两种贵州地方鸡线粒体DNA控制区全序列分析
2011-11-18王文涛陈彬傅筑荫
王文涛 陈彬 傅筑荫

摘要:采用PCR和直接测序的方法测定兴义矮脚鸡和长顺绿壳蛋鸡线粒体DNA控制区全序列,分析比较两者mtDNA D-loop序列3个区的变异。结果表明,兴义矮脚鸡和长顺绿壳蛋鸡线粒体DNA控制区全序列长分别为1 231和1 230 bp,其中兴义矮脚鸡mtDNA控制区的碱基摩尔分数分别为A 26.92%、T 33.41%、G 13.39%、C 26.29%,长顺绿壳蛋鸡的分别是A 27.13%、T 33.58%、G 13.20%、C 26.09%。t检验显示两种贵州地方鸡种间线粒体DNA控制区碱基A与C的含量差异显著,说明A与C之间的颠换较多。兴义矮脚鸡和长顺绿壳蛋鸡的遗传距离是0.038 6,根据分子钟计算,二者大约在193万年前分歧进化。
关键词:线粒体DNA控制区;兴义矮脚鸡;长顺绿壳蛋鸡;遗传变异
中图分类号:Q343.3文献标识码:A文章编号:0439-8114(2011)15-3114-02
Analysis on the mtDNA Control Region of Two Kinds of Guizhou Native Chicken
WANG Wen-taoa,CHEN Bina,FU Zhu-yinb
(a. Key Laboratory of Animal Genetic, Breeding and Reproduction in the Plateau Mountainous Region, Ministry of Education;
b. College of Animal Sciences,Guizhou University,Guiyang 550025,China)
Abstract: PCR and direct sequencing methods were used to analyze the entire mtDNA D-loop in Xingyi Bantam Chicken and Chang shun Blue-shelled Egg Chicken. The D-loop of Xingyi Bantam and Changshun Blue-shelled Egg Chicken was
1 231 and 1 230 nucleotides long, respectively. The nucleotide molar fraction of Xingyi Bantam was A 26.92%, T 33.41%, G 13.39%, and C 26.29% , while the compsition of Changshun Blue-shelled Egg Chicken was A 27.13%, T 33.58%, G 13.20% and C 26.09%. The t-test results showed that the difference of the nucleotide A and C content in the mtDNA control region of the two chicken breeds was significant. The genetic distance between the two breeds was 0.038 6. Calculated by molecular clock, the two breeds diverged and evolved about 1.93 million years ago.
Key words: mtDNA D-loop; Xingyi Bantam chicken; Changshun Blue-shelled Egg Chicken; genetic variation
线粒体DNA控制区是非编码基因区,在进化过程中受环境的选择压力小,表现出进化速率快和母系遗传等特点,在鸟类的分类研究中常用作种系发生的分子标记[1]。
在漫长的家鸡养殖历史中,贵州形成了丰富的地方鸡品种资源。兴义矮脚鸡和长顺绿壳蛋鸡是两种优良的贵州地方鸡种,近年来由于生存环境遭到破坏、引进外地品种杂交改良等人为因素以及某些自然因素的影响,二者的开发利用受到限制[2,3]。对二者的亲缘关系和起源分化的研究不仅可以弄清它们的亲缘关系状况,为进一步选育和杂交配套提供理论基础,还可探讨人工选择对同一个地理群体遗传分化的影响。本研究通过比较兴义矮脚鸡和长顺绿壳蛋鸡两类群间的mtDNA序列变异,分析了二者的亲缘关系。
1材料与方法
1.1试验材料
试验用的兴义矮脚鸡、长顺绿壳蛋鸡各为15个样本。翅静脉采血0.5~1.0 mL,肝素钠抗凝,4 ℃保存备用。采用TIANamp Blood DNA Kit试剂盒提取血样中DNA,-20 ℃保存备用。兴义矮脚鸡、长顺绿壳蛋鸡均由贵州大学科研鸡场提供。
1.2PCR反应及测序
PCR扩增引物为PHDL(5′-AGGACTACGGCTT
GAAAAGC-3′)、PHlH(5′-TTATGTGCTTGACCGAG
GAACCAG-3′)和L400(5′-ATTTATTGATCGTCCA
CCTCACG-3′)、PHDH(5′-CATCTTGGCATCTTCAG
TGCC-3′)(引物名称末尾字母L、H分别代表轻链和重链)[4]。25 μL PCR反应液中含DNA样品2 μL,上下游引物各0.8 μL,2×Taq PCR MasterMix试剂12.5 μL,dd H2O 8.9 μL。PCR循环参数(30循环)为94 ℃预变性3 min,94 ℃变性30 s,61~63 ℃复性30 s,72 ℃延伸1 min,最后72 ℃延伸4 min。用2%琼脂糖凝胶检测PCR产物。扩增产物经TIANgel Midi Purification Kit试剂盒回收纯化后由北京诺赛基因组研究中心完成测序。
1.3序列分析
利用Vector NTI Suite 8对原始DNA序列进行对位排列,并经人工仔细核查后,利用DnaSP 4.0[5]提取变异位点。利用MEGA 4.0[6]统计碱基组成,并计算基于Kimura双参数模型的遗传距离。以原鸡为外群(GenBank登录号:AB007720),构建NJ分子系统发生树。
2结果
2.1mtDNA D-loop区序列变异
用Vector NTI Suite 8对原始DNA序列进行对位排列和剪切对齐后,得到兴义矮脚鸡、长顺绿壳蛋鸡线粒体DNA控制区序列全长分别为1 231 bp、1 230 bp。与兴义矮脚鸡相比,长顺绿壳蛋鸡mtDNA D-loop区全序列在第446位点缺失。两者之间共发现 23个变异位点,包括11个转换和12个颠换,11个转换分别是5次T-C间转换,6次A-G间转换;颠换中,A-T间2次,G-T间1次,A-C 间8次,G-C间1次。
兴义矮脚鸡和长顺绿壳蛋鸡mtDNA D-loop区Ⅰ区有13个变异位点,占总变异数的56.5%,序列变异率为4.11%;Ⅱ区有 2个变异位点,序列变异率为0.43%;Ⅲ区有8个变异位点,序列变异率为1.79%。
2.2mtDNA D-loop区序列碱基组成
兴义矮脚鸡、长顺绿壳蛋鸡mtDNA控制区的碱基含量见表1。由表1可见,两种贵州地方鸡A、T、G、C 4种碱基的平均摩尔分数分别为27.02%、33.49%、13.29%、26.19%,A+T含量高于G+C,这符合鸡mtDNA D-loop区是A+T富含区、而T含量最高的特点,表现出mtDNA D-loop区碱基组成的偏倚性。t检验显示两种贵州地方鸡种间线粒体DNA控制区碱基A与C的含量差异显著(P<0.05),说明A与C之间的颠换较多。
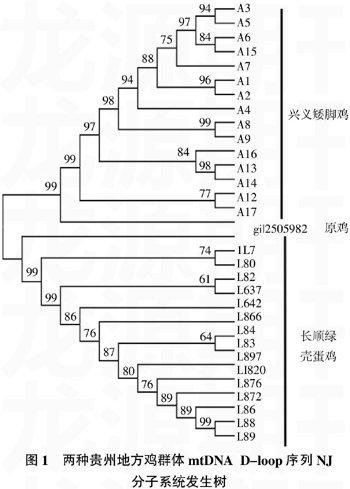
2.3系统发生树
基于Kimura双参数模型计算兴义矮脚鸡与长顺绿壳蛋鸡的遗传距离为0.038 6,两种贵州地方鸡种与原鸡的遗传距离为0.210 6。
依据测定的序列,以原鸡为外群,构建兴义矮脚鸡、长顺绿壳蛋鸡的NJ分子系统发生树(图1)。兴义矮脚鸡、长顺绿壳蛋鸡在系统树上各聚成一支,并且各支的置信度很高,特别是种间的置信度达到99%。
3讨论
鸟类mtDNA D-loop区的进化速率为每百万年2%[7,8],以此计算兴义矮脚鸡与长顺绿壳蛋鸡的分歧进化时间约为193万年。但碱基转换易饱和,因此可用颠换估算二者间的分歧时间。由于没有贵州地方鸡种进化时间参考,我们以石鸡和大石鸡为参照。两种鸡mtDNA D-loop区全序列的颠换数有10个,其分歧时间约为190万年[9],碱基颠换速率为5.26个/百万年。兴义矮脚鸡和长顺绿壳蛋鸡间的碱基颠换数为12个,由此推算二者的分歧时间大约为220万年前,属于更新世第二次寒冷期[10]。贵州地处我国西南地区,可能是更新世第二次寒冷期严重的山脊冰川作用将二者隔离在不同的低地,最终分歧进化成不同的地方鸡种。
参考文献:
[1] 刘益平,曾凡同,张思婉,等.几种山地乌骨鸡线粒体DNA序列多态性分析[A]. 家禽研究最新进展——第十一次全国家禽学术讨论会论文集 [C].北京:中国畜牧兽医学会,2003.198-201.
[2] 潘周雄,张福平,曹娟,等.贵州长顺绿壳蛋鸡简介[J].中国家禽,2008,30(4):53-54.
[3] 王文涛,陈彬,傅筑荫,等.贵州地方鸡种资源的现状、存在问题及对策[J].贵州农业科学,2010,38(8):155-157.
[4] RANDI E, LUCCHINI V.0rganization and evolution of the mitochondrial DNA control region in the avian genus Alectoris[J]. J Mol Evol,1998,47(4):449-462.
[5] ROZAS J, ROZAS R. DnaSP Version 3:an integrated program for molecular population genetics and molecular evolution analysis[J]. Bioinformatics,1999,15(2):174-175.
[6] TAMURA K, DUDLEY J, NEI M, et al. MEGA4:Molecular evolutionary genetics analysis(MEGA) software version 4.0 [J].Mol Boil Evol,2007,24(8):l596-1599.
[7] KLBINSKL D O F,GALLAGHER C,EBYNON C M,et al. Mitochondrial DNA inheritance[J].Nature, 1994,368(6474):817-818.
[8] 刘益平,朱庆,曾凡同,等.原鸡线粒体DNA部分序列多态性分析[J].畜牧兽医学报,2004,35(2):134-140.
[9] 黄族豪,刘迺发,龙进,等.从线粒体DNA控制区基因比较石鸡和大石鸡的遗传变异[J].2006,28(3):420-424.
[10] 李吉均,文世宣,张青松,等.青藏高原隆起的时代、幅度和形式的探讨[J].中国科学,1979(6):608-616.
