早期运动训练对帕金森小鼠中脑和纹状体的影响:自噬与线粒体动力学关系的研究
2012-02-27姜宁曹玮宋超郭晶晶刘洪涛张勇
姜宁曹玮宋超郭晶晶刘洪涛 张勇,
1军事医学科学院卫生学环境医学研究所(天津300050)2天津体育学院天津市运动生理与运动医学重点实验室
帕金森症(Parkinson’s disease,PD),又称震颤麻痹,是一种与年龄相关的进行性神经退行性运动病变。随着我国人口老龄化,该病的患病率呈增加态势。尽管国内外对PD进行了大量研究,但PD的发病机制尚未明确。自噬(Autophagy)是胞浆大分子物质和细胞器在溶酶体中批量降解并涉及亚细胞膜重构的动力学过程。自噬在改善损伤的细胞功能方面的作用已引起广泛关注[1]。研究表明[2],自噬在清除与神经性疾病相关的细胞器,尤其是在PD的发生、发展过程中发挥重要作用。国外流行病学调查显示,早期规律从事体育运动或体力劳动的人口,PD发病率显著降低[3]。本课题组前期研究也表明,运动预适应可以提高PD小鼠脑线粒体功能,延缓或减少中脑多巴胺神经元的死亡,增加中脑黑质细胞数量[4]。但其确切机制还需要进一步研究。
本实验通过建立小鼠PD模型,观察早期运动训练对PD小鼠中脑和纹状体线粒体呼吸功能、ATP酶合成活力和自噬相关蛋白的变化,探讨运动训练对脑组织的神经保护作用机制,为运动防治帕金森等神经退化性疾病提供新的理论依据。
1 材料和方法
1.1 实验动物和分组
雄性C57BL/6小鼠(22~24g),6~8周龄。日光照时间12小时,室温20℃。随机分为4组:安静+生理盐水组(N组)、运动+生理盐水组(NE组)、安静+ MPTP(1-甲基-4-苯基-1,2,3,6-四氢吡啶)组(M组)和早期运动训练+MPTP组(ME组),每组15只。
1.2 训练方案和帕金森病模型制作
运动组小鼠进行2天适应性跑台训练 (5 m/ min,10 min),以后每天上午进行1次中等强度跑台训练(12 m/min,20 min,6天/周),连续6周。训练结束后,安静+MPTP组和早期运动训练+MPTP组接受中等剂量MPTP注射(30 mg/kg×2次,ip,间隔16小时),其余两组小鼠在相同时间、以相同方式接受同等剂量的生理盐水注射。每次小鼠腹腔内注射MPTP 10 min后,出现静止性振颤、运动减少、后肢僵硬竖尾、立毛等行为学表现,约持续30 min后振颤消失,但运动减少仍然存在。接受生理盐水注射的动物无上述表现。于造模后第7天取材。
1.3 脑组织线粒体提取
实验小鼠取材前一天禁食大于12小时后,脱臼处死,按小鼠脑立体定位图取中脑和纹状体。按照5 ml/g组织的用量加入分离介质,在预冷的玻璃匀浆器中上下均匀匀浆10次,匀浆液离心(1200g、4℃、10 min)。将上清4℃暂放,取沉淀,加入分离介质,重复匀浆离心,取上清,与前次匀浆后上清合并离心(12000g、4℃、10 min),弃上清,沉淀即为粗制线粒体。每克组织加入保存介质约80 μl,取样测定蛋白浓度。采用考马斯亮蓝法测定,用280 nm下校正的BSA作标准曲线,室温下测定。
1.4 线粒体呼吸功能检测
线粒体呼吸功能采用 Clark氧电极法,用Oxytherm测氧仪(Hansatech-Instruments Co.,UK)测定态3呼吸速率(state 3)、态4呼吸速率(state 4)及呼吸控制比(respiratory control ratio,RCR)。反应体系 1 ml,30℃恒温。反应介质为:130 mM KCl,10 mM Hepes,1 mM EDTA,2.5 mM KH2PO4,1.5 mg defatted BSA,pH 7.4。线粒体蛋白浓度为1 mg/ml。加入0.1 mmol/L苹果酸和1 mmol/L谷氨酸启动态4呼吸。平稳后加入200 nmol ADP启动态3呼吸。ADP耗尽后重又回到态4呼吸。记录仪记录耗氧曲线。由此可算出态3、态4呼吸以及RCR(态3/态4)。
1.5 线粒体ATP合成酶活力测定
采用20/20n型发光仪荧光素-荧光素酶发光法测定ATP合成酶活力。反应温度为25℃,在0.5 ml反应体系中含有0.5 mM EDTA,10 mM HEPES,5 mM磷酸盐缓冲液,2.5 mM MgCl2,0.5 M苹果酸/0.25 M谷氨酸,100 μl荧光素-荧光素酶,50 μg线粒体。记录以上体系的发光强度为本底,加入4 μl ADP启动反应,记录发光强度在不加线粒体和ADP的平行实验中,加入1 nM ATP,记录发光强度,标定ATP合成量。酶活力单位为nmol/min·mg pro。
1.6 自噬相关蛋白Beclin 1、LC3及线粒体分裂相关蛋白Drp1、Fis1 mRNA表达定量测定
另取中脑和纹状体组织。Trizol Reagent抽提总RNA,经紫外分光光度计检测OD值及OD260/280比值,计算RNA浓度。按照RevertAid First Strand cDNA Synthesis Kit(Fermentas)说明书进行逆转录反应。Real-time PCR采用TAKARA的Mix Taq试剂盒于ABI StepOne Real-time PCR仪进行。引物序列见表1。所有试剂均以DEPC处理的双蒸水配制。PCR反应体系:SYBR@Premix Ex Taq(2×)10μl,PCR Forward Primer (10μM)0.4μl,PCR Reverse Primer(10μM)0.4μl,ROX Reference Dye (50×)0.4μl,cDNA 2μl,ddH2 O6.8μl。

表1 荧光定量PCR引物序列
1.7 Western Blot测定 Beclin 1、LC3-II、Drp1、Fis1蛋白表达
RIPA法提取小鼠骨骼肌总蛋白,考马斯亮蓝法测定上清蛋白浓度。使用Tris-甘氨酸SDS进行聚丙烯酰胺凝胶电泳。采用湿法电转印将蛋白转到PVDF膜上。用含有5%BSA的抗体稀释液按比例(分别按1:10000、1:1000、1:5000、1:3000)稀释一抗,4℃孵育过夜。然后用1×TBST洗膜3次,每次5分钟。用含有5%脱脂奶粉的抗体稀释液按照1:10000的比例稀释二抗,室温孵育1小时后用1×TBST洗膜3次,每次5分钟,加KPL发光底物,于暗室曝光,胶片经显影、定影后进行扫描,以β-tubulin为内参经Quantity ONE(BIO-RAD)软件对目的蛋白质进行光密度相对定量分析。
1.8 统计学分析
所有实验数据由SPSS13.0统计软件处理,计算均值和标准差(±s),组间比较采用双因素方差分析(UNIANOVA-General Linear Models),显著性水平为P<0.05。
2 结果
2.1 线粒体呼吸功能
如图1所示,M组小鼠中脑和纹状体线粒体态3呼吸显著低于N组(P<0.01)。ME组态3呼吸显著高于M组(P<0.01)。NE组态3呼吸较N组有所提高,但无显著性。各组间态4呼吸变化无显著性。如图2所示,M组、ME组小鼠中脑线粒体RCR显著低于N组(P<0.01,P<0.05),ME组显著高于M组(P<0.01)。NE组和N组相比RCR变化无显著性。

图1 中脑和纹状体线粒体态3和态4呼吸速率
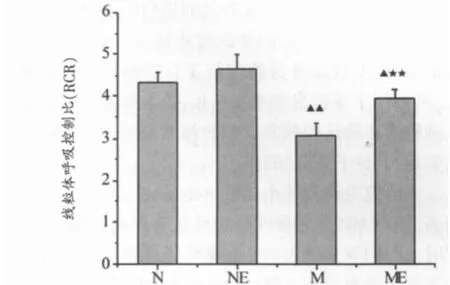
图2 中脑和纹状体线粒体呼吸控制比(RCR)
2.2 中脑和纹状体线粒体ATP酶合成活力
如图3所示,与N组相比,NE组小鼠中脑和纹状体线粒体ATP酶合成活力显著升高(P<0.05);M组显著降低 (P<0.01)。ME组较M组显著升高(P <0.05)。
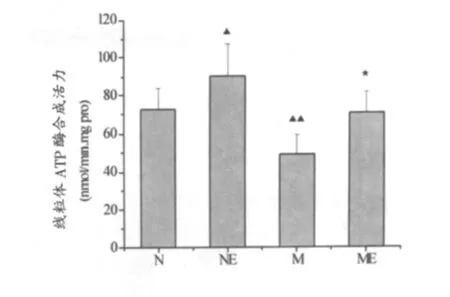
图3 中脑和纹状体线粒体ATP酶合成活力
2.3 中脑和纹状体组织Beclin 1,LC3基因和蛋白表达
如图4所示,与N组相比,NE组、M组、ME组各组Beclin 1 mRNA均显著升高(P<0.01),分别是N组的2倍、2.54倍和4.47倍;LC3 mRNA显著升高,分别是N组的1.7倍、1.6倍、2.09倍。与M组相比,ME组Beclin 1 mRNA显著升高,为M组的1.76倍;LC3 mRNA显著升高(P<0.01),为M组的1.31倍。如图5所示,与N组相比,NE组、M组、ME组各组Beclin 1蛋白含量均显著升高,分别是N组的1.25倍、1.39倍、1.65倍。与N组相比,M组,ME组LC3-II蛋白含量显著增高,分别为N组的1.45倍、1.77倍。与M组相比,ME组Beclin 1蛋白、LC3-II蛋白均显著增高(P<0.01)。
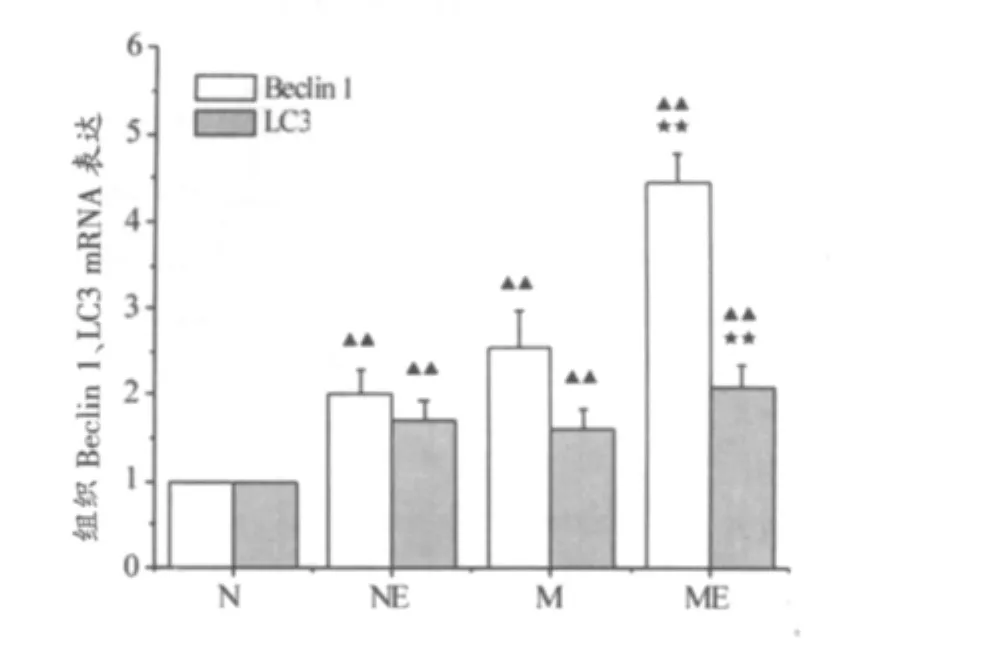
图4 中脑和纹状体组织Beclin 1、LC3 mRNA表达

图5 中脑和纹状体组织Beclin 1、LC3-II蛋白表达
2.4 线粒体分裂蛋白Fis1和Drp1的mRNA和蛋白表达
如图6所示,与N组相比,NE组、M组、ME组各组Fis1mRNA均显著升高,分别是N组的2倍、1.91倍、3.41倍;与N组相比,M组和ME组Drp1 mRNA显著升高,分别是N组的1.62倍、1.95倍。和M组相比,ME组Fis1 mRNA显著升高(P<0.01)。如图7所示,与N组相比,NE组、M组、ME组各组Fis1蛋白含量显著升高,分别是N组的1.43倍、1.29倍和1.53倍;与N组相比,M组、ME组Drp1蛋白含量显著增高(P<0.01),分别为N组的1.42倍和1.58倍。与M组相比,ME组Fis1蛋白显著升高(P<0.01)。

图6 中脑和纹状体组织Fis1、Drp1 mRNA表达
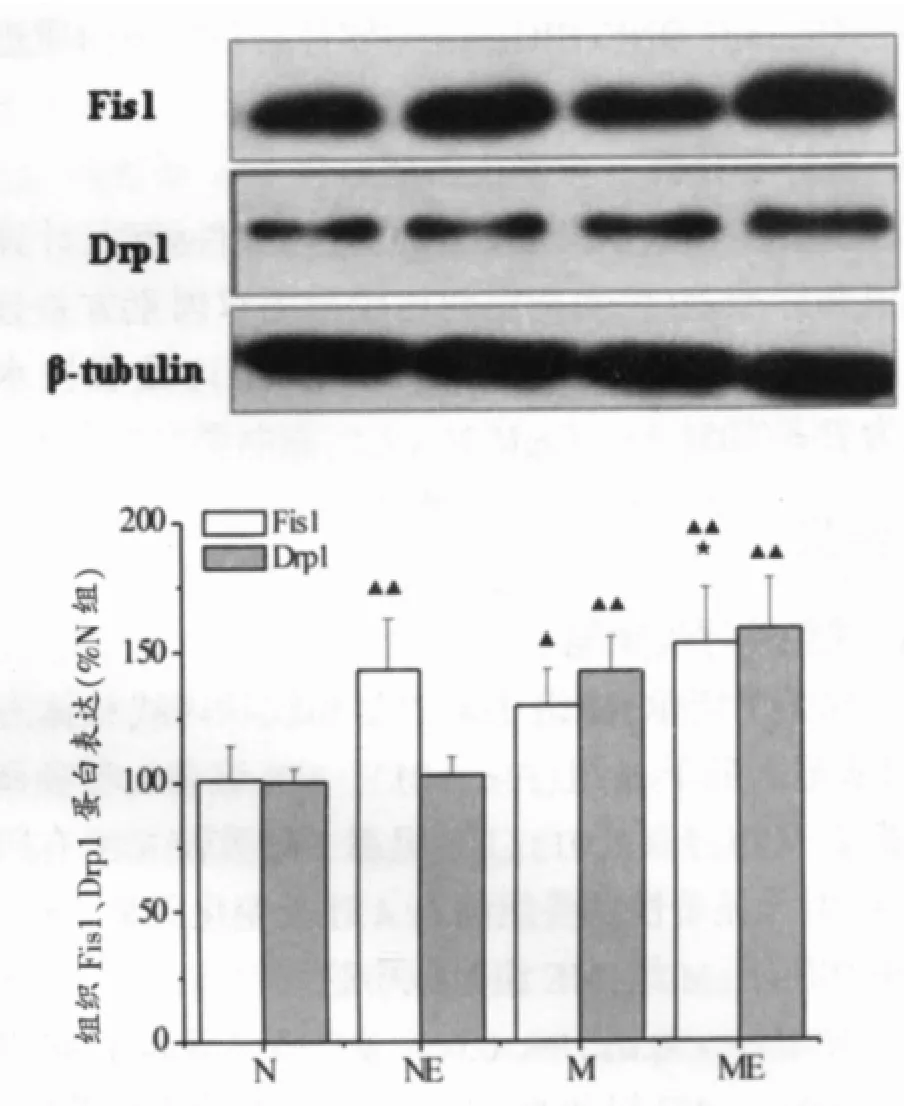
图7 中脑和纹状体组织Fis1、Drp1蛋白表达
3 讨论
自噬是广泛存在于真核细胞内的一种溶酶体依赖性的降解途径,降解内容包括长寿命蛋白质和受损的细胞器。它是一把双刃剑,适度自噬可有效降解胞质大分子和细胞器,为蛋白质合成提供能量,在机体处于氧化应激环境胁迫时发挥保护作用,但其过度活跃或功能低下也会引发自噬应激,从而对细胞造成伤害[5]。
微管相关蛋白1轻链3(microtubule-associated protein 1 light chain 3,LC3)为真核细胞中第一种被认定的与自噬小泡膜形成有关的蛋白[6],LC3-I合成后通过对其羧基末端的甘氨酸残基进行剪切修饰,形成LC3-II,LC3-II最终定位于自噬前体和自噬体上,其可用于评价自噬水平。Beclin1是第一个在哺乳动物中被确认的调节自噬的相关基因,对自噬的发生起到“分子开关”的作用[7]。
本研究发现,PD小鼠脑Beclin1、LC3-II表达均增高,提示MPTP的毒性作用可显著提高自噬水平。同时,我们观察到MPTP的毒性作用造成线粒体功能下降,表现为态3呼吸、RCR和ATP酶合成活性下降。造模使用的MPTP浓度为30 mg/kg,为中等剂量,虽可造成明显帕金森症状,但其行为学改变在1周可恢复正常。因此,该模型造成的脑自噬水平的提高是机体对外界损伤的代偿反应,可发挥保护作用。
在单纯运动训练的小鼠脑中,参与调控自噬的Beclin1水平提高,早期运动训练的PD小鼠脑自噬蛋白Beclin1、LC3-II较PD模型组显著升高。不仅如此,伴随自噬水平的提高,也观察到早期运动训练的PD模型小鼠脑线粒体功能显著提高,表现为态3呼吸提高,ATP生成增多。统计结果显示,预运动训练与MPTP造模有显著交互作用,即早期运动训练可以提高小鼠中脑和纹状体线粒体对MPTP毒性的抵抗,改善线粒体功能。研究提示:早期运动训练初步启动自噬,此时如果中脑和纹状体遭遇MPTP的毒性作用,即可及时调控自噬过程,改善线粒体功能。
自噬,起初被认为是非选择性的活动。然而,进一步研究发现,细胞在应对多种刺激时可通过自噬选择性降解损伤的线粒体,这种现象在酵母细胞和哺乳动物细胞普遍存在。当酵母细胞营养匮乏时,其自噬水平上调,快速降解线粒体,以节约能量消耗,在哺乳动物中,也同样可以发现这一情况[8]。Goldman等[9]认为细胞线粒体发生损伤时,线粒体自噬活性增强,从而使得受损线粒体被及时降解,这也是体内清除受损线粒体的主要途径。
目前,关于损伤的线粒体如何通过自噬途径被清除,及其相关信号通路间的相互作用机制尚无定论。Twig等[10]针对线粒体融合分裂在线粒体自噬中的作用进行了研究,发现线粒体在经历了融合之后会被分裂成为两个子个体,一部分发生进一步融合,另一部分则不会。有融合能力的子线粒体会在融合发生之前首先恢复膜电位(ΔΨm),而另一部分子线粒体无法恢复ΔΨm,进而被自噬清除,即线粒体自噬参与了线粒体质量控制过程。
Parone等[11]研究发现,敲除了线粒体分裂相关蛋白Drp1的Hela细胞,线粒体呼吸功能丧失,ATP浓度下降,细胞周期缩短,mtDNA部分缺失,而LC3-Ⅱ表达增加,提示线粒体自噬在敲除Drp1的细胞中被抑制。Arnoult等[12]同样发现,Drp1过表达可以驱动线粒体分裂,从而使线粒体消失。而线粒体分裂的另一相关蛋白Fis1突变也可导致线粒体结构形态及功能的改变,引起其功能障碍,导致线粒体被清除[13]。这些证据都提示,线粒体分裂可能是线粒体吞噬前发生的必要步骤,即线粒体动力学变化与线粒体相关自噬有着密切联系。
本研究发现,运动训练可上调中脑和纹状体组织的线粒体分裂水平。有研究发现,低剂量MPTP可促进Drp1依赖的线粒体片段化[14],而线粒体趋向分裂可能有利于加强ATP合成,减少ATP的消耗,满足细胞对能量需求的增加[15]。
目前,对运动训练在延缓PD发病中的作用机制尚不清楚。本研究发现早期运动训练可提高PD小鼠对MPTP所致线粒体功能损害的抵抗作用。在这一过程中,伴随着自噬水平提高,线粒体分裂水平亦显著升高,提示线粒体动力学的改变参与了早期运动训练对PD小鼠自噬水平增高的调控过程。但其相关机制还需进一步研究。
4 总结
中等剂量的MPTP可损害中脑和纹状体线粒体功能,使自噬水平代偿性增高;早期运动训练可初步启动中脑和纹状体自噬。而当MPTP损害发生时,这一过程可及时调控线粒体分裂,促进自噬水平上调,进而改善中脑和纹状体线粒体功能,在运动防治帕金森病中发挥神经保护作用。
[1] Mizushima N,Klionsky DJ.Protein turnover via autophagy:implications for metabolism.Annu Rev Nutr,2007,27:19-40.
[2] Werling K.Role of autophagy in the pathogenesis of liver diseases.Orv Hetil,2011,152(49):1955-1961.
[3] Chen H,Zhang SM,Schwarzschild MA,et al.Physical activity and the risk of Parkinson disease.Neurology,2005,64(4):664-669.
[4] 薛宏斌,张勇,刘洪涛,等.早期运动训练通过增强小鼠脑线粒体呼吸功能预防MPTP神经毒性作用.中国运动医学杂志,2007,26(7):402-406.
[5] Shintani T,Klionsky DJ.Autophagy in health and disease:a double-edged sword.Science,2004,306(5698):990-995.
[6] Tanida I,Ueno T,Kominami E.LC3 conjugation system in mammalian autophagy.Int J Biochem Cell Biol,2004,36(12):2503-2518.
[7] Aita VM,Liang XH,Murty VV,et al.Cloning and genomic organization of beclin 1,a candidate tumor suppressor gene on chromosome 17q21.Genomics,1999,59(1):59-65.
[8] Lemasters JJ.Selective mitochondrial autophagy,or mitophagy,as a targeted defense against oxidative stress,mitochondrialdysfunction,and aging.Rejuvenation Res,2005,8(1):3-5.
[9] Goldman SJ,Taylor R,Zhang Y,et al.Autophagy and the degradation of mitochondria.Mitochondrion.2010,10(4):309-315.
[10]Twig G,Hyde B,Shirihai OS.Mitochondrial fusion,fission and autophagy as a quality control axis:the bioenergetic view.Biochim Biophys Acta,2008,1777(9):1092-1097.
[11]Parone PA,Da CS,Tondera D,et al.Preventing mitochondrial fission impairs mitochondrial function and leads to loss of mitochondrial DNA.PLoS One,2008,3(9):e3257.
[12]Arnoult D,Rismanchi N,Grodet A,et al.Bax/Bak-dependent release of DDP/TIMM8a promotes Drp1-mediated mitochondrial fission and mitoptosis during programmed cell death.Curr Biol,2005,15(23):2112-2118.
[13]Twig G,Elorza A,Molina AJ,et al.Fission and selective fusion govern mitochondrial segregation and elimination by autophagy.EMBO J,2008,27(2):433-446.
[14]Trimmer PA,Swerdlow RH,Parks JK,et al.Abnormal mitochondrialmorphology in sporadic Parkinson's and Alzheimer's disease cybrid celllines.Exp Neurol,2000,162(1):37-50.
[15]Meeusen S,McCaffery JM,Nunnari J.Mitochondrial fusion intermediates revealed in vitro. Science,2004,305(5691):1747-1752.
