青藏高原东部4科植物种子大小和比叶面积随海拔和生境的变异
2012-04-12齐威郭淑青崔现亮阳敏张永东杜国祯卜海燕
齐威,郭淑青,崔现亮,阳敏,张永东,杜国祯,卜海燕
(1.兰州大学生命科学学院生态研究所,甘肃 兰州730000;2.甘肃民族师范学院化学与生命科学系,甘肃 合作747000;3.思茅师范高等专科学校生命科学系,云南 普洱665000;4.武威市凉州区畜牧兽医局,甘肃 武威733000)
*种子大小是植物重要的生活史特征之一,它影响一个物种繁殖对策的多个方面,如萌发特征、幼苗存活率和种子数量[1]。另外,种子的散布、幼苗定居、被捕食的几率,以及群落中物种的丰富度、种群的分布格局和群落的更新演替也都与种子大小有关。种子大小的变异是植物在自然环境下选择和遗传上的一种进化行为,不同大小的种子是植物适应环境和进化的结果。种子大小变异既受遗传的控制,又受性状间协同进化的约束,还受环境的影响[2-4]。作为种子大小变异的一个主要选择压力,环境因素对种子大小的影响主要体现在温度、水分、光照以及土壤组成等形成的综合因子差异上。在区域尺度上,尤其是高山地区,海拔差异因构成了不同的水、光、热、土壤梯度,已成为影响种子大小的主要因子之一。目前,不同海拔梯度下种子大小变异已经成为生态学的研究热点之一,Pluess等[5]在阿尔卑斯山的研究后发现随着海拔升高属内种子重量随着海拔升高而增大,并认为高海拔地区严酷的环境压力不利于幼苗成功建植,大种子物种因为能产生较大的幼苗而被选择下来。但一些其他研究并不同意这一观点[6-8],他们认为净初级生产力(NPP)是解释种子大小在海拔方向上变异的变量之一,因为随着海拔升高NPP通常降低,所以单位面积内可以转化成种子的生物量和种子大小也会相应减少。
比叶面积(specific leaf area,SLA)是指单位干重的鲜叶表面积,其数值大小直接受叶片厚度、形状和重量的影响,在一定程度上反应了叶片截获光的能力和在强光下的自我保护能力[9]。SLA往往与植物的生长和生存对策有紧密的联系,能反映植物对不同生境的适应特征,也是评价群落生产力水平及其结构是否合理的指针之一。比叶面积不仅可以用来推断光合作用、蒸发、蒸散等过程以及估算陆地生态系统净生产力,而且还可以作为参数输入到水平衡、全球碳循环等模型中,SLA的测量对提高这些模型的运行效果至关重要[10,11]。另外,比叶面积是物种层面的特征参数,是物种在长期适应环境过程中形成的属性之一。正因为如此,有人提出把比叶面积作为植物适应环境策略的分类依据之一[12]。因此,加强对这一叶性状的格局分析研究将有助于从机理上解释植被对全球气候变化尤其对青藏高原高寒环境的适应与响应机制。
作为全球海拔最高的一个独特地域单元,青藏高原是敏感响应全球变化和人为干扰的关键性研究地区,也是全球生物多样性保护的重点区域之一[13-15],其东部山地是青藏高原、黄土高原和秦岭山地的交汇地带,该区海拔变化大,地形复杂,气候寒冷多变,植被垂直差异显著,群落结构复杂,物种丰富度高。毛茛科、虎耳草科、龙胆科和石竹科植物在该区域分布广泛,属青藏高原优势种。本研究致力于了解种子大小和比叶面积在高寒环境中随海拔梯度和生境变化的变异模式及其生物学和进化意义。
1 材料与方法
1.1 研究区概况
研究区位于青藏高原东部甘肃省甘南藏族自治州(100°50′~104°30′E,33°05′~35°25′N),海拔1 500~4 000 m。气候特点是高寒湿润,降水主要受西南季风控制,自东南向西北方向逐渐减少;降水季节分配极不均匀,雨季和旱季分明,年均降水量500~800 mm,雨季集中在7—9月;地面气温区域差异显著,温度季节差异明显,沿海拔气候从年均温近14℃且无霜期超7个月的暖温带南缘过渡到年均温不足-4℃且没有无霜期的高寒地带。该地区太阳辐射量达0.586~0.795 MJ/(cm2·a),是我国太阳总辐射值最高的地区之一。其植被以多年生草本和灌木为主。植被类型主要包括典型高寒草甸、高寒灌丛、亚高山针叶林、高寒沼泽草甸和高寒山地草甸等。
1.2 试验材料和方法
63种植物的成熟种子和叶片于2006年和2007年的6-10月在青藏高原东部地区的甘南藏族自治州境内采集,同时记录每种植物所处的海拔和生境特点(表1)。每一物种的种子随机在20株以上的个体上尽量多而且均匀采取,然后将每一物种的所有种子充分混合,以避免母株对种子大小的影响。去除不成熟的种子和被昆虫啃食的种子,并人工去除果肉、自然风干(种子含水量保持在12%~15%)、贮藏。以每100粒种子为单位用万分之一电子天平进行称量,3次重复,其平均值为种子平均百粒重量。每一物种的成熟、健康、完整的叶片随机在10株以上的个体上尽量多而且均匀采取,然后将每一物种的所有叶片充分混合,以避免母株对叶面积及比叶面积的影响。用扫描仪(Epson-1670)扫描植物叶片以测量叶面积。根据叶片大小,每次扫描叶子3~15片,共扫描3组。扫描完毕后用水冲洗干净,80℃下烘干至恒重,称重。比叶面积(specific leaf area,SLA)计算方法如下[16,17]:
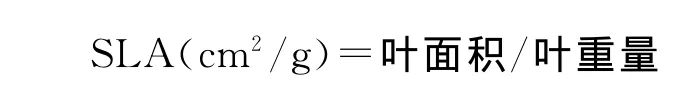
1.3 数据分析
数据分析前,种子大小和比叶面积均经对数转化以消除原始数据的不正态性,经转换后的数据用SPSS 16.0软件进行分析。其中,Person’s相关分析用于检测种子大小和比叶面积的关系,线性回归分析用于检测种子大小和比叶面积与海拔的关系,单因素方差分析(One-way ANOVA)和最小显著差数法(LSD)用于不同生境下植物种子大小和比叶面积的比较及其差异显著性检验;另外,由于海拔和生境2个因子对种子大小和比叶面积的影响可能存在共线性问题,用单因素协方差分析(One-way ANCOVA)来比较和区分二者的影响。最后,考虑到科内不同生境下的种子大小和比叶面积方差不齐,用非参数的Kruskal-Wallis H分析来检验生境对科内种子大小和比叶面积的影响,并用线性回归分析来检验科内种子大小和比叶面积随海拔的变异趋势。
2 结果与分析
2.1 种子大小与比叶面积的关系
相关分析显示63种植物的种子大小与比叶面积(表1)存在边际明显的负相关关系(R=-0.233,P=0.066)。
2.2 种子大小随海拔及生境的变异
回归分析显示不但总体上63种植物的种子大小与海拔不相关(P=0.376;图1A),而且每个科科内的种子大小与海拔也没有明显的相关性(表2)。
另外,单因素方差分析(One-way ANOVA)和最小显著差数法(LSD)未发现不同生境下植物种子大小有显著差异(图2A)。
同时,Kruskal-Wallis H检验显示4个科植物在不同生境下的种子大小均没有明显的变异趋势(表2),即生境类型对科内种子大小没有显著影响。
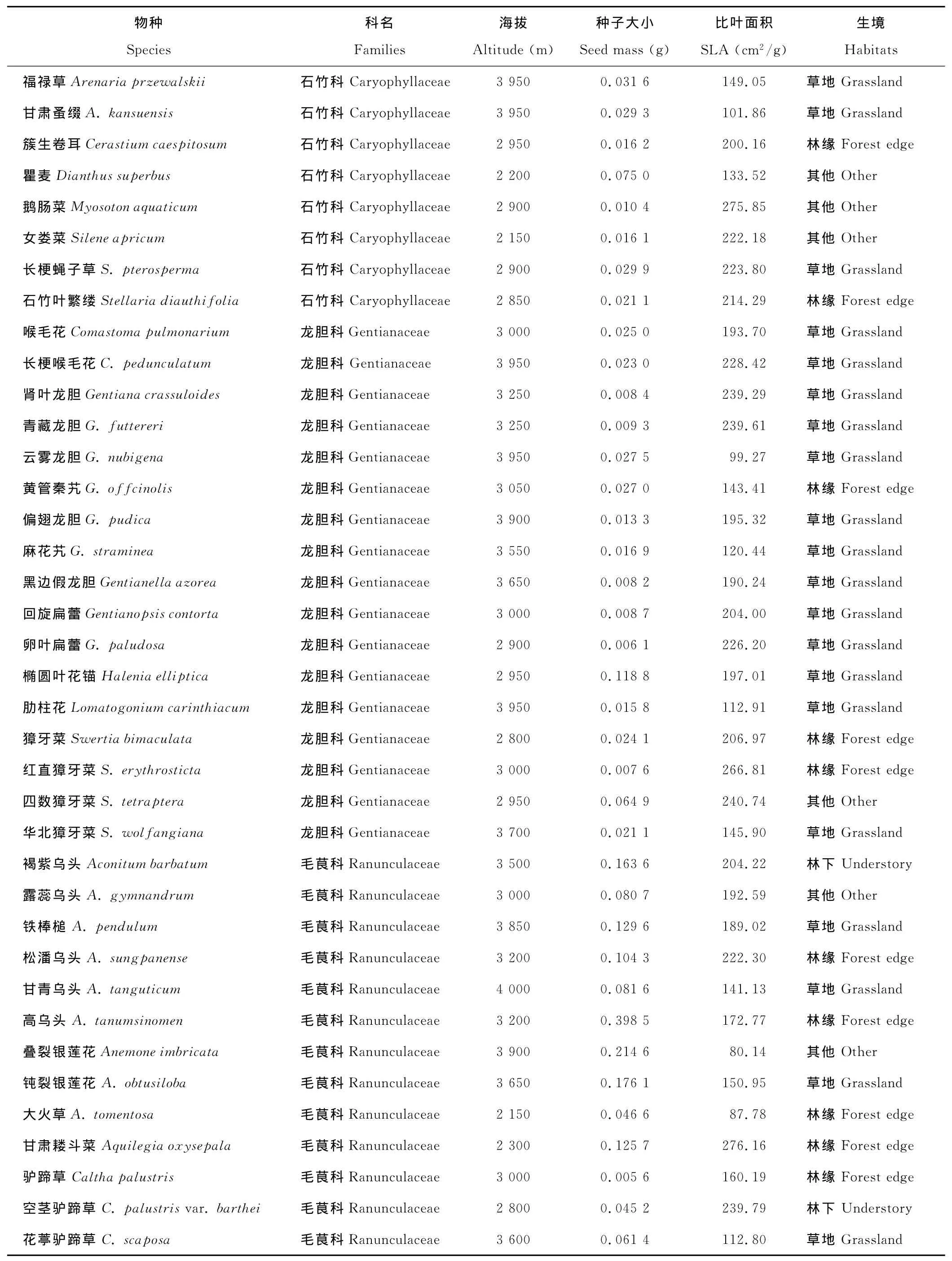
表1 试验用物种及部分数据Table 1 Species included in the experiment and part of the data
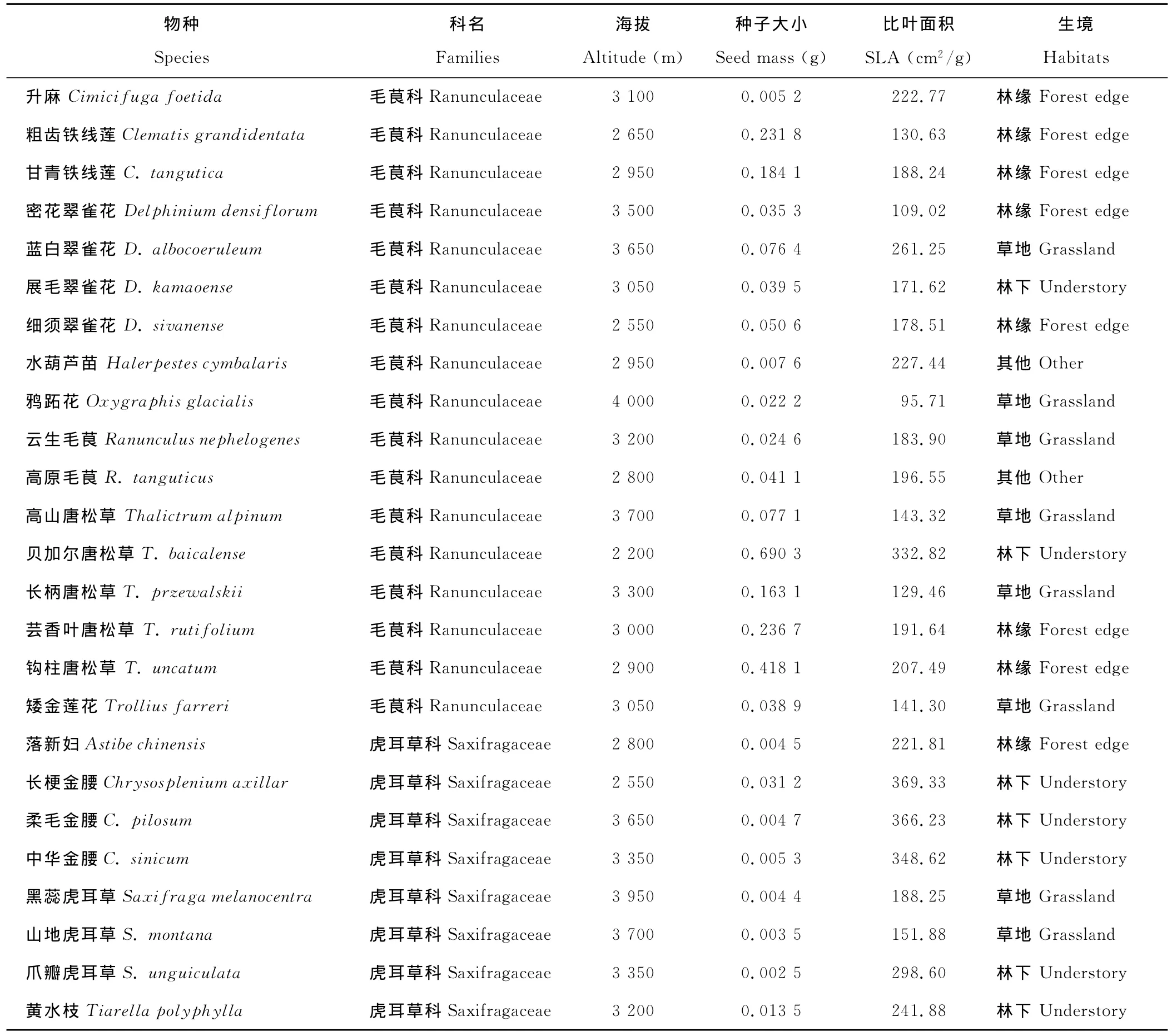
续表1 Continued
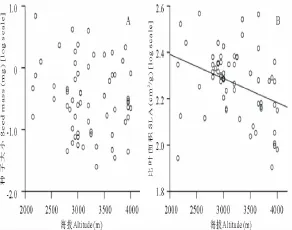
图1 海拔对63种植物种子大小和比叶面积的影响Fig.1 Effect of altitude on seed mass(A)and specific leaf area(B)of 63 plant species
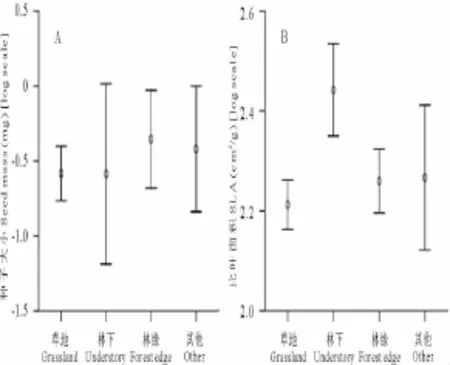
图2 不同生境下的植物种子大小和比叶面积Fig.2 Seed mass(A)and specific leaf area(B)in different habitats
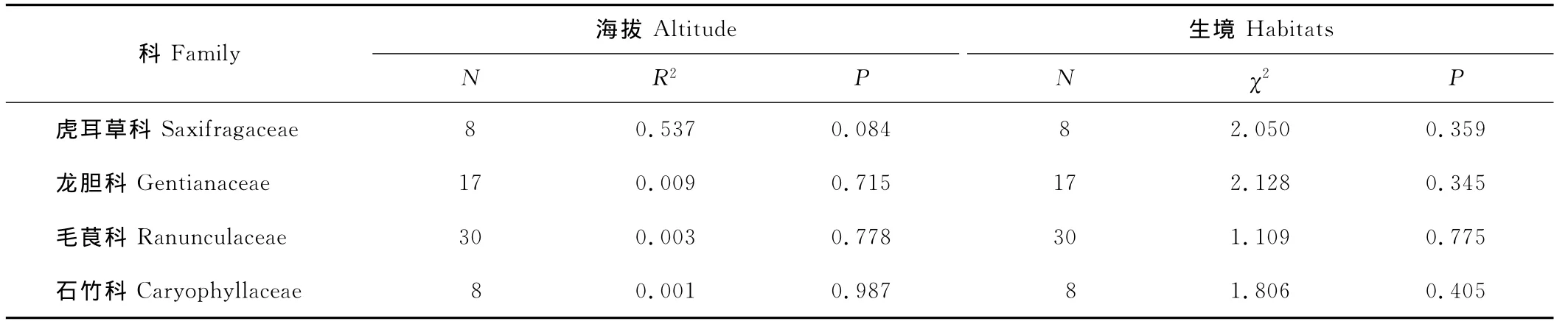
表2 科内种子大小与海拔和生境的关系Table 2 Relationship between seed mass and altitude or habitats within family
2.3 海拔对不同生境下植物种子大小的影响
协方差分析显示海拔和生境对种子大小变异均无显著影响(表3),这与单因素方差分析结果相似。然而,当用线性回归分析海拔对不同生境下植物种子大小的影响时,发现在林下环境中,种子大小与海拔有显著的负相关关系(R=-0.687,P=0.049),即种子大小随海拔的升高有减小的趋势;在草地(R=0.045,P=0.822)、林缘(R=-0.087,P=0.733)及其他生境(R=0.436,P=0.280)中海拔对种子大小没有影响(图3)。
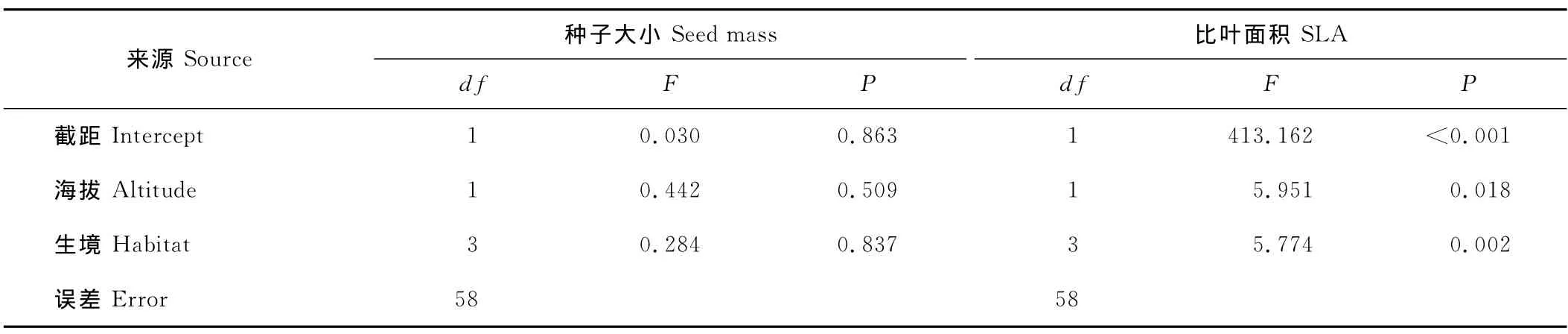
表3 海拔和生境对种子大小和比叶面积的协同影响Table 3 Combined effects of altitude and habitat on seed mass and SLA
2.4 比叶面积和海拔以及生境之间的关系
整体来看,63种植物的比叶面积和海拔之间呈显著的负相关关系(R=-0.351,P=0.005),即比叶面积随着海拔的升高而显著减小(图1 B)。另外,石竹科和虎耳草科植物的比叶面积不受海拔梯度的影响,而龙胆科和毛茛科植物的比叶面积与海拔显著负相关,即其比叶面积随海拔的升高显著减小(表4)。
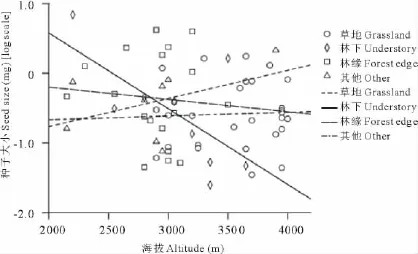
图3 不同生境下海拔对种子大小的影响Fig.3 Effect of altitude on seed size under different habitats
表4 科内比叶面积与海拔和生境的关系Table 4 Relationship between specific leaf area and altitude or habitats within family
比叶面积在不同生境条件下表现出明显的差异,LSD的多重比较显示,比叶面积在林下环境中最大(图2 B)。另外,Kruskal-Wallis H检验显示4个科植物在不同生境下的比叶面积均没有明显的变异趋势(表4),即生境类型对科内比叶面积没有显著影响。
2.5 海拔对不同生境下植物比叶面积的影响
协方差分析显示海拔和生境对植物比叶面积变异均有显著影响(表3),这与单因素方差分析结果相同。当用线性回归分析海拔对不同生境下植物比叶面积的影响时,发现在草地生境中随着海拔升高比叶面积显著减小(R=-0.479,P=0.010),而在林下(R=-0.154,P=0.692)、林缘(R=0.478,P=0.763)和其他(R=-0.478,P=0.231)生境下海拔对比叶面积没有显著影响(图4)。
3 讨论
3.1 种子大小和比叶面积的关系
作为植物功能性状的2个重要组成部分,有关比叶面积和种子大小关系的研究并不多。仅有的研究[18,19]发现具有较大种子的植物一般具有较小的比叶面积,并认为产生这种结果的原因可能与植物的生活史策略有关,即常绿的、寿命长久的植物种类(典型的K-对策者)一般具有较低的比叶面积、较长的叶片寿命和较低的潜在相对生长率,并具有较大的种子;而短命的植物(典型的r-对策者)一般具有较高的潜在相对生长率和比叶面积,并具有较小而多的种子,因此,比叶面积和种子大小间的关系可能是间接的,它们通过植株和叶片寿命以及潜在生长率而发生联系。本研究在青藏高原也发现了这种负相关(边际显著),表明比叶面积和种子大小间的这种由植物生活史策略(如K-r对策等)介导的联系具有普适性。
3.2 种子大小与海拔和生境的关系
一般情况下,环境不同,物种间或同一物种的不同种群间都可能存在种子大小的变异。然而,从整体上看,本研究结果并没显示海拔梯度和生境类型对青藏高原东部这4个典型植物科的种子大小有明显的影响,其可能的解释有以下两点。第一,高海拔地区严酷的环境压力选择大种子及其幼苗生存,但较低的净初级生产力(NPP)又不利于种子发育,这2个截然相反的压力很多时候可能会保持平衡,使得种子大小随海拔没有明显的变化趋势[5-8]。第二,本实验所用的材料全部为草本植物,并不包括本区系内的木本植物。草本植物本身选择大种子受到限制,因为草本植物没有木质化结构,植物茎尤其是末端所能承受的重量(大部分为种子的重量)有限,这样就内在地限制了草本植物的种子大小[20]。
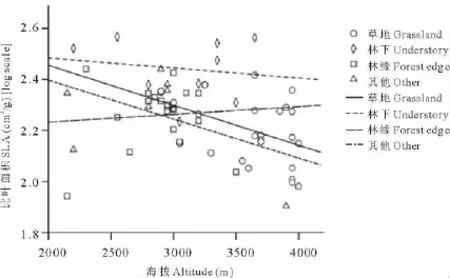
图4 不同生境下比叶面积随海拔的变异Fig.4 Effect of altitude on specific leaf area in different habitats
值得注意的是,尽管海拔和生境整体上对种子大小都没有影响,但是种子大小在生境和海拔的协同作用下却表现出了差异。本研究结果表明,林下生长植物的种子大小随海拔梯度有相对大的变异,其原因可以用种子大小在群落学中的作用来解释。一般而言,大种子植物(尤其植物在幼苗阶段)对光照、土壤水分和营养有较强的竞争能力,也更能忍受资源(光、土壤营养等)的缺失。在低海拔的森林中,建群种主要以各种槭树和栎树等大种子植物为主,在这类植物的竞争压力下,林下的草本植物必须有较大的种子才不至于被排斥出去而实现共存;相反,在高海拔的灌丛中,建群种主要以各种杜鹃花和金露梅等小种子植物为主,林下的草本植物可以有不太大的种子便不会被木本排斥出去而实现共存。
3.3 比叶面积与海拔和生境的关系
与种子大小不同,青藏高原东缘这4个常见植物科的比叶面积随海拔升高显著减小,其原因可能是随着海拔的升高,光照强度增大,环境变得严酷,叶中很大一部分物质用于构建保卫结构(防虫食、防止过度失水等)或者增加叶肉细胞密度,常形成厚度较大而面积较小即比叶面积较小的叶片[10]。另外,比叶面积在不同生境条件下也表现出明显的差异,表现出遮阴越大其值越大的特点。这可能是因为在遮阴环境下,植物从种子萌发到幼苗建植都处在高大木本植物的“保护”下,植物叶不需要面对强光、强风、动物践踏和虫食等外界压力,因此也不需要构建强大的保护组织。相反,由于遮阴使单位叶面积能获得的光资源变少,植物通常发育厚度薄而表面积大的叶片来增加自身的光捕获能力[19]。
值得注意的是,并不是所有生境下的比叶面积随海拔都有明显的变化,本研究结果显示,植物的比叶面积只在草地中随海拔变异显著。草地的生境相对开阔,光照和风可以直接作用于植物叶片表面,因此,随着海拔升高,太阳光辐射、紫外线、风速明显变强,草地上的植物唯有改变比叶面积才能适应如此的环境变化;相反,在相对郁闭的林下、林缘环境中,木本植物的躯干和冠层阻挡了部分的太阳光辐射和紫外线,减弱了风速,并创造了微环境而减弱巨大的昼夜温差,因此,在这种木本植物的“保护”下,林下和林缘草本所面临的太阳光辐射、紫外线和风速随海拔变化并不明显,其比叶面积随海拔的变化也不明显。然而,这种解释并不适用于同样开阔的其他生境中的植物。“其他”生境在实验中多是干扰严重或资源贫瘠的生境,可能在这种生境下光照不是植物适应环境时的主要压力,植物并不是选择比叶面积,而是选择其他功能性状的变异来适应海拔的变化,至于是选择哪些功能性状并不清楚,有待进一步研究。
3.4 科水平上的变异
根据本研究结果,青藏高原东缘这4个常见植物科科内的种子大小变异均不依赖于海拔和生境(表2),其原因可能是种子大小变异受系统发育影响,具有系统发育上的保守性。一些研究表明种子大小变异本身受到与分类学上相联系的特征地制约,包括心皮数、胎座类型、有无胚乳和胚珠类型等,这些特征是不易改变的,在科内以保守的方式表达,使得科内种子大小变异独立于环境变化[21-23]。相反地,这4个常见植物科科内比叶面积随海拔变异却出现科间差异(表4),表现为龙胆科和毛茛科植物的比叶面积与海拔显著负相关而另两科不相关,这一结果暗示在宏观地理因素(如海拔)的选择压力下,不同植物类群可能会采取不同的生存策略,即改变不同的植物性状,来适应这一压力。另外,本研究还发现,没有一个科的比叶面积和生境有明显关系,这一结果可能表明微观的地理因素(如生境)还不足以对科水平上的种子大小或比叶面积产生较大的影响。
本实验中所采集的物种并没有包含该科在青藏高原东部分布的全部物种,也没有考虑诸如水分、温度等气候因子的影响,有一定的局限性。考虑到本实验中的材料都是本区最典型和最常见的一些植物,本研究结果对研究本区域植物种子大小和比叶面积的变异有一定的参考价值。
[1] 张世挺,杜国祯,陈家宽.种子大小变异的进化生态学研究现状与展望[J].生态学报,2004,23(2):353-364.
[2] Hodgson J G,Mackey J M L.The ecological specialization of dicotyledonous families within a local flora:some factors constraining optimization of seed size[J].New Phytologist,1986,104:497-515.
[3] 郭淑青,齐威,王玉林,等.青藏高原东缘海拔对植物种子大小的影响[J].草业学报,2010,19(1):50-58.
[4] 柯君,王慧春,周华坤,等.三江源区高寒草甸43种植物繁殖体质量比较[J].草业科学,2010,27(3):15-20.
[5] Pluess A R,Schütz W,Stcklin J.Seed weight increase with altitude in Swiss Alps between related species but not among population of individual species[J].Oecologia,2005,144:55-61.
[6] Baker H G.Seed weight in relation to environmental conditions in California[J].Ecology,1972,53:997-1010.
[7] Bu H Y,Ch X L,Xu X L,etal.Seed mass and germination in an alpine meadow on the eastern Tsinghai-Tibet Plateau[J].Plant Ecology,2007,191:127-149.
[8] Bondeau A,Kicklighter D W,Kaduk J.Comparing global models of terrestrial net primary productivity(NPP):importance of vegetation structure on seasonal NPP estimates[J].Global Change Biology,1999,5:35-45.
[9] Wright I J,Cannon K.Relationships between leaf lifespan and structural defences in low nutrient,sclerophyll flora[J].Functional Ecology,2001,15:351-359.
[10] Reich P B,Ellsworth D S,Walters M B,etal.Generality of leaf train relationships:a test across six biomass[J].Ecology,1999,80:1955-1969.
[11] Wright I J,Westoby M,Reich P B.Convergence towards higher leaf mass per area in dry and nutrient poor habitats has different consequences for leaf life span[J].Journal of Ecology,2002,90:534-543.
[12] Westoby M A.Leaf-height-seed(LHS)plant ecology scheme[J].Plant and Soil,1998,199:213-227.
[13] 韩立辉,尚占环,任国华,等.青藏高原“黑土滩”退化草地植物和土壤对秃斑面积变化的响应[J].草业学报,2011,20(1):1-6.
[14] 曹文侠,徐长林,张德罡,等.杜鹃灌丛草地土壤容重与水分特征对不同休牧模式的响应[J].草业学报,2011,20(3):28-35.
[15] 杨霞,梁艳,陈学林,等.青藏高原东缘地区常见植物种子大小变异研究[J].生态科学,2007,26(6):483-489.
[16] 冯燕,王彦荣,胡小文.水分胁迫对幼苗期霸王叶片生理特性的影响[J].草业科学,2011,28(4):577-581.
[17] 冯燕,王彦荣,胡小文.水分胁迫对两种荒漠灌木幼苗生长与水分利用效率的影响[J].草业学报,2011,20(4):293-298.
[18] Garnier E.Growth analysis of congeneric annual and perennials grass species[J].Journal of Ecology,1992,80:665-675.
[19] Reich P B,Walters M B,Ellsworth D S.Leaf life-span in relation to leaf,plant,and stand characteristics among diverse ecosystems[J].Ecological Monographs,1992,62:365-392.
[20] Westoby M,Falster D S,Moles A T,etal.Plant ecological strategies:some leading dimensions of variation between species[J].Annual Review of Ecology and Systematics,2002,33:125-159.
[21] Leishman M R,Westoby M,Jurado E.Correlates of seed size variation-a comparison among five temperate floras[J].Journal of Ecology,1995,83:517-529.
[22] Lord J,Westoby M,Leishman M.Seed size and phylogeny in six temperate floras:constraints,niche conservatism,and adaptation[J].American Naturalist,1995,146:349-364.
[23] Crisp M D,Arroyo M T K,Cook L G,etal.Phylogenetic biome conservatism on a global scale[J].Nature,1995,458:754-756.
