[17]。图3结果显示,光合有效辐射瞬时变化较大,对植物的光合作用有显著的影响,为先升后降的单峰曲线,13∶00时达到最大值。气温呈现先升后降的变化趋势,与光合有效辐射变化相似,而相对湿度则呈现先降后升的变化趋势,与光合有效辐射的变化相反。
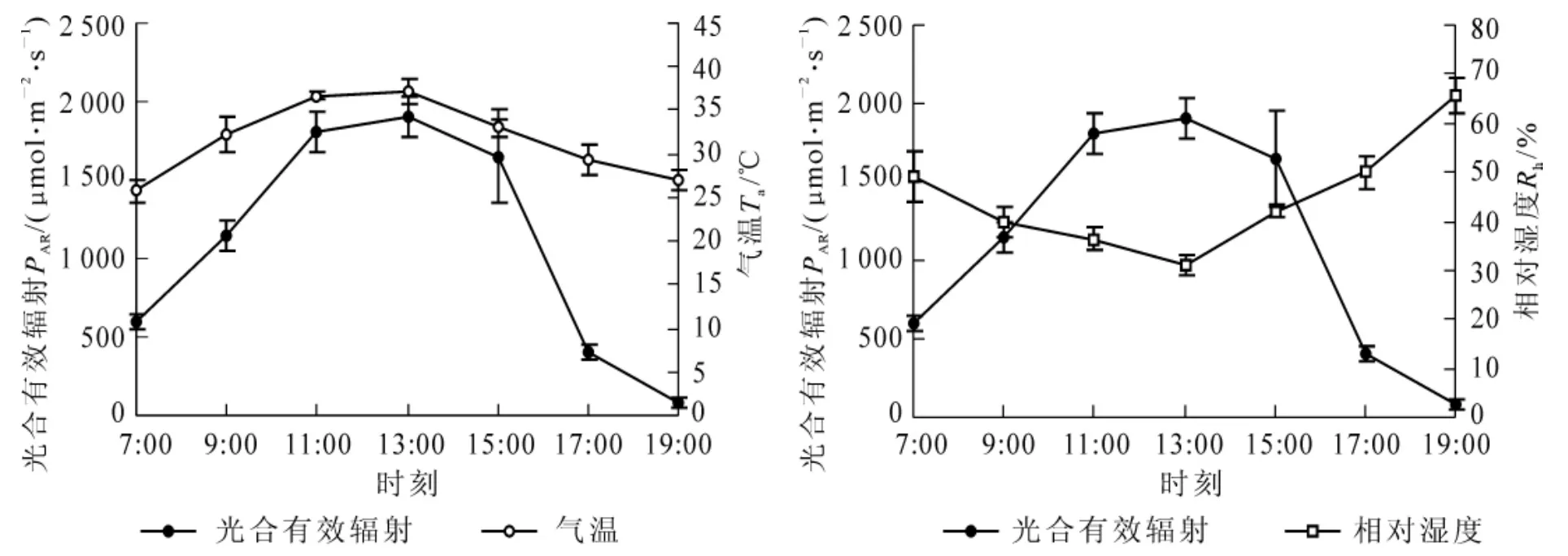
图3 光合有效辐射、气温和相对湿度日变化Figure3 Diurnal changes of photosynthetic active radiation,air temperature and relative humidity
2.4 3种金鸡菊的光合参数日变化
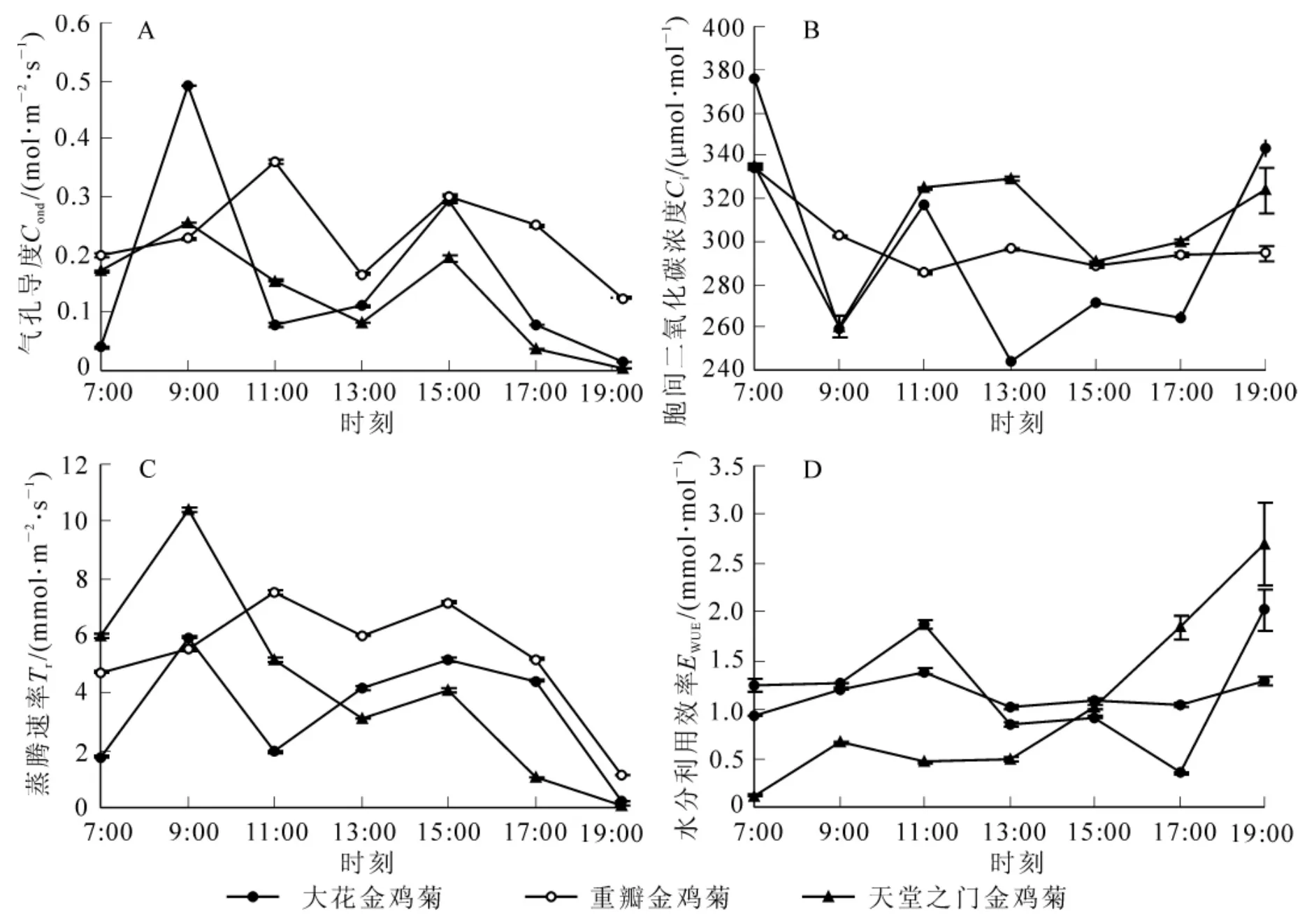
图4 3种金鸡菊的光合特征参数日变化曲线Figure4 Diurnal changes of photosynthetic character parameters of three species of Coreopsis
影响植物光合作用的生理生态因子主要包括:光合有效辐射、叶片气孔导度、胞间二氧化碳摩尔分数、蒸腾速率等。这些因子共同作用影响植物的光合速率变化。3种金鸡菊的气孔导度日变化均呈双峰型(图4),与净光合速率日变化趋势相似,大花金鸡菊和天堂之门金鸡菊的第1次峰值均出现在9∶00,峰值分别为 0.49 μmol·m-2·s-1和 0.26 μmol·m-2·s-1,重瓣金鸡菊的第 1 次峰值出现在 11∶00,为 0.36 mol·m-2·s-1,3 种金鸡菊的第 2 次峰值均出现在 15∶00,由大至小依次为重瓣金鸡菊(0.30 μmol·m-2·s-1)>大花金鸡菊 (0.29 μmol·m-2·s-1)>天堂之门金鸡菊(0.19 μmol·m-2·s-1)。气孔导度在 9∶00 以后迅速下降至13∶00达到最低,而这期间正是光合有效辐射迅速增强,气温升高,大气相对湿度降低的时候,植物能针对外界环境条件变化与植株自身状况,通过调节气孔的开闭程度,在保持植株正常生理活动的前提下,达到最大二氧化碳固定量与最小水分散失量。这也说明金鸡菊对气孔的调节能力是其能够耐干旱的生理适应机制之一[18]。3种金鸡菊的胞间二氧化碳摩尔分数与气孔导度呈相反的变化趋势,基本随着气孔导度的升高而逐渐降低,在气孔导度出现第1峰值的时刻,3种金鸡菊的胞间二氧化碳摩尔分数分别出现低谷,谷值大小分别为重瓣金鸡菊[(285.3±0.48)μmol·mol-1]>天堂之门金鸡菊[(260.57±5.09)μmol·mol-1]>大花金鸡菊[(259.38 ± 4.27) μmol·mol-1],说明随着上午金鸡菊叶片光合作用的增强,消耗更多的二氧化碳原料,使得胞间二氧化碳摩尔分数降低。3种金鸡菊的蒸腾速率也均呈现双峰曲线,出现蒸腾 “午休”现象,2次出现峰值时刻与其净光合速率达到峰值时刻相一致,但峰值大小有所变化,说明3种金鸡菊的水分利用效率有差异。由图4D可知:3种金鸡菊的水分利用效率在上午都有1个峰值,在傍晚时都呈现上升的趋势。大花金鸡菊在11∶00达到最大,为(1.87±0.05)mmol·mol-1,17∶00达到最低,随后急剧上升;重瓣金鸡菊水分利用效率日变化不大,在11∶00达到峰值 (1.39±0.03)mmol·mol-1之后有稍微下降,然后呈现缓慢上升趋势;天堂之门金鸡菊在9∶00时即达到1个峰值为(0.66±0.01)mmol·mol-1,至13∶00较平稳,之后呈上升趋势。3种金鸡菊的日平均水分利用效率大小依次为大花金鸡菊[(1.22 ± 0.015) mmol·mol-1]>重瓣金鸡菊[(1.14 ± 0.008) mmol·mol-1]>天堂之门金鸡菊[(1.03 ±0.004)mmol·mol-1]。方差分析表明:大花金鸡菊和天堂之门金鸡菊之间的日平均水分利用效率差异显著(P<0.05),与重瓣金鸡菊差异显著(P<0.05),重瓣金鸡菊与天堂之门金鸡菊之间差异也显著(P<0.05)。对于傍晚水分利用效率上升的现象,李小俊等[17]认为在午间,植物的大部分水分被用于蒸腾作用,从而相对减少了用于光合作用的水分,傍晚由于根部及时补水应用于植物叶片的光合作用,所以水分利用效率有所提高,据此可以更好地进行金鸡菊的水分管理。
3 讨论与结论
光合能力强弱在一定程度上取决于物种的遗传特性[19],光饱和点和光补偿点的变化具有一定的物种遗传稳定性[20]。3种金鸡菊的光补偿点大小排序依次为天堂之门金鸡菊>重瓣金鸡菊>大花金鸡菊;光饱和点大小顺序为重瓣金鸡菊>天堂之门金鸡菊>大花金鸡菊;最大净光合速率排序为重瓣金鸡菊>大花金鸡菊>天堂之门金鸡菊。一般情况下,光补偿点和光饱和点均较低属于耐阴植物,能充分利用弱光进行光合作用,反之属于阳性植物。光补偿点较高、光饱和点较低的植物对光照的适应性较弱,而光补偿点较低、光饱和点较高的植物对光环境的适应性较强[21]。由此得出:重瓣金鸡菊光适应范围最广,天堂之门金鸡菊光适应范围较窄。综合比较3种金鸡菊对光的适应范围大小为重瓣金鸡菊>大花金鸡菊>天堂之门金鸡菊,对强光的利用能力依次为重瓣金鸡菊>天堂之门金鸡菊>大花金鸡菊。
通过分析3种金鸡菊的光合日进程,结果发现:净光合速率均表现为双峰型,最大净光合速率均出现在上午和下午,说明3种金鸡菊在夏季出现明显的光合“午休”现象。一般认为:午间强光引起净光合速率下降,产生光抑制[18],导致植物的光合“午休”现象。Berry等[22]研究认为,导致产生光合“午休”现象的主要原因可分为气孔因素和非气孔因素,如果气孔导度减小而光合作用增强,胞间二氧化碳摩尔分数降低时,可表明气孔因素是使植物净光合速率减少,产生光合“午休”的主要原因;如果相反,当光合速率降低伴随着胞间二氧化碳摩尔分数升高时,则表明影响植物光合“午休”的原因主要是非气孔因素[23]。研究结果表明:大花金鸡菊、重瓣金鸡菊、天堂之门金鸡菊的光合“午休”主要是受非气孔限制因素即细胞光合性能下降的影响,跟Farquhar等[24]的研究结果相似。
净光合速率的变化是由植物生理生态因子共同作用的结果,而生理生态因子则受到环境因子的影响。在光合有效辐射、相对湿度、气温等环境因子相同的条件下,3种金鸡菊的净光合速率与叶片气孔导度、胞间二氧化碳摩尔分数、蒸腾速率等生理因子各不相同,说明植物的生长受多个环境因子的综合影响,各环境因子间相互制约,不同时期起主导作用的环境因子对净光合速率的影响有所差异,同一环境因子对不同植物的净光合速率、胞间二氧化碳摩尔分数和叶片气孔导度的影响也不同[18]。
水分利用效率反映了植物对水分的利用效率,也在一定程度上也反映了植物的耗水性和抗旱性[17]。仅根据水分利用效率就可以得出3种金鸡菊抗旱能力大小依次为大花金鸡菊>重瓣金鸡菊>天堂之门金鸡菊。由于水资源的日益紧缺与城市环境对于园林绿化发展用水需求之间的矛盾,在全国大部分城市,尤其是北方城市尤为突出,已经成为长时期以来城市园林绿化发展的“瓶颈”之一。如何调整城市园林绿化的品种结构入手,大力推广节水型园林植物来综合解决城市水资源紧缺与绿化发展用水需求这一矛盾,已成为摆在全国各地园林绿化建设工作者面前的紧要课题。大花金鸡菊和重瓣金鸡菊就是在中国北方城市推广节水型园林植物的一个很好的选择。
[1]秦贺兰.北京奥运用花品种系列介绍之十一大花金鸡菊生产技术[J].中国花卉园艺,2008(8):28.QIN Helan.Beijing Olympic Games with the introduction of flower varieties series(Ⅺ)production technology of Coreopsis grandiflora [J].China Flowers&Hortic,2008 (8):28.
[2]孙磊,黄建荣.新优地被“靓”世博[J].园林,2009 (8):62.SUN Lei,HUANG Jianrong.New excellent ground cover plants on the Ecpo [J].Garden,2009 (8):62.
[3]梁玉.外来种大花金鸡菊入侵的影响因子及其遗传多样性研究[D].济南:山东大学,2007.LIANG YU.The Study on Factors Influencing Invasions and Genetic Diversity in Alien Species Coreopsis grandiflora Hogg [D].Ji’nan:Shandong University,2007.
[4]任真,赵洪波.大花金鸡菊栽培[J].新农业,2008 (5):52 - 53.REN Zhen,ZHAO Hongbo.Cultivation of Coreopsis grandiflora [J].New Agric,2008 (5):52 - 53.
[5]常爽,闫瑞凤,史根花.金鸡菊花草栽培管理技术[J].北京农业,2008(36):42-43.CHANG Shuang,YAN Ruifeng,SHI Genhua.Cultivation and management techniques of Coreopsis basalis [J].Beijing Agric,2008 (36):42 - 43.
[6]梁淑红,哈木拉提,庞市宾,等.金鸡菊提取物降血压化学成分实验研究[J].时珍国医国药,2010,21(7):1619-1621.LIANG Shuhong,Hamulati,PANG Shibin,et al.Experimental study on hypertensive effects of extracts of Coreopsis basalis [J].Lishizhen Med Mat Med Res,2010,21 (7):1619 - 1621.
[7]梁淑红,庞市宾,刘晓燕,等.金鸡菊提取物降血脂作用的研究[J].中国实验方剂学杂志,2010,16(8):234-235.LIANG Shuhong,PANG Shibin,LIU Xiaoyan,et al.Study on lipid-lowering effects of extracts of Coreopsis basalis[J].Chin J Exp Trad Med Form,2010,16 (8):234 - 235.
[8]朱笃,陈飞彪,夏剑辉,等.金鸡菊总黄酮的提取及含量测定[J].食品科学,2005,26(9):314-316.ZHU Du,CHEN Feibiao,XIA Jianhui,et al.Extraction and determination of flavonoids in flowers of Coreopsis basalis [J].Food Sci,2005,26 (9):314 - 316.
[9]宗磊,李秀岚,程永芳,等.大花金鸡菊乙醇提取物对植物病原真菌的抑制活性测定[J].安徽农业大学学报,2010,37 (2):200 - 203.ZONG Lei,LI Xiulan,CHENG Yongfang,et al.Antifungal activity of ethanol extract from Coreopsis grandiflora[J].J Anhui Agric Univ,2010,37 (2):200 - 203.
[10]杜明利,高岩,张汝民,等.大花金鸡菊水浸液对6种常见园林植物种子萌发的化感作用[J].浙江农林大学学报,2011,28 (1):109- 114.DU Mingli,GAO Yan,ZHANG Rumin,et al.Allelopathic effects from aqueous extracts of exotic Coreopsis grandiflora on seed germination of six native ornamental plant species [J].J Zhejiang A & F Univ,2011,28 (1):109 - 114.
[11]尤鑫,龚吉蕊,葛之葳,等.2种杂交杨叶绿素荧光特性及光能利用[J].植物生态学报,2009,33(6):1148-1155.YOU Xin,GONG Jirui,GE Zhiwei,et al.Light energy utilization and chlorophyll fluorescence in two crossbreed poplars [J].Chin J Plant Ecol,2009,33 (6):1148 - 1155.
[12]王凯,朱教君,于立忠,等.遮阴对黄波罗幼苗的光合特性及光能利用效率的影响[J].植物生态学报,2009,33(5):1003 - 1012.WANG Kai,ZHU Jiaojun,YU Lizhong,et al.Effects of shading on the photosynthetic characteristics and light use efficiency of Phellodendron amurense seed [J].Chin J Plant Ecol,2009,33 (5):1003 - 1012.
[13]林夏珍,卢婷.遮光对窄头橐吾形态及光合特性的影响[J].浙江林学院学报,2008,25(5):614-618.LIN Xiazhen,LU Ting.Morphological and photosynthetic characteristics of Ligularia stenocephala with shading [J].J Zhejiang For Coll,2008,25 (5):614 - 618.
[14]张真珍.红花檵木的光合生理特性及其在园林中的应用研究[J].青海农林科技,2011(2):17-19.ZHANG Zhenzhen.Photosynthetic character of Loropetalum chinense and its application in the landscape and architecture [J].Sci Technol Qinghai Agric For,2011 (2):17 - 19.
[15]蒋文伟,陈娅琼,黄建荣,等.4个美国紫菀品种光合生理特征比较[J].浙江林学院学报,2010,27(6):865-871.JIANG Wenwei,CHEN Yaqiong,HUANG Jianrong,et al.Photosynthetic characteristics with four cultivars of Aster novae-angliae [J].J Zhejiang For Coll,2010,27 (6):865 - 871.
[16]吴泽民,黄成林,韦朝领.黄山松群落林隙光能效应与黄山松的更新[J].应用生态学报,2000,11(1):13-18.WU Zemin,HUANG Chenglin,WEI Chaoling.Light effect of gaps in Huangshan pine community and regeneration of Huangshan pine [J].Chin J Appl Ecol,2000,11 (1):13 - 18.
[17]李小俊,张明如,张利阳,等.太行山低山丘陵区5种木本植物光合特性的比较[J].浙江农林大学学报,2011,28(2):180 - 186.LI Xiaojun,ZHANG Mingru,ZHANG Liyang,et al.Photosynthesis,light-use efficiency and water-use efficiency for woody species in the hilly area of the Taihang Mountains [J].J Zhejiang A & F Univ,2011,28 (2):180 - 186.
[18]林保花,刘金祥,肖生鸿,等.粤西乡土香根草光合生理生态特征日动态分析[J].应用生态学报,2006,17(11):2041 - 2045.LIN Baohua,LIU Jinxiang,XIAO Shenghong,et al.Diurnal change of photosynthetic characteristics of native Vetiveria zizanioides in Western Guangdong [J].Chin J Appl Ecol,2006,17 (11):2041 - 2045.
[19]何文兴,易津,李洪梅.根茎禾草乳熟期净光合速率日变化的比较研究[J].应用生态学报,2004,15(2):205-209.HE Wenxing,YI Jin,LI Hongmei.Comparative study on daily change of photosynthesis rate of the rhi zomatous grasses in milky ripe stage [J].Chine J Appl Ecol,2004,15 (2):205 - 209.
[20]BJORKMAN O,HOLMGREN P.Adaptability of the photosynthetic apparatus to light intensity in ecotypes from exposed and shaded habitat[J].Physiol Plant,1963,16 (4):889 - 914.
[21]金则新,柯世省.云锦杜鹃叶片光合作用日变化特征[J].植物研究,2004,24(4):447-452.JIN Zexin,KE Shisheng.The diurnal variation of photosynthesis in leaves of Rhododendron fortunei[J].Bull Bot Res,2004,24 (4):447 - 452.
[22]BERRY J A,DOWNTON W J S.Environmental Regulation of Photosynthesis [M].New York:Academic Press,1982:263-345.
[23]许大全.气孔的不均匀关闭与光合作用的非气孔限制[J].植物生理学通讯,1995,31(4):246-252.XU Daquan.Non-uniform stomatal closure and non-stomatal limitation of photosynthesis [J].Plant Physiol Commun,1995,31 (4):246 - 252.
[24]FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis [J].Ann Rev Plant Physiol,1982,33:317- 345.
