利用线粒体DNA控制区部分序列分析不同地理群体大泷六线鱼遗传多样性
2012-01-11孟凡平李占东
李 莹, 王 伟, 孟凡平, 李占东
(1.大连海洋大学 农业部北方海水增养殖重点实验室, 辽宁 大连 116023; 2.东港出入境检验检疫局, 辽宁东港 118300)
利用线粒体DNA控制区部分序列分析不同地理群体大泷六线鱼遗传多样性
李 莹1, 王 伟1, 孟凡平1, 李占东2
(1.大连海洋大学 农业部北方海水增养殖重点实验室, 辽宁 大连 116023; 2.东港出入境检验检疫局, 辽宁东港 118300)
利用PCR技术扩增了6个野生地理群体大泷六线鱼(Hexagrammos otakii)线粒体DNA控制区的部分序列, 共获得 352bp碱基序列, 31尾个体定义了 22种单倍型。中性检验 Fu′sFs值为-0.14881(P<0.01)。分子变异分析(AMOVA)结果显示: 遗传分化指数Fst=0.7398(P<0.01), 73.98%的变异来自群体间, 26.02%的变异来自群体内。根据不同个体间的遗传距离构建了NJ和MP系统树, 两种方法获得了相似的进化树。其中青岛群体与旅顺群体的一部分个体单独成支, 其它群体聚为 1支。研究结果表明, 大泷六线鱼群体遗传多样性较为丰富, 群体间发生了较大的遗传分化, 证明线粒体DNA控制区基因可用于大泷六线鱼群体内及群体间遗传多样性的分析。
大泷六线鱼(Hexagrammos otakii); 野生地理群体; 线粒体DNA控制区部分序列; 遗传多样性
大泷六线鱼(Hexagrammos otakii), 也称为欧氏六线鱼、六线鱼等[1-2], 山东、辽宁沿海俗称黄鱼, 隶属于鲉形目(Scorpaeniformes)、六线鱼科(Hexagrammidae)、六线鱼属(Hexagrammos)[2], 主要分布于中国的辽宁和山东等地的近海多岩礁海区,日本、韩国和朝鲜等国的近海也有分布[3-4]。该鱼肉味鲜美, 营养价值高, 很受山东、辽宁沿海当地人民的欢迎, 除供我国市场外, 还可以出口。该鱼目前已成为北方海水养殖鱼类之一。日本于20世纪70年代就开始对其人工繁殖进行研究, 中国于20世纪80年代也开始研究。有关该鱼的生物学研究主要是在人工育苗[5-8], 营养成分分析[9-10], 生物学指标[11-12]等方面, 目前国内仅见刘奇[13]一篇关于大泷六线鱼群体遗传多样性方面的研究报道。
研究物种的遗传多样性有助于了解物种的进化历史和种群恢复潜力, 从而制定有效的保护和管理对策。线粒体DNA(mtDNA)具有母性遗传的特点, 它的拷贝数多、突变率高, 突变固定后形成的多态位点可反映群体遗传特征、种群分化和种属关系等特点,是系统进化和种群遗传研究中理想的分子标记[14-16]。控制区(D-loop)是线粒体上的非编码区, 进化速度快,适合群体水平的遗传变异分析[17-18]。以序列测定技术为支持, 鱼类线粒体 D-loop序列已被应用于多种鱼类群体遗传多样性的分析,例如, 四大家鱼[19]、云南 倒 刺鲃(Spinibarbus denticulatus yunnanensis)[20]、黄吻姬鲷(Pristipomoides multidens)[21]、雅罗鱼(Leuciscus leuciscus)[22]、铲颔鱼(Varicorhinus barbatulus)[23]、细鳞鲑(Brachymystax lenok)[24]、大口黑鲈(Micropterus salmoides)[25]、沙鳅(Botia superciliaris)[26]、银鲳(Pampus argenteus)[27]和裂腹鱼(Schizothorax kozlovi)[28]等。近年来, 大泷六线鱼的资源量日益减少, 本实验利用线粒体DNA控制区序列对 6个群体大泷六线鱼的遗传结构和变异进行分析, 旨在了解其遗传背景, 为加强对其遗传多样性的保护和开发, 合理利用其渔业资源提供基础性的资料。
1 材料与方法
1.1 样本采集
本实验所用野生大泷六线鱼分别采自青岛、丹东和大连等 6个地区。采样地的地理位置和样本数见图1和表1。
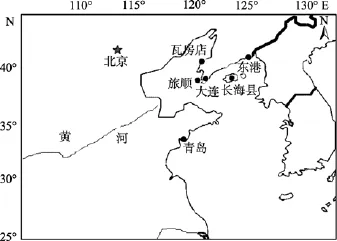
图1 大泷六线鱼群体的采样地点Fig. 1 Map of Hexagrammos otakii sampling sites
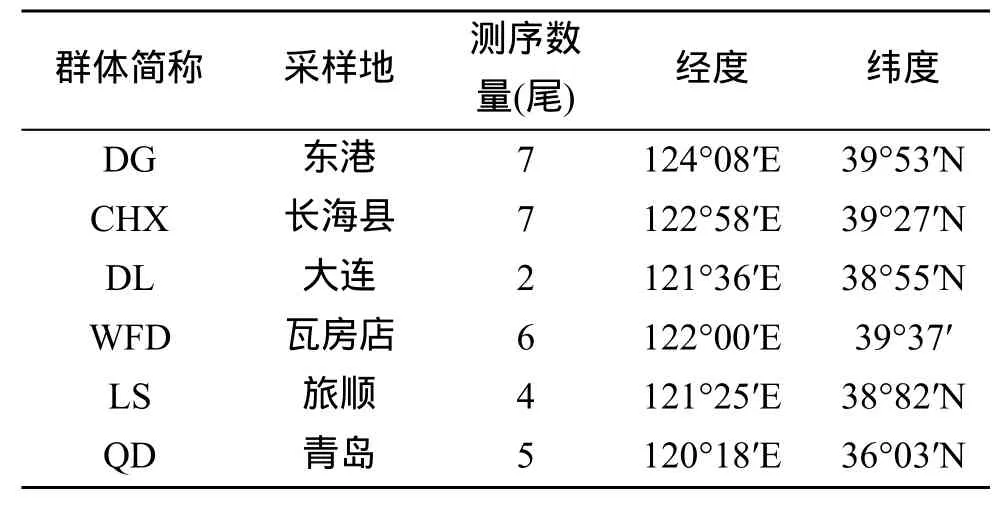
表1 大泷六线鱼群体简称、采样地、数量及地理坐标Tab. 1 Population codes, locality, numbers and geographical coordinates of Hexagrammos otakii
1.2 基因组DNA的提取
采用了一种快速提取DNA的方法。取50 mg左右鱼尾部肌肉组织加入匀浆液、SDS以及蛋白酶K,消化大约 2 h, 经乙酸钾溶液抽提, 两倍体积无水乙醇沉淀, 75%酒精洗涤后, 用适量 TE溶解, -20℃存放。
1.3 PCR扩增及测序
所用通用引物序列为 DL1(5’-TCA AAG CTT ACA CAG TCT TGT AAA CC-3’), DL2 (5’-CCT GAA GTA GGA ACC AGA TG -3’)。PCR反应使用大约50 ng基因组DNA为模板。反应体积为25 μL, 其中基因组 DNA2 μL, 10×PCR Buffer 2.5 μL, MgCl25.0 μmmol/L, dNTPs 0.5 mmol/ L, 引物各1 mol/L, Taq酶2U。PCR反应程序为: 95℃预变性3 min, 然后进行35个循环(94℃变性30 s, 56℃退火45 s, 72℃延伸1 min), 最后72℃延伸10 min。扩增产物经琼脂糖凝胶回收纯化, 送至上海鼎安生物科技有限公司进行测序。
1.4 数据分析
用 ClustalX1.83软件进行序列比对, 用Mega4.10软件统计变异位点和碱基组成, 计算群体遗传距离, 构建 NJ和 MP分子系统树。用DNASP4.10软件计算序列的单倍型数(h)、单倍型多样性指数(Hd)、核苷酸多样性指数(π)以及平均核苷酸差异数(K)。用Arlequin 3.11软件进行中性检验, 计算Fu′sFs值来推断群体发生扩张的历史。用分子变异分析方法(AMOVA)分析群体间遗传分化指数Fst并用排列测验法(permutation test)检测Fst的显著性。
2 结果
2.1 序列特征
PCR扩增得到约500 bp的碱基序列, 测序结果经比对剪切共获得 31尾大泷六线鱼线粒体 D-loop序列, 其5′端的长度为352 bp。T、A、C、G碱基的平均含量分别为37.0%、34.2%、14.1% 和14.8%。A+T 含量(71.1%)明显高于 G+C含量(28.9%), 表现出显著的碱基组成偏向性, 符合脊椎动物 mtDNA D-loop区碱基组成的特点。大连群体 T+A含量最高,高达71.9%, 而青岛群体最低(69.4%)。G+C 含量则相反, 青岛群体最高(30.6%), 大连群体最低(28.1%)。
31尾个体共定义了 22种单倍型(表 2): 群体间共享的单倍型有4个, 占单倍型总数的18.2% ,其中2个单倍型为3个群体所共有, 2个单倍型为两个群体共有。18种单倍型为单个群体特有, 占单倍型总数的81.8%。
2.2 遗传多样性
群体遗传多样性参数统计表明(表 3): 旅顺群体呈现出最高的核苷酸多样性指数(0.28938), 东港、大连和青岛群体呈现出最高的单倍型多样性指数(1.00000)。东港、长海县、瓦房店和青岛群体序列的 Fu′sFs值均为负, 并达到统计学的显著性差异,提示这4个群体的检验结果均偏离中性模式。
2.3 遗传结构
Kimura 2-paramter遗传距离和遗传分化指数Fst见表 4。6个群体间的平均遗传距离为 0.00739~0.66972, 其中青岛与瓦房店群体间的遗传距离最高,大连与瓦房店群体间的最低。6个群体间的遗传分化指数Fst为-0.04828~0.97565, 其中青岛与长海县群体间的遗传分化指数最高, 长海县与瓦房店群体间的最低。分子变异分析(AMOVA)结果显示(表5),Fst=0.73983(P<0.01), 即 73.98%的变异来自群体间,26.02%的变异来自群体内, 表明大泷六线鱼 6个群体间发生了显著的遗传分化。
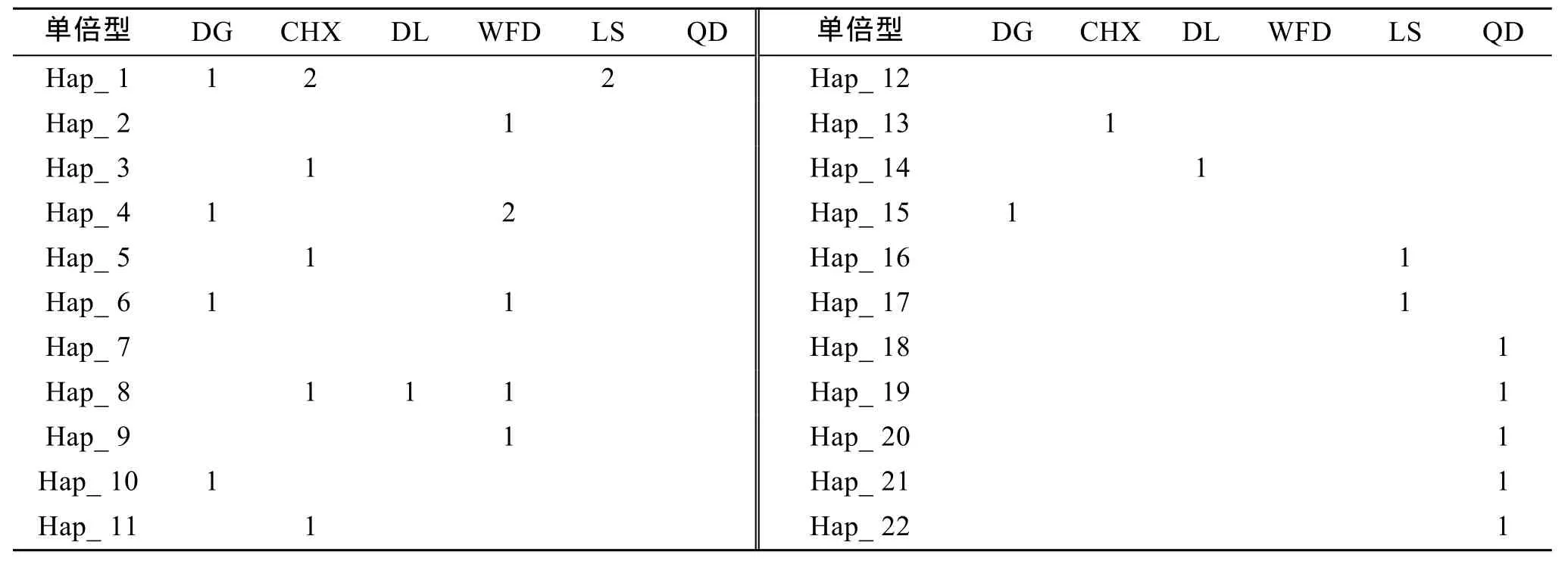
表2 大泷六线鱼mtDNA控制区单倍型在各群体中的分布Tab. 2 The distribution of haplotypes in the six populations of Hexagrammos otakii
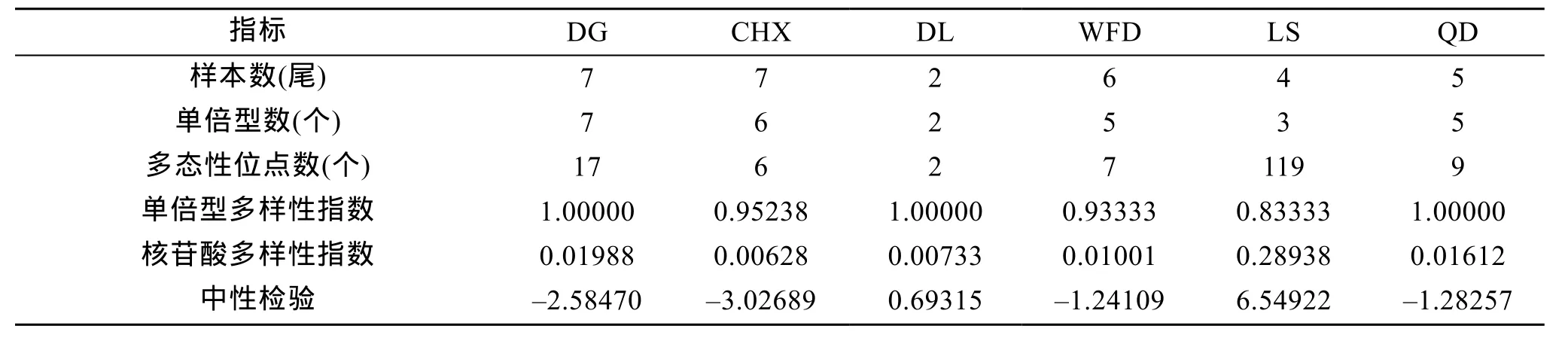
表3 大泷六线鱼群体遗传多样性参数Tab. 3 Intra-populational genetic diversity in different populations of Hexagrammos otakii
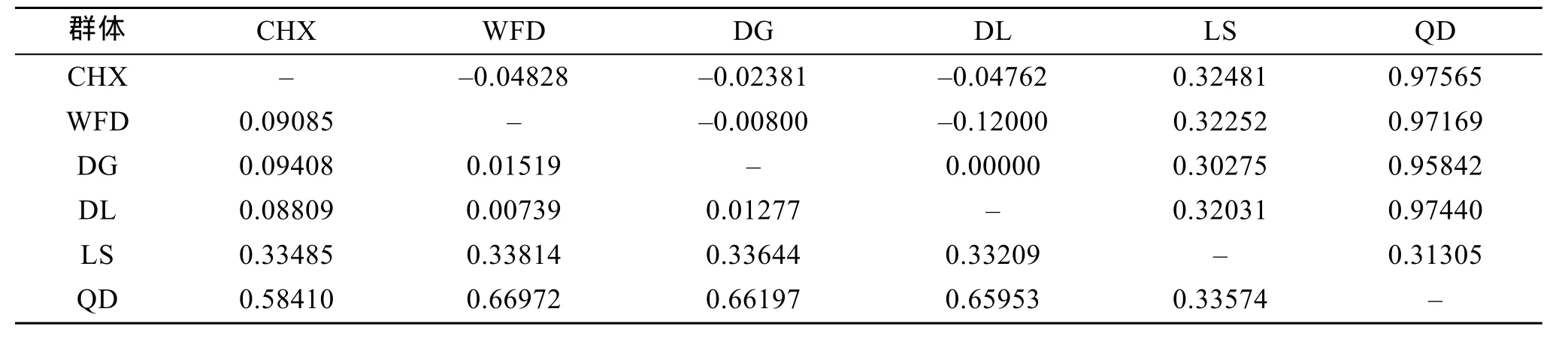
表4 6个大泷六线鱼群体间的遗传距离(对角线下方)与遗传分化指数Fst(对角线上方)Tab. 4 Pairwise genetic distance (below diagonal) and Fst (above diagonal) among the six populations of Hexagrammos otakii
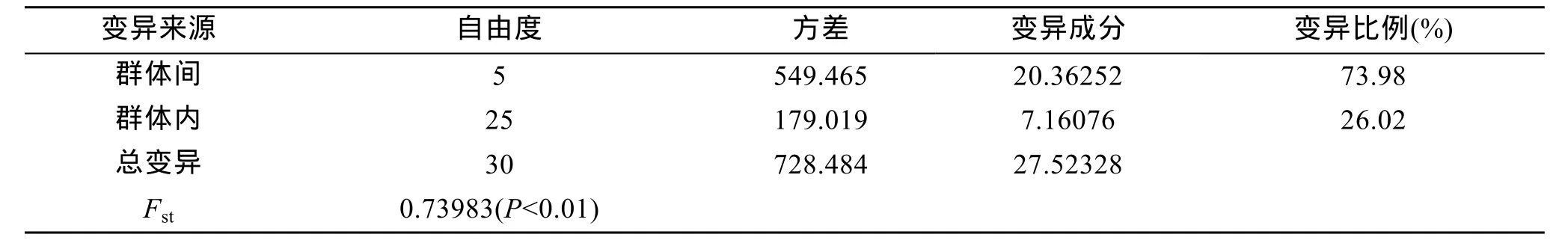
表5 大泷六线鱼群体间遗传差异的分子方差分析Tab. 5 Analysis of molecular variance (AMOVA) among the populations of Hexagrammos otakii
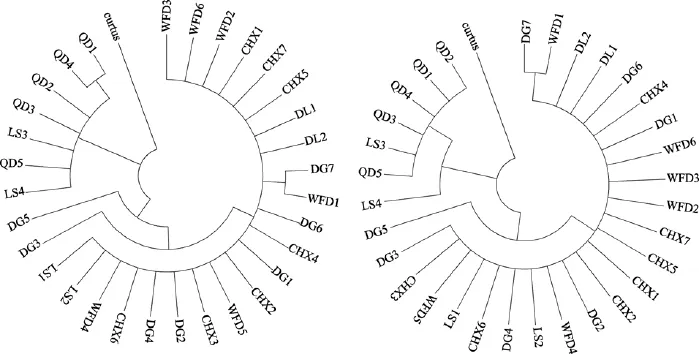
图2 大泷六线鱼部分线粒体DNA D-oop序列的NJ和MP分子系统树Fig. 2 Neighbor-joining and Maximum Parsimony tree based on partly mtDNA D-loop sequences
2.4 分子系统树的构建
选择短鲬(Parabembras curtus)的同源序列作为外群(登录号:GU139032.1), 基于Kimura双参数模型构建 NJ和 MP分子系统进化树, 节点支持率使用1 000次 bootstrap检验(图 2)。可以看出, 两种方法获得的进化树相似, 大泷六线鱼31尾个体聚为两大分支。其中东港、长海县、大连和瓦房店群体亲缘关系较近, 构成一个分支, 青岛群体构成另外一个分支, 旅顺群体在两大分支中各有分布。
3 讨论
一个物种遗传多样性的高低与其适应能力、生存能力和进化潜力密切相关。丰富的遗传多样性意味着比较高的适应生存潜力, 蕴藏着比较大的进化潜能以及比较丰富的育种和遗传改良的潜力, 而贫乏的遗传多样性则会给物种生存、进化及种质资源的保护和利用带来许多不利影响[29]。在本研究中, 6个群体31尾大泷六线鱼共检测到22种单倍型, 单倍型多样性指数为0.83333~1.00000。已有的资料表明,种群能够维持较高单倍型多样性的原因可能在于较大的种群数量、环境的不均一性或者具有适应种群快速增长的生活特性[30]。大泷六线鱼是地方性的鱼类, 资源量不大[31], 间接说明中国的近海多岩礁海区比较适合其生长繁殖, 这可能是其维持较高遗传多样性的基础。本研究发现, 与旅顺群体的核苷酸多样性指数(0.28938)相比, 东港群体(0.01988)、青岛群体(0.01612)、瓦房店群体(0.01001)、大连群体(0.00733)和长海县群体(0.00628)的核苷酸多样性均低。Grant等[32]指出单倍型多样性值高与核苷酸多样性值低的原因可能是由于种群是由一个较小的有效群体快速扩张而来的, 虽然通过变异积累了单倍型的多态性, 但却还未能积累核苷酸序列的多样化,这些群体的许多种类可能起源于上新世或更新世早期。此现象在鲸鲨[33]、尖吻鲭鲨、红拟石首鱼与西大西洋小沙丁鱼[32]中也存在。Fu[34]认为Fs中性检验显著偏离中性, 负的Fs值和差异显著的P值表明种群可能在历史上有扩张的迹象。在本实验中, 1000次模拟取样的情况下, 青岛、东港、瓦房店和长海县群体的Fs值均为负值(P<0.05), 表明这4个群体可能曾经历过种群扩张的历史, 而旅顺与大连群体的Fs值均为正值, 说明这两个群体没有经历过种群扩张的历史, 种群大小稳定。
Fst是用来测量群体之间遗传分化的重要指标,Fst接近于 0时, 说明群体间没有发生遗传分化, 在 0到1的范围内,Fst值越大, 两种群的分化程度越高[35]。本实验中旅顺和青岛群体与其他群体间Fst值较大(0.30275~0.97565), 已达高等分化水平, 其他四个群体之间没有出现遗传分化。另外, 群体间的相对遗传距离能反映群体间的亲缘关系, 本研究中旅顺和青岛群体与其他群体间的遗传距离较大(0.33209~0.66972), 其他 4个群体间的遗传距离均较小(0.00739 ~ 0.01519)。AMOVA 分析结果显示, 群体间遗传变异占总遗传变异较大的比例(73.98% ), 说明大泷六线鱼群体间差异较大, 遗传变异主要发生在群体间。构建的NJ和MP分子系统树显示, 青岛群体和旅顺群体的一部分个体组成一个分支, 东港、长海县、大连、瓦房店和旅顺群体的另一部分个体为另一大分支。单倍型在群体中的分布也显示, 除青岛群体外, 其他5个群体均有单倍型互相交叉。本实验结果表明, 可以将这6个群体划分为两大区域, 基本符合其所在的地理区域, 即青岛群体属于山东地区,东港、长海县、大连、瓦房店群体属于辽宁地区。由于mtDNA属于母系遗传, 由此可推断这6个群体的31尾个体可能来源于两个不同的母系祖先。但是作者看到旅顺群体的一部分与青岛群体的亲缘关系极近, 分析可能由于引种问题或者遗传漂变使两群体间发生了基因交流。
通过研究作者发现, 高比例的遗传变异分布在地区之间, 表明大泷六线鱼群体间具有显著的遗传分化。山东地区和辽宁地区之间没有共享的单倍型,虽然不排除有少量共享单倍型未被检出的可能性(尤其是在青岛群体和旅顺群体之间), 但我们仍然建议按照不同的地理区域来保护大泷六线鱼种群, 避免不同区域的种群之间发生基因交流, 因为基因交流在某种程度上会阻碍群体间的遗传分化[36]。本研究结果显示, 与其他鱼类的相比, 如细鳞鲑(0.0005)[24]、大口黑鲈(0.0028)[25]、沙鳅( 0.00365)[26]、银鲳(0.0007)[27]和裂腹鱼(0.00324)[28], 大泷六线鱼群体具有较高的核苷酸多样性(旅顺、东港、青岛、瓦房店、大连、长海县分别为: 0.28938、0.01988、0.01612、0.01001、0.00733、0.00628), 遗传多样性较为丰富。近年来, 随着近海经济的发展, 填海造地等严重破坏了大泷六线鱼的栖息场所, 而滥采滥捕等又加大了对该鱼种群的选择压力, 致使出现捕捞个体小型化、性早熟等种质衰退的现象。大泷六线鱼的怀卵量很少, 繁殖力低, 故很难形成大群体, 资源量也得不到大量补充,一旦资源被破坏, 很难恢复, 也可能会遭受灭顶之灾[31]。从长远角度看, 过度捕捞势必破坏其遗传多样性和种质资源的稳定性, 不能因为目前相对丰富的遗传多样性水平而忽视对大泷六线鱼资源必要的保护与管理。今后应针对中国范围内不同群体大泷六线鱼资源的遗传多样性展开更为详细的研究工作,后继的相关研究, 应尝试结合其他mtDNA基因信息以及其他分子标记方法进一步比较研究大泷六线鱼群体内及群体间遗传多样性的变动规律。以期为今后定期的遗传多样性检测奠定基础, 并根据其遗传多样性的变化趋势采取相应的保护措施, 科学的保护野生大泷六线鱼的种质资源。
[1]刘蝉馨, 秦克静. 辽宁动物志(鱼类)[M]. 沈阳: 辽宁科学技术出版社, 1987: 393-396.
[2]成庆泰, 郑宝珊. 中国鱼类系统检索[M]. 北京: 科学出版社, 1987.
[3]辽宁省海洋局. 辽宁省海岛资源综合调查研究报告[M].北京: 海洋出版社, 1996: 357.
[4]唐启升, 叶懋中. 山东近海渔业资源开发与保护[M].北京: 农业出版社, 1990: 80-102.
[5]Osamu Fukuhara, Toru Fushimi. Development and early live history of the greenlingsHexagrammos otakii(spices:Hexagrammidae) reared in the laboratory[J].Bulletin of the Japanese Society of Scientific Fishries,1983, 49(12): 1843-1848.
[6]吴立新, 秦克静, 姜志强, 等. 大泷六线鱼(Hexagrammos otakii)人工育苗初步试验[J]. 海洋科学,1996, 20(4): 32-34.
[7]于鸿仙, 庄虔增, 徐春华, 等. 六线鱼人工鱼苗技术研究[J]. 齐鲁渔业, 1998, 15(5): 21-24.
[8]庄虔增, 于鸿仙, 刘岗, 等. 六线鱼苗种生产技术的研究[J]. 中国水产科学, 1999, 6(1): 103-106.
[9]康斌, 武云飞. 大泷六线鱼的营养成分分析[J]. 海洋科学, 1999, 23(6): 23-25.
[10]Sang-Min L, Sung-Hwoan C, Kyoung-Duck K.Effect of dietary protein and energy levels on growth and body composition of juvenile flounderParalichthys olivaceus[J]. World Aquae Soc, 2000, 31(3): 306-315.
[11]王书磊, 姜志强, 苗治欧. 大连海区大泷六线鱼生物学指标的季节变化[J]. 水产科学, 2005, 24(5): 1-3.
[12]刘奇, 王亮. 北黄海大泷六线鱼主要生物学特征比较研究[J]. 中国海洋大学学报, 2009, 39(Sup.): 13-18.
[13]刘奇. 大泷六线鱼(Hexagrammos otakii)生物学特征与遗传多样性研究[D]. 山东: 青岛海洋大学硕士学位论文, 2010.
[14]Lu G, Li S. Advances in the study and application of fish mitochondrial DNA polymorphism[J]. Journal of Fisheries of China, 1998, 3: 94-103.
[15]吕国庆, 李思发. 鱼类线粒体 DNA多态研究和应用进展[J]. 中国水产科学, 1998, 5(3): 94-103.
[16]李春枝, 张邦杰, 李本旺, 等. 尖塘鳢属鱼类线粒体12SrRNA 基因序列分析[J]. 生态科学, 2006, 25(5):433-436.
[17]GattM H, FergusonM M, Liskauskas A P. Comparison of control region sequencing and fragment RFLP analysis for resolving mitochondrial DNA variation and phylogenetic relationships amongGreatLakesWal leyes[J]. Trans American Fish Soc, 2000, 129(6):1288-1299.
[18]Zardoya R, Meyer A. Mitochondrial evidence on the phylogenetic position of caecilians (Amphibia Gynnnophiona) [J]. Geneties, 2000, 2: 765-775.
[19]李思发, 吕国庆, 贝纳切兹. 长江中下游鲢鳙草青四大家鱼线粒体DNA多样性分析[J]. 动物学报, 1998,44(1): 82-93.
[20]郑冰蓉, 张亚平. 云南倒刺鲃mtDNAD-loop序列的遗传多样性研究[J]. 水利渔业, 2002, 22(3): 15-16.
[21]Ovenden J R, Lloyd J, Newman S J. Spatial genetic subdivision between northern Australian and southeast Asian populations ofPristipomoides multidens: a tropical marine reef fish species[J]. Fisheries Research,2002, 59: 57-69.
[22]胡文革, 段子渊, 王金富, 等. 新疆 3种雅罗鱼线粒体 DNA 控制区序列的差异和系统进化关系[J]. 遗传学报, 2004, 31(9): 970-975.
[23]Wang J P, Lin H D, Huang S. Phylogeography ofVaricorhinus barbatulus( Cyprinidae) in Taiwan based on nucleotide variation of mtDNA and allozymes[J].Molecular Phylogenetics and Evolution, 2004, 31:1143-1156.
[24]夏颖哲, 盛岩, 陈宜瑜. 利用线粒体 DNA 控制区序列分析细鳞鲑种群的遗传结构[J]. 生物多样性, 2006,14(1): 48-54.
[25]李胜杰, 白俊杰, 叶星, 等. 基于线粒体D-loop区探讨我国养殖大口黑鲈的分类地位和遗传变异[J]. 海洋渔业, 2008, 30(4): 292-296.
[26]刘红艳, 陈大庆, 刘绍平, 等. 长江上游中华沙鳅遗传多样性研究[J]. 淡水渔业, 2009, 39(3): 8-13.
[27]彭士明, 施兆鸿, 陈超, 等. 根据 mtDNA D-loop序列分析东海银鲳群体遗传多样性[R]. 海洋科学,2010, 34(2): 28-32.
[28]代应贵, 邹习俊, 肖海. 四川裂腹鱼乌江种群mtDNA控制区序列的遗传多样性分析[J]. 四川动物,2010, 29(4): 505-509.
[29]季维智, 宿兵. 遗传多样性研究的原理与方法[M].杭州: 浙江科学技术出版社, 1999.
[30]Nei M. Molecular Evolutionary Genetics [M]. New York: Columbia University Press, 1987.
[31]冯昭信, 韩华. 大泷六线鱼资源合理利用的研究[J].大连水产学院学报, 1998, 13(2): 24-28.
[32]Grant W S, Bowen B W. Population histories in deep evolutionary lineages of marine fishes: insights from sardines and anchovies and lessons for conservation [J].J Heredity, 1998, 89: 415-426.
[33]Ramirez-Macias D, Vazquez-Juarez R, Galvan-Magana F, et al. Variations of the mitochondrial control region sequence in whale sharks (Rhincodon typus) from the Gulf of California, Mexico [J]. Fisheries Research,2007, 84: 87-95.
[34]Fu Y X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection [J]. Genetics, 1997, 147: 915-925.
[35]Wright S. Evolution and the Genetics of Populations.Vol. 4[M]// Variability Within and Among Natural Populations. Chicago: University of Chicago Press,1978.
[36]黄原. 分子系统学-原理、方法及应用[M]. 北京: 中国农业出版社, 1998.
Genetic diversity analysis between different stocks ofHexagrammos otakiibased on analysis of mitochondrial DNA control region partial sequence
LI Ying1, WANG Wei1, MENG Fan-ping1, LI Zhan-dong2
(1. Key Laboratory of North Mariculture, Ministry of Agriculture, Dalian Ocean University Dalian 116023, China;2. Donggang Entry-Exit Inspection and Quarantine Bureau, Liaoning Province, Donggang 118300, China)
May,17,2011
Hexagrammos otakii; wide stocks; mitochondrial DNA control region partial sequence; genetic diversity
Nucleotide sequences of mitochondrial DNA control region partial sequence from six wide stocks ofHexagrammos otakiiwere amplified using PCR technique. Twenty-two haplotypes were identified from 31 individuals according to the determined sequences. The value of Fu′s Fs of neutrality tests was -0.14881 (P<0.01).AMOVA analysis demonstrated that theFstwas 0.7398 (P<0.01), and 73.98% variances occurred among populations and 26.02% variances occurred within populations. The NJ and MP molecular phylogenetic trees constructed by the distances among different individuals were similar. The phylogenetic trees were all divided into two branches.The Qingdao population and one part of the Lvshun population made up one branch, the rest populations were clustered into the other branch. The results suggested that the genetic diversity of theHexagrammos otakiipopulations was abundant, and there was a huge genetic differentiation among populations. It was also verified that the gene sequence of mitochondrial DNA control region could be used to analyze the genetic diversity ofHexagrammos otakiiwithin or among populations.
Q347
A
1000-3096(2012)08-0040-07
2011-05-17;
2011-07-04
博士启动基金资助项目(SYYJ200805); 省教育厅计划项目(2009A175)
李莹(1985-), 女, 吉林通化人, 硕士研究生, 主要从事鱼类群体遗传学的研究, E-mail: 000liying000@163.com; 王伟, 通信作者,博士, E-mail: wangwei@dlou.edu.cn
谭雪静)
