植物生长调节剂浸球对花芽分化期石蒜和换锦花鳞茎生化特性的影响
2011-12-31王呈伟郑玉红何树兰
李 莹,王呈伟,郑玉红,何树兰,彭 峰
〔江苏省·中国科学院植物研究所(南京中山植物园),江苏 南京 210014〕
石蒜〔Lycoris radiata(L’Hér.)Herb.〕和换锦花(L.sprengeri Comes ex Baker)均为石蒜属(Lycoris Herb.)多年生草本球根花卉。石蒜属植物原产中国和日本,在中国主要分布于长江流域及西南各省区,华东地区为其多样性分布中心[1-2]。石蒜属植物的花不仅形态多样而且色彩丰富,具有很高的观赏价值,现已成为切花、园林地被和盆栽的新型球根花卉。石蒜属植物的花期主要集中在夏、秋季节(8月份至9月份),且花期不长,在10月份前基本结束,这一特性极大的限制了石蒜属植物的推广应用。研究其花期生化特性,对石蒜属植物的花期调控具有重要的理论和应用意义。
石蒜属植物具有较高的观赏、药用价值以及花叶不相见的生物学特性,20世纪80年代以来,已有许多学者对该属的系统分类、种间关系和种质资源遗传多样性[3-6]、形态与生长发育规律[7-9]、生态地理分布和种群生物学[2,10]以及种间杂交和花粉生活力[11-12]等方面进行了广泛的研究,而关于石蒜属植物花期的生化特点只有零星的研究报道。梁素秋等[13]对台湾忽地笑〔L.aurea(L’Hér.)Herb.〕的花芽分化、发育特性和开花诱导因素的研究结果表明,忽地笑的花芽为预生型,早在2月停止抽叶后便开始花芽分化,由花芽原基分化到配子形成直至抽薹开花约需7~8个月的时间,但植株个体间的变异比较大。王磊等[14]观察了石蒜等3种石蒜属植物的开花习性,其中石蒜在日均温度约25℃时开始开花,而且在一定范围内,日均温度与小花开放的数量呈显著的负线性关系;开花高峰持续时间约为3 d;单朵花持续开放时间约为6 d。在开花期间,石蒜内源激素(GA、IAA、ZR和ABA)、多胺及核酸代谢等也均呈规律性的变化,对这一现象也有研究报道[15-16]。赵天荣等[17]则观察了石蒜等8种石蒜属植物花期的外部性状(如花蕾数、花直径、花被片的长度和宽度)等。但对于外源植物生长调节剂对石蒜属植物花芽分化期生化特性的影响目前尚未见研究报道。
作者以石蒜属2种展叶型植物——石蒜(秋出叶型)和换锦花(春出叶型)为实验材料,采用生产上应用广泛且快捷有效的3种植物生长调节剂(6-BA、GA3和乙烯利)对休眠鳞茎进行处理,测定花芽分化期间鳞茎中可溶性糖、可溶性蛋白质和核酸的含量,研究花芽分化期间石蒜和换锦花鳞茎生化特性的变化,了解其调控机制,以期为石蒜属植物的花期调控提供相关的理论依据。
1 材料和方法
1.1 材料
供试植物石蒜和换锦花均选自南京中山植物园苗圃地。于2009年11月底分别挖取大小一致、生长健壮的石蒜和换锦花鳞茎(挖球时尽量不伤根),鳞茎栽植前分别用质量浓度0(即对照CK,为蒸馏水)、60、120和180 mg·L-1的6-BA、GA3和乙烯利溶液浸球处理2 h,每处理30球,各重复3次。将处理后的鳞茎栽植于温室花盆中,选择排水良好且富含有机质的砂质壤土栽植,土面埋至鳞茎球颈部即可(若种植过浅,日照强烈时易发生日烧;若种植过深,高温高湿条件下易发生鳞茎腐烂)。管理过程中注意适时适量浇水、遮阳和通风。
1.2 方法
1.2.1 样品采集及前处理 石蒜属植物的花芽分化过程可分为花芽分化准备期(4月上旬)、分生组织膨大期(4月中下旬)、花原基形成期(5月上旬)、花被片分化期(5月下旬)、雄蕊分化期(6月上旬)、心皮分化期(6月中下旬)、雌蕊分化期(7月上旬)和花芽分化后期(7月中下旬)8个时期(待发表)。根据这个过程,在石蒜和换锦花的花芽分化期(4月至7月),每隔15天取样1次,每次均剥取对照及各处理组石蒜和换锦花鳞茎的中间部位鳞片(即花芽部位) 3~5片,每处理重复3球;于60℃干燥至恒质量,粉碎后用于可溶性糖、可溶性蛋白质和核酸含量的测定。
1.2.2 生化指标的测定 采用蒽酮比色法[18]195-197测定可溶性糖含量;采用考马斯亮蓝G-250染色法[18]184-185测定可溶性蛋白质含量;参照文献[16]的方法测定核酸含量。
实验采用随机区组设计,每样品重复测定9个鳞茎球(各处理重复3次,每一重复测定3个鳞茎球),每球重复测定3次,结果取平均值。
1.3 数据处理与分析
采用Excel 2007和SPSS 13.0软件对数据进行统计分析。
2 结果和分析
2.1 不同生长调节剂对花芽分化期内石蒜和换锦花鳞茎中可溶性糖含量的影响
2.1.1 6-BA的影响 60、120和180 mg·L-16-BA浸球处理对花芽分化期内石蒜和换锦花鳞茎中可溶性糖含量的影响见表1。由表1可以看出,在对照及各处理条件下,在花芽分化期(4月至7月),随着时间的推移,石蒜和换锦花鳞茎中可溶性糖含量总体上先逐渐增加,在6月25日(心皮分化期)或7月10日(雌蕊分化期)达到最高值,然后逐渐降低。同一处理条件下,石蒜鳞茎中可溶性糖含量总体上比同期换锦花高。
对照组石蒜鳞茎的可溶性糖含量在7月10日达到最高值,而经质量浓度60、120和180 mg·L-16-BA处理的石蒜鳞茎的可溶性糖含量均于6月25日达到最高值,分别比对照组高49.82%、53.70%和70.39%。相关性分析结果表明,在7月10日(雌蕊分化期)前,石蒜鳞茎中可溶性糖含量与6-BA质量浓度呈正相关关系。
在对照组和120 mg·L-16-BA处理组中,换锦花鳞茎中的可溶性糖含量在6月25日达到最高值;而60 mg·L-16-BA处理组换锦花鳞茎中的可溶性糖含量在7月25日(花芽分化后期)最高,比对照组高19.40%;在180 mg·L-16-BA处理组中,换锦花鳞茎中的可溶性糖含量在7月10日最高,较对照组高63.09%。相关性分析结果表明,在7月10日前,换锦花鳞茎中的可溶性糖含量与6-BA质量浓度呈负相关关系。
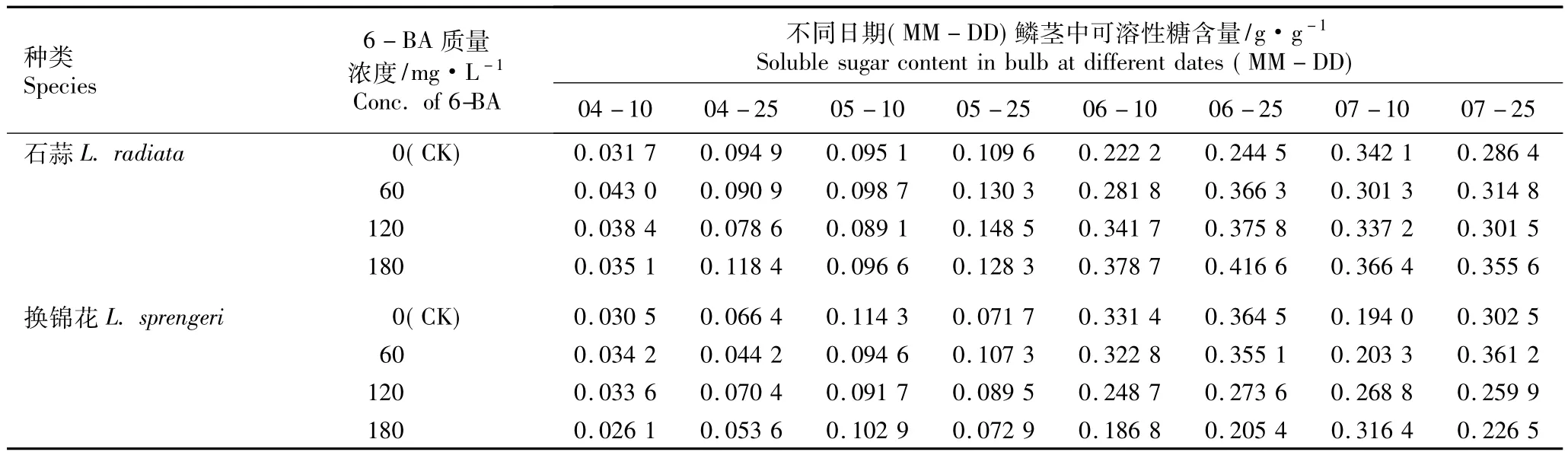
表1 不同质量浓度6-BA浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性糖含量的影响Table 1 Effect of different concentrations of 6-BA for soaking bulbs on soluble sugar content in bulbs of Lycoris radiata(L’Hér.)Herb.and L.sprengeri Comes ex Baker during flower bud differentiation period
2.1.2 GA3的影响 不同质量浓度GA3浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性糖含量的影响见表2。由表2可以看出,在花芽分化期(4月至7月),随时间的推移,对照组石蒜和换锦花鳞茎中可溶性糖含量呈先增加后降低的趋势,而在60、120和180 mg·L-1GA3处理组中,石蒜和换锦花鳞茎中可溶性糖含量的变化趋势不一致且与对照也有一定的差异。同一处理条件下,石蒜鳞茎中可溶性糖含量总体上比同期换锦花高。
在对照组和120 mg·L-1GA3处理组中,石蒜鳞茎的可溶性糖含量在7月10日(雌蕊分化期)达到最高值;在60和180 mg·L-1GA3处理组中,石蒜鳞茎的可溶性糖含量分别于7月25日(花芽分化后期)和6月25日(心皮分化期)达到最高值,分别为对照的1.22和1.50倍。相关性分析结果表明,在7月10日前,石蒜鳞茎中可溶性糖的含量与GA3的质量浓度呈正相关。
在对照组中,换锦花鳞茎中可溶性糖含量也在7月10日达到最高值;而在60、120和180 mg·L-1GA3处理组中,换锦花鳞茎中可溶性糖含量分别在6月25日、7月25日和7月25日达到最高值,分别为同期对照的76.34%、102.38%和85.62%。相关性分析结果表明,在7月10日前,换锦花鳞茎中的可溶性糖含量与GA3的质量浓度呈负相关。
在5月10日(花原基形成期)、6月10日(雄蕊分化期)和6月25日,60、120和180 mg·L-1GA3处理组中换锦花鳞茎中的可溶性糖含量显著低于对照,说明用GA3浸球处理在一定程度上抑制了换锦花的糖代谢过程;而用GA3浸球处理后石蒜鳞茎中可溶性糖含量的变化没有6-BA处理组显著,说明在花芽分化期间石蒜对6-BA处理的生化响应更明显。
2.1.3 乙烯利的影响 不同质量浓度乙烯利浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性糖含量的影响见表3。由表3可以看出,在花芽分化期内(4月至7月),随着时间的推移,对照组石蒜和换锦花鳞茎中的可溶性糖含量呈先增加后降低的趋势;在60、120和180 mg·L-1乙烯利处理组中,石蒜鳞茎中可溶性糖含量的变化趋势不一致,而换锦花鳞茎中可溶性糖含量总体上呈逐渐增加的趋势。同一处理条件下,同期石蒜鳞茎中的可溶性糖含量总体上高于换锦花,这一现象与6-BA和GA3处理组的结果相似。经乙烯利浸球后整个花芽分化期内石蒜鳞茎的可溶性糖含量总体上比同期对照高,而换锦花鳞茎的可溶性糖含量大多低于同期的对照。
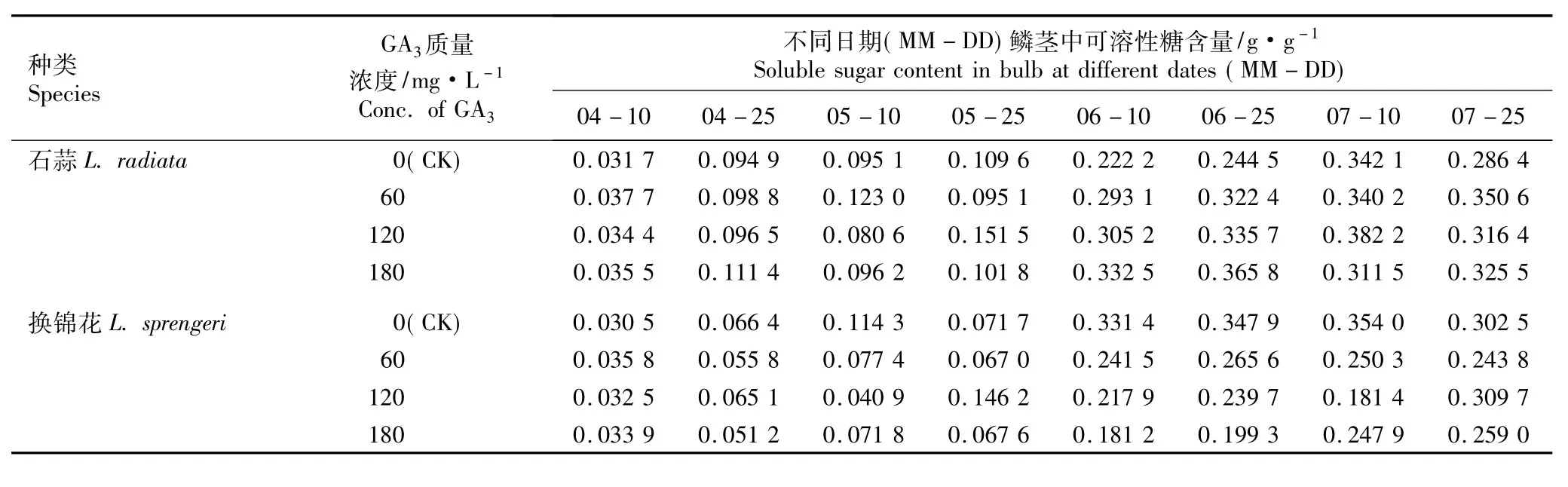
表2 不同质量浓度GA3浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性糖含量的影响Table 2 Effect of different concentrations of GA3 for soaking bulbs on soluble sugar content in bulbs of Lycoris radiata(L’Hér.)Herb.and L.sprengeri Comes ex Baker during flower bud differentiation period
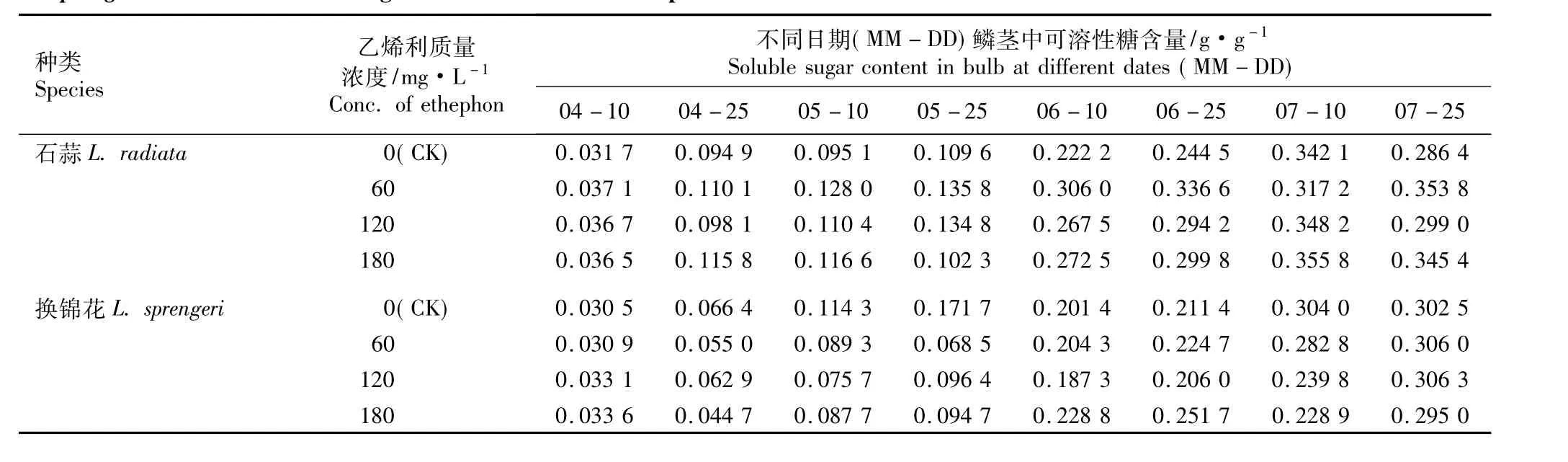
表3 不同质量浓度乙烯利浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性糖含量的影响Table 3 Effect of different concentrations of ethephon for soaking bulbs on soluble sugar content in bulbs of Lycoris radiata(L’Hér.)Herb.and L.sprengeri Comes ex Baker during flower bud differentiation period
在对照组中,石蒜和换锦花鳞茎中可溶性糖含量都在7月10日(雌蕊分化期)达到最高值。经120和180 mg·L-1乙烯利浸球处理后,石蒜鳞茎中可溶性糖含量也在7月10日达到最高值并略高于对照;而经60 mg·L-1乙烯利浸球处理后,石蒜鳞茎中可溶性糖含量在7月25日(花芽分化后期)达到最高值,比对照增加了23.53%。经60、120和180mg·L-1乙烯利浸球处理后,换锦花鳞茎中可溶性糖含量均在7月25日达到最高值,且与对照的差异均较小。
2.2 不同生长调节剂对花芽分化期内石蒜和换锦花鳞茎中可溶性蛋白质含量的影响
2.2.1 6-BA的影响 不同质量浓度6-BA浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性蛋白质含量的影响见表4。由表4可见,在花芽分化期内(4月至7月),随时间的推移,对照组石蒜和换锦花鳞茎中的可溶性蛋白质含量呈先增加后降低的趋势,而在60、120和180 mg·L-16-BA处理组中,石蒜和换锦花鳞茎中可溶性蛋白质含量的变化趋势有差异。
在对照组中,石蒜和换锦花鳞茎中可溶性蛋白质含量分别在7月10日(雌蕊分化期)和6月25日(心皮分化期)达到最高值。在60 mg·L-16-BA处理条件下,石蒜鳞茎中可溶性蛋白质含量在7月25日(花芽分化后期)最高,为对照的1.19倍;在120和180 mg·L-16-BA处理组中,石蒜鳞茎中可溶性蛋白质含量在6月25日最高,分别为对照的1.08和1.12倍。在60和120 mg·L-16-BA处理组中,换锦花鳞茎中可溶性蛋白质含量均在7月25日达到最高值并略高于对照;而在180 mg·L-16-BA处理组中,换锦花鳞茎中可溶性蛋白质含量在6月25日最高,且也略高于对照。
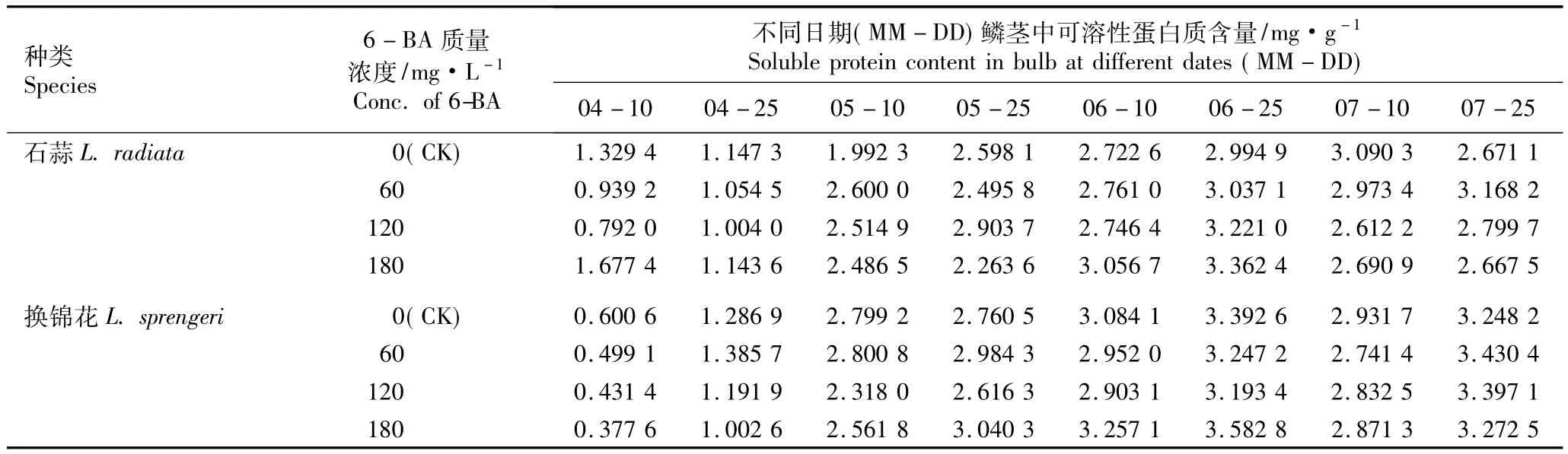
表4 不同质量浓度6-BA浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性蛋白质含量的影响Table 4 Effect of different concentrations of 6-BA for soaking bulbs on soluble protein content in bulbs of Lycoris radiata(L’Hér.)Herb.and L.sprengeri Comes ex Baker during flower bud differentiation period
在对照和各处理组中,在5月10日(花原基形成期)前石蒜和换锦花鳞茎中可溶性蛋白质含量均较低;自花原基形成后,可溶性蛋白质含量增加了约1倍,并在之后的花芽分化期有不同程度的增加。差异显著性分析结果表明,对照组和各处理组间石蒜和换锦花鳞茎中可溶性蛋白质含量的差异均未达到显著水平,说明60、120和180 mg·L-16-BA浸球处理对花芽分化期内石蒜和换锦花鳞茎可溶性蛋白质含量的影响不显著。
2.2.2 GA3的影响 不同质量浓度GA3浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性蛋白质含量的影响见表5。由表5可见,在花芽分化期内(4月至7月),随时间的推移,对照组及60、120和180 mg·L-1GA3处理组中石蒜和换锦花鳞茎中可溶性蛋白质含量总体上呈先增加后降低的趋势。
在对照组及60、120和180 mg·L-1GA3处理组中,石蒜和换锦花鳞茎中可溶性蛋白质的含量在5月10日(花原基形成期)前均较低;自花原基形成后,可溶性蛋白质含量都维持在较高的水平且有不同程度的增加,与6-BA处理的结果相似。
与对照组相比,经GA3浸球处理后石蒜和换锦花鳞茎中可溶性蛋白质含量变化不明显,这一现象与6-BA处理的结果不同,说明在花芽分化期内石蒜和换锦花鳞茎对GA3浸球处理的响应明显不如6-BA。在花芽分化期的不同阶段,用180 mg·L-1GA3浸球处理的石蒜和换锦花鳞茎中可溶性蛋白质含量大都低于或略高于60和120 mg·L-1GA3处理组,说明用较高质量浓度的GA3浸球处理对石蒜和换锦花鳞茎中可溶性蛋白质的合成没有促进作用。
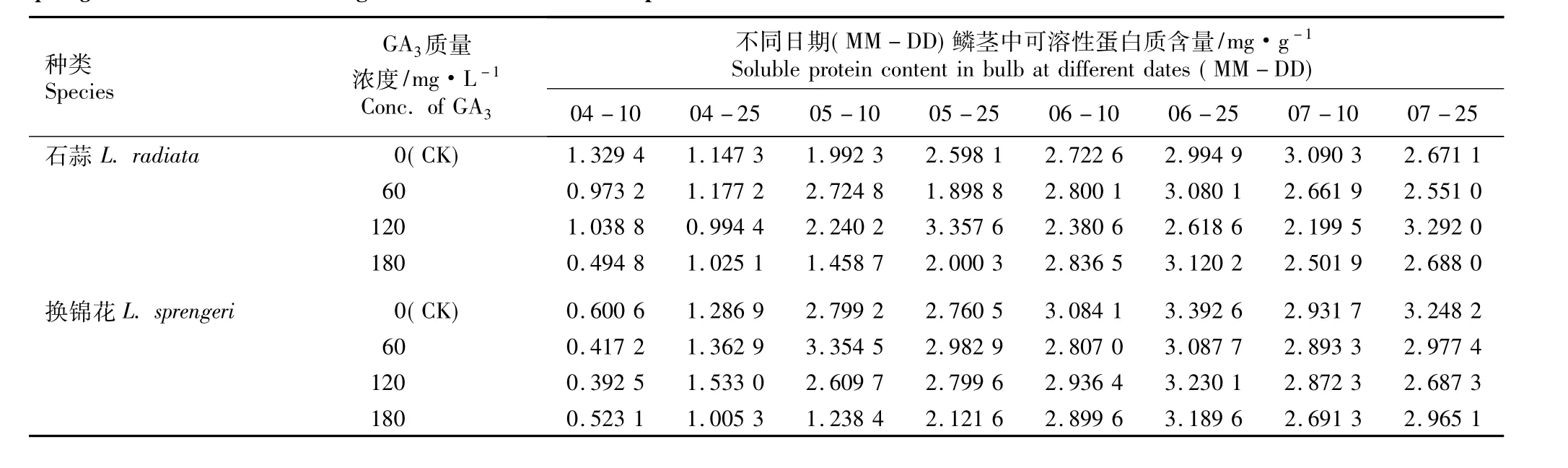
表5 不同质量浓度GA3浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性蛋白质含量的影响Table5 Effect of different concentrations of GA3 for soaking bulbs on soluble protein content in bulbs of Lycoris radiata(L’Hér.)Herb.and L.sprengeri Comes ex Baker during flower bud differentiation period
2.2.3 乙烯利的影响 不同质量浓度乙烯利浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性蛋白质含量的影响见表6。由表6可以看出,在花芽分化期内(4月至7月),随时间的推移,对照组及60、120和180 mg·L-1乙烯利处理组石蒜和换锦花鳞茎中的可溶性蛋白质含量总体上呈先增加后降低的趋势。
在对照组及60、120和180 mg·L-1乙烯利处理组中,石蒜和换锦花鳞茎中可溶性蛋白质的含量均在5月10日(花原基形成期)前较低;自花原基形成后,石蒜和换锦花鳞茎中可溶性蛋白质含量都维持在较高的水平且有不同程度的增加,这一现象与6-BA和GA3处理的结果均相似。
在对照组中,石蒜鳞茎中可溶性蛋白质含量在7月10日(雌蕊分化期)达到最高值,而用不同质量浓度乙烯利浸球处理后石蒜鳞茎中可溶性蛋白质含量提前在6月25日(心皮分化期)达到最高值。在对照组及120和180 mg·L-1乙烯利处理组中,换锦花鳞茎中的可溶性蛋白质含量均在6月25日达到最高值;而在60 mg·L-1乙烯利处理组中,换锦花鳞茎中可溶性蛋白质含量却推后至7月25日(花芽分化后期)达到最高值。从这一现象看,乙烯利浸球处理对石蒜鳞茎的影响较换锦花显著。
2.3 不同生长调节剂对花芽分化期内石蒜和换锦花鳞茎中核酸含量的影响
2.3.1 6-BA的影响 不同质量浓度6-BA浸球对花芽分化期内石蒜和换锦花鳞茎中核酸含量的影响见表7。由表7可以看出,在花芽分化期内(4月至7月),在对照组及60、120和180 mg·L-16-BA处理组中,石蒜和换锦花鳞茎中的核酸含量在7月10日(雌蕊分化期)前都维持在较低的水平,雌蕊分化期迅速增加并达到峰值,之后又急剧下降;此时(7月10日),对照组及60、120和180 mg·L-16-BA处理组石蒜鳞茎中的核酸含量分别为4月10日(花芽准备期)的26.98、26.08、60.95和50.39倍,换锦花中的核酸含量分别为4月10日的10.03、16.92、6.82和10.57倍,说明雌蕊分化期是石蒜和换锦花鳞茎细胞分裂最旺盛的时期。
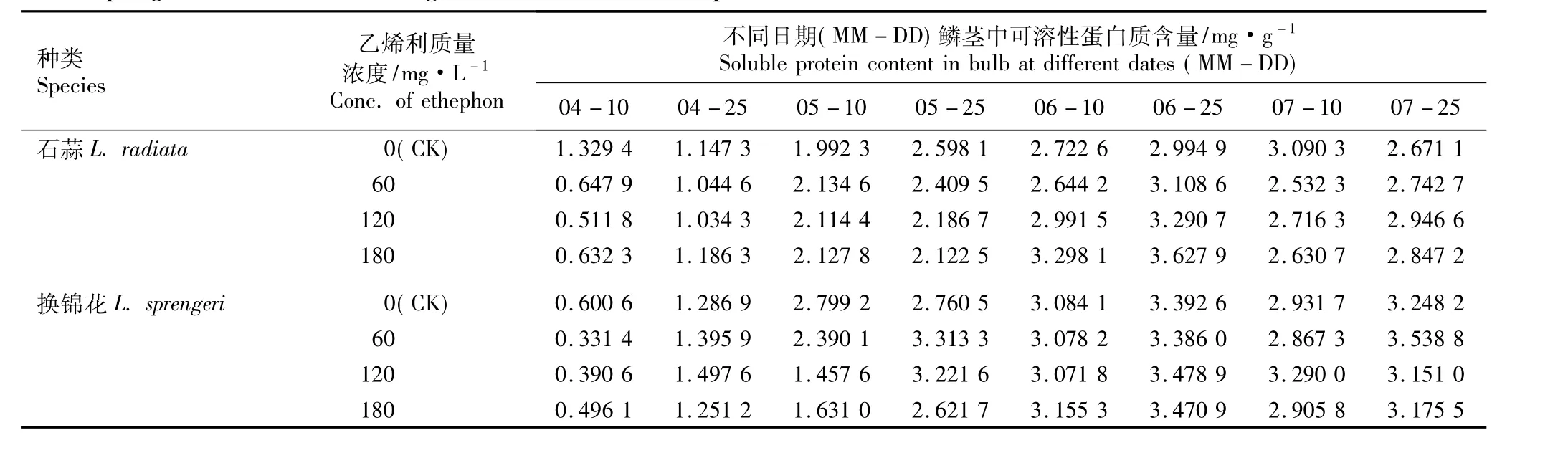
表6 不同质量浓度乙烯利浸球对花芽分化期内石蒜和换锦花鳞茎中可溶性蛋白质含量的影响Table 6 Effect of different concentrations of ethephon for soaking bulbs on soluble protein content in bulbs of Lycoris radiata(L’Hér.)Herb.and L.sprengeri Comes ex Baker during flower bud differentiation period

表7 不同质量浓度6-BA浸球对花芽分化期内石蒜和换锦花鳞茎中核酸含量的影响Table 7 Effect of different concentrations of 6-BA for soaking bulbs on nucleic acid content in bulbs of Lycoris radiata(L’Hér.)Herb.and L.sprengeri Comes ex Baker during flower bud differentiation period
2.3.2 GA3的影响 不同质量浓度GA3浸球对花芽分化期内石蒜和换锦花鳞茎中核酸含量的影响见表8。由表8可见,在花芽分化期内(4月至7月),在对照组及60、120和180 mg·L-16-BA处理组中,石蒜和换锦花鳞茎中的核酸含量在7月10日(雌蕊分化期)前均较低,雌蕊分化期迅速增加并达到峰值,之后又迅速降低,这一现象与6-BA处理的结果相似。
在雌蕊分化期,随GA3质量浓度的提高,石蒜鳞茎中核酸含量随之增加,经180 mg·L-1GA3浸球处理的石蒜鳞茎中核酸含量比同期对照高56.67%,说明用高质量浓度的GA3浸球处理促进了该时期石蒜鳞茎细胞分裂的速度;而随GA3质量浓度的提高,雌蕊分化期换锦花鳞茎中的核酸含量较对照有不同程度降低,但降低幅度不大,在一定程度上说明高质量浓度GA3浸球处理对该时期换锦花鳞茎的细胞分裂有轻微的抑制作用。
2.3.3 乙烯利的影响 不同质量浓度乙烯利浸球对花芽分化期内石蒜和换锦花鳞茎中核酸含量的影响见表9。由表9可以看出,在花芽分化期内(4月至7月),在对照组及60、120和180 mg·L-16-BA处理组中,石蒜和换锦花鳞茎中的核酸含量在7月10日(雌蕊分化期)前也都维持在较低水平,在雌蕊分化期迅速升高并达到峰值,之后又迅速降低。在雌蕊分化期,各处理组间石蒜和换锦花鳞茎中核酸含量的差异不显著;但此时期各处理组石蒜鳞茎中的核酸含量均明显高于对照,60、120和180 mg·L-1处理组石蒜鳞茎中的核酸含量分别比对照高31.59%、28.57%和30.54%。而同在雌蕊分化期,各处理组换锦花鳞茎中核酸含量仅略高于对照,说明乙烯利浸球处理对雌蕊分化期石蒜鳞茎核酸代谢的影响效应强于换锦花。
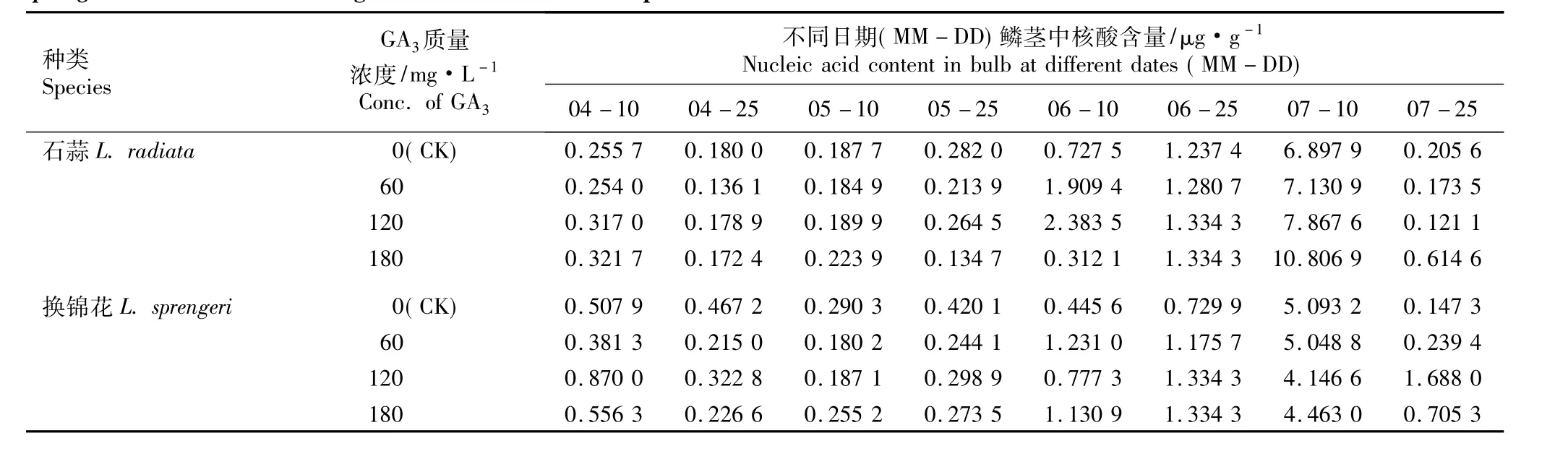
表8 不同质量浓度GA3浸球对花芽分化期内石蒜和换锦花鳞茎中核酸含量的影响Table 8 Effect of different concentrations of GA3 for soaking bulbs on nucleic acid content in bulbs of Lycoris radiata(L’Hér.)Herb.and L.sprengeri Comes ex Baker during flower bud differentiation period

表9 不同质量浓度乙烯利浸球对花芽分化期内石蒜和换锦花鳞茎中核酸含量的影响Table 9 Effect of different concentrations of ethephon for soaking bulbs on nucleic acid content in bulbs of Lycoris radiata(L’Hér.)Herb.and L.sprengeri Comes ex Baker during flower bud differentiation period
3 讨 论
3.1 6-BA、GA3和乙烯利浸球处理对花芽分化期内石蒜和换锦花鳞茎中生化指标的影响效应分析
经60、120和180 mg·L-1的6-BA、GA3和乙烯利浸球处理后,在花芽分化期内(4月至7月),随着时间的推移,石蒜和换锦花鳞茎中可溶性糖含量逐渐升高,至心皮分化期(6月25日)可溶性糖含量或达到峰值后开始下降或继续升高,这一现象与经温度诱导的蝴蝶兰(Phalaenopsis aphrodita Rchb.F.)花芽分化时期叶片可溶性糖含量的变化趋势有相似之处[19]。李哖等[20]在研究蝴蝶兰花发育进程中碳水化合物含量的变化时指出:在花芽发育时期植物体内的可溶性糖含量发生变化是因为此时的生长中心为花芽。糖类代谢是植物体有机物代谢的中心,它为各种有机物代谢提供碳骨架。石蒜和换锦花花芽分化初期可溶性糖含量的增加为花芽的分化和正常生长储备了物质和能量;花芽分化后期可溶性糖含量有所下降,可能是因为可溶性糖大多被分配到花芽中,为石蒜和换锦花的抽薹开花奠定基础。本研究结果显示,用60、120和180 mg·L-1的6-BA、GA3和乙烯利浸球处理后,花芽分化期内石蒜和换锦花鳞茎中可溶性糖含量总体上与对照间差异显著,间接说明外源植物生长调节剂(6-BA、GA3和乙烯利)浸球处理确实对石蒜和换锦花花芽分化过程有一定的影响。
用60、120和180 mg·L-16-BA、GA3和乙烯利浸球处理后,花芽分化期内石蒜和换锦花鳞茎中可溶性蛋白质和核酸含量的变化趋势与可溶性糖含量相似,但与对照间的差异大多不显著。石蒜和换锦花鳞茎中可溶性蛋白质含量在进入花原基形成期(5月10日)后始终维持在较高水平,与此时旺盛的新陈代谢相适应。因为可溶性蛋白质主要包含了催化新陈代谢的各种酶,其含量的增加也保证了各种代谢过程的顺利进行和花芽分化的进程。在花芽分化期的前期,石蒜和换锦花鳞茎中核酸含量较低,而在雌蕊分化期(7月10日)迅速增加,之后又急剧下降,表明此时鳞茎细胞分裂旺盛,是花芽形态建成的关键时期,这与王磊等[16]的研究结果一致。
王磊等[16]的研究结果表明:石蒜花芽分化期间内源激素含量的变化明显,其中IAA(吲哚乙酸)和GA在雄蕊形成期含量最高,ZR(玉米素)在雌蕊形成期含量最高。内源激素含量的变化影响着石蒜的新陈代谢,进而调控花芽分化的进程。本研究结果表明,外源植物生长调节剂也在一定程度上影响了花芽分化期石蒜和换锦花鳞茎的生理代谢过程。这一结果为采用外源植物生长调节剂调控石蒜属植物的花芽分化进程及花期奠定了理论基础。
3.2 花芽分化期内石蒜和换锦花鳞茎对6-BA、GA3和乙烯利浸球处理的生化响应分析
本研究结果显示,秋出叶型的石蒜和春出叶型的换锦花对6-BA、GA3和乙烯利浸球处理的响应不同。与对照相比,3种植物生长调节剂处理总体上导致石蒜鳞茎中可溶性糖含量有不同程度的提高;但3种植物生长调节剂处理对换锦花鳞茎中可溶性糖含量的影响不明显,与对照相比没有明显的变化趋势。说明6-BA、GA3和乙烯利浸球处理对石蒜鳞茎的花芽分化进程有促进作用,而对换锦花鳞茎的花芽分化进程的影响效应不明显。而且经不同质量浓度6-BA、GA3和乙烯利浸球处理后,总体上石蒜鳞茎中可溶性糖含量高于换锦花,可溶性蛋白质含量低于换锦花,而核酸含量与换锦花鳞茎的差异不明显。形成这一现象的原因可归结为二方面:一方面石蒜和换锦花二者本身遗传及代谢特性有差异;另一方面换锦花和石蒜的物候期不同,前者的花期略早于后者,而本研究中二者同时取样也会造成实验结果的差异。
[1]中国科学院中国植物志编辑委员会.中国植物志:第十六卷第一分册[M].北京:科学出版社,1985:18-27.
[2]王仁师.关于石蒜属(Lycoris)的生态地理[J].西南林学院学报,1990,10(1):41-48.
[3]张 露,蔡友铭,诸葛强,等.石蒜属种间亲缘关系RAPD分析[J].遗传学报,2002,29(10):915-921.
[4]吴 玲,卢毅军,史树德,等.中国石蒜属种间亲缘关系ITS序列分析[J].亚热带植物科学,2007,36(1):31-35.
[5]陈 妍,高燕会,廖望仪,等.乳白石蒜rDNA-ITS序列分析及种内系统发育研究[J].植物资源与环境学报,2009,18(3): 25-31.
[6]邓传良,周 坚,卢龙斗,等.长筒石蒜种质资源的RAPD及ISSR研究[J].云南植物研究,2006,28(3):300-304.
[7]林定勇,李 哖.石蒜属球根花卉之分类、形态、生长与开花[J].中国园艺,1993,39(2):67-72.
[8]王 磊,赵九州,汤庚国.几种植物生长调节剂对石蒜生理生化特性的影响[J].南京林业大学学报:自然科学版,2004,28 (4):39-42.
[9]李爱荣,周 坚.中国石蒜叶片的生长周期及其发育过程的研究[J].植物学通报,2005,22(6):680-686.
[10]秦卫华,周守标,汪恒英,等.石蒜属植物的研究进展[J].安徽师范大学学报:自然科学版,2003,26(4):385-390.
[11]徐炳声,林巾箴,俞志洲,等.换锦花和中国石蒜的种间杂交[J].园艺学报,1986,13(4):283-284.
[12]林巾箴,俞志洲,徐炳声,等.红蓝石蒜和中国石蒜的种间杂交[J].广西植物,1988,8(2):165-168.
[13]梁素秋,许圳塗.金花石蒜(Lycoris aurea Herb.)预生花芽分化与发育特性及其诱导因素探讨[J].中国园艺,1992,38(3): 139-148.
[14]王 磊,汤庚国,赵九州.3种石蒜属植物开花特性的研究[J].江苏农业科学,2008(1):112-115.
[15]金雅琴,黄雪芳,李冬林,等.石蒜花期前后鳞茎内源多胺含量的动态变化[J].南京林业大学学报:自然科学版,2007,31 (5):117-120.
[16]王 磊,汤庚国,刘 彤.石蒜花芽分化期内源激素和核酸含量的变化[J].南京林业大学学报:自然科学版,2008,32(4): 67-70.
[17]赵天荣,施永泰,蔡建岗,等.石蒜属植物生长发育习性的研究[J].黑龙江农业科学,2008(5):89-91.
[18]李合生.植物生理生化实验原理和技术[M].1版.北京:高等教育出版社,2000.
[19]刘晓荣,王碧青,朱根发,等.高山低温诱导蝴蝶兰花芽分化过程中的生理变化[J].中国农学通报,2006,22(4):310-313.
[20]李 哖,李嘉慧.蝴蝶兰花芽诱引和花序发育时之碳水化合物变化[J].中国园艺,1996,42(3):262-275.
