乌苏里江秋大麻哈鱼生殖洄游群体结构特征与分析
2011-06-08王继隆唐富江潘志强
王继隆,唐富江 ,朱 翥 ,潘志强 ,刘 伟
(1.中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 150070;2.农业部黑龙江流域渔业资源环境科学观测实验站,黑龙江 哈尔滨 150070;3.黑龙江省抚远县鲑鱼放流站,黑龙江 抚远 156500)
大麻哈鱼(Oncorhynchus keta Walbaum)是典型的溯河洄游鱼类,幼鱼春季降海、成鱼秋季溯河洄游至出生地产卵直至生命终结,终生只繁殖一次。其广泛分布于白令海、北太平洋、鄂霍次克海、日本海及沿岸河流中[1],其中加拿大、美国、日本、俄罗斯数量分布较多。中国境内的大麻哈鱼可分为4个地方种群,即黑龙江、乌苏里江、绥芬河、图们江[2]。在乌苏里江大麻哈鱼幼鱼春节随水流降海,最终在黑龙江河口进入鄂霍次克海,而后在西太平洋、白令海等进入海洋生活阶段,直至性成熟后上溯至出生地产卵繁殖。
国外学者对大麻哈鱼洄游群体生物学特性进行了较深入的研究[3-5],上世纪90年代学者研究发现太平洋大麻哈鱼个体小型化及成熟年龄增大的现象,认为影响因素包含环境因子、捕捞选择性和种群压力等[6-10]。近年来日本学者研究发现大麻哈鱼小型化现象主要受到种间竞争的影响[11-12]。中国对大麻哈鱼资源调查也开展了一些研究,成熟个体小型化等现象也逐渐被揭示[13-15]。但对大麻哈鱼资源调查尚缺乏持续性和深入性,尤其是近几十年来大麻哈鱼资源增殖放流工作的不断开展,同时伴随着捕捞强度的居高不下,大麻哈鱼资源及种群结构变动趋势更加难以预料。笔者对2010年乌苏里江大麻哈鱼回归群体捕捞情况进行了持续监测,并对其回归群体数量结构进行分析,旨在了解大麻哈鱼洄游群体数量结构及生物学特征,为研究大麻哈鱼资源提供参考。
1 材料与方法
1.1 调查时间及区域
该研究在大麻哈鱼禁渔期(2010年10月1日到2010年10月20日)跟踪调查6艘增殖放流亲鱼采捕船只作业情况。禁渔期增殖放流捕捞船只数量固定,且作业地点也统一集中在通江口江段(其他江段禁捕)。
乌苏里江是一条中俄界河,发源于俄罗斯远东地区的锡霍特山脉,自南向北于黑瞎子岛地区注入黑龙江,最终注入鄂霍次克海。洄游期大麻哈鱼从黑龙江河口上溯至黑、乌交界处,然后分别游向黑龙江和乌苏里江。研究调查地点位于黑龙江省抚远县通江口江段(48.3°N,134.7°E)(图 1),也是中国境内大麻哈鱼洄游至乌苏里江的起点。
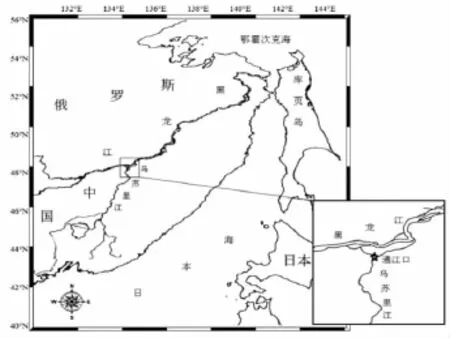
图1 调查站点
1.2 数据收集
大麻哈鱼捕捞网具为3层流刺网,网目大小为12.67 cm,网高3.9m,长约500m,作业时网具随水漂流,同时网具下纲触底。由于大麻哈鱼捕捞船大小、主机功率及网具较为统一,所以单位捕捞努力量渔获量(CPUE)统一为kg/(船·日),调查期间逐日统计了6艘捕捞船只捕获雌、雄大麻哈鱼质量。在调查期间每天随机收集一部分大麻哈鱼样本,共采集样本505尾,其中雄鱼237尾,雌鱼268尾,测量体长、体质量,并采集背鳍末端至臀鳍之间且侧线上方6~10枚鳞片供年龄鉴定。
1.3 处理方法
对6条渔船的雌、雄大麻哈鱼产量进行平均值处理,并根据雌雄个体平均质量换算为雌雄个体数量。然后对收集的大麻哈鱼样本进行测量,分别统计雌雄个体体长、体质量、年龄等生物学指标,并且通过方差分析判断不同洄游时期的大麻哈鱼的生物学指标的差异性。
2 结果与分析
2.1 回归群体数量特征
由于10月1日和10月20日捕捞船只的作业时间和其他日期作业时间不一致,故未将其统计在内。回归群体的数量特征反映了大麻哈鱼的群体结构特点,具体特征如下。
从图2可以看到禁渔期开始时回归群体的数量最多,10月9日处于低潮期,10月2日到9日基本成递减趋势,10日到15日成递增趋势,而后基本呈波动下降趋势,直至渔期结束。该调查发现9月25日大麻哈鱼开始形成渔汛,随后回归数量增减增多。由于缺乏9月25日到10月1日的数据,未能给出整个渔期的数量结构图,但从中也可看出大麻哈鱼回归数量出现两个波峰,即10月初出现一次大高峰,10月15日出现一次小高峰。禁渔期过后大麻哈鱼数量很少,已不能形成渔汛。因此,根据大麻哈鱼洄游数量结构将洄游期分为两个阶段,即10月10日前为第一阶段,10月10日后为第二阶段。
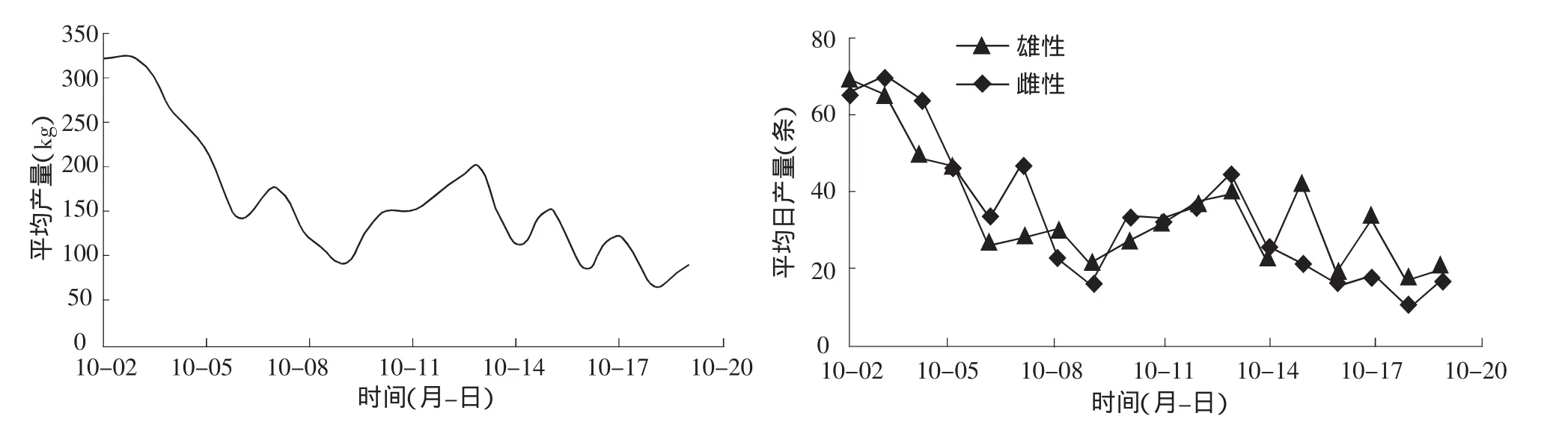
图2 大麻哈鱼日均产量及雌雄个体数量分布
在整个调查期内大麻哈鱼雌、雄个体数量比例为1.01∶1。从图2也可以看出,雌、雄大麻哈鱼的数量以及和总捕捞量变化趋势基本一致,第一阶段和第二阶段大麻哈鱼雌、雄个体间数量比例变动是随机的。
2.2 不同洄游期阶段群体间的差异性分析
对洄游期两个阶段的大麻哈鱼的各项生物学指标(性比、叉长、体质量和年龄)进行了方差分析(表1),方差分析结果p值均大于0.05,表明各指标差异性均不显著。
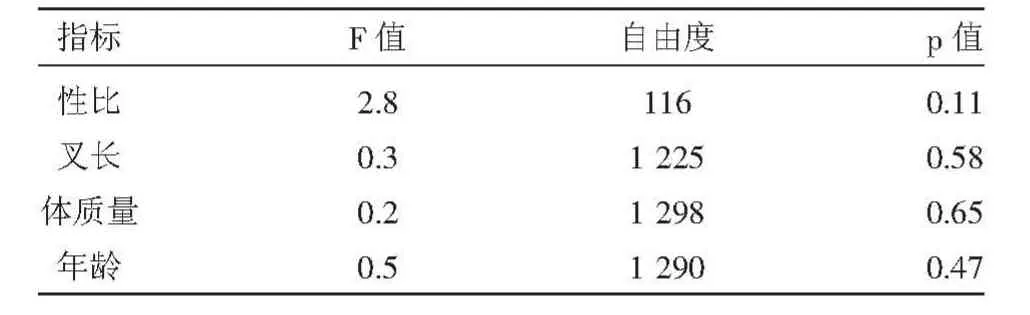
表1 两洄游阶段的大麻哈鱼各项生物学指标的方差分析
2.3 大麻哈鱼洄游群体的生物学特征
2.3.1 年龄组成 总体上说,大麻哈鱼年龄范围为2+~4+龄,其中 2+占 42.8%、3+占 53.4%、4+占 3.8%,可见3+为优势年龄组。方差分析表明雌雄群体间的年龄组成差异显著(p<0.01)。雄性个体中2+占53.4%、3+占 42.7%、4+占 3.8%,2+为优势年龄组;雌性个体中2+占 34.2%、3+占 62.1%、4+占 3.7%,3+为优势年龄组。可见雄性大麻哈鱼成熟期比雌性要短。

表2 大麻哈鱼的年龄组成 (%)
2.3.2 体长、体质量 大麻哈鱼雌、雄性个体的叉长、体长、体质量情况如表3所示,雄性个体平均叉长、体长、体质量分别为60.6 cm、63.5 cm、2 309.5 g;雌性个体平均叉长、体质量分别为62.4 cm、65.5 cm、2 532.3 g。可见,雄性个体平均体长和体质量都小于雌性个体。雄性个体叉长、体质量主要集中在 58~64 cm、2 150~2 750 g;雌性个体叉长、体质量主要集中在 59~65 cm、2 250~2 750 g。

表3 雌、雄大麻哈鱼的体长、叉长、体质量分布
3 讨论
3.1 大麻哈鱼生殖洄游群体数量特征
根据大麻哈鱼回归群体数量结构特征,将洄游期分为10月10日前、后的两个阶段。而以往的研究将这两个阶段统一起来,没有深入区分[13,15],这种分期现象更能详细反映实际捕捞状况。不同洄游阶段的大麻哈鱼的各项生物学指标方差分析表明其差异性并不显著,这也说明不同洄游期间的大麻哈鱼只是数量结构上的变化,生物学指标没有明显差异。因此,这种数量的波动可能是由不同洄游群体间的分散效应造成。此外,俄罗斯在黑龙江河口对大麻哈鱼回归群体也有一定的捕捞,其对洄游至中国乌苏里江的大麻哈鱼数量有直接影响。
3.2 洄游群体生物学特征
3.2.1 成熟年龄低龄化 20世纪60年代以来,洄游至中国大麻哈鱼的性成熟平均年龄呈现是在波动中下降趋势(图3),2010年大麻哈鱼性成熟年龄较近几年呈下降趋势[13],并且达到历史最低值。90年代国外有研究发现大麻哈鱼性成熟年龄有升高的趋势[16-17]。大麻哈鱼种群分布的不同也会造成不同的结果[18]。对成熟年龄变化的原因研究相对较少,一直没有明确的结论。渔业捕捞的持续选择性会造成大麻哈鱼性成熟年龄的降低[18-23],这是因为高年龄鱼个体大,被捕捞的风险也较大,所以渔业捕捞的长期选择性会使得性成熟年龄降低,但是成熟年龄大小的波动却不能完全解释。
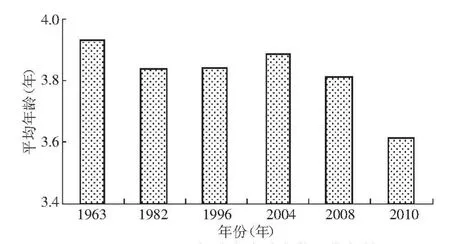
图3 不同年份大麻哈鱼的平均年龄
有学者通过模型模拟出大麻哈鱼性成熟年龄和生长速度之间的负相关关系[24]。20世纪90年代国外学者分析了大麻哈鱼性成熟年龄变大和体长减小的现象[10,16]时,指出这是减缓生长速度应对环境变化的响应[22]。根据这种理论,近年来乌苏里江大麻哈鱼成熟年龄的降低是由于生长速度变快造成的,同样可以解释图3中成熟年龄的波动原因。然而在生态系统中影响大麻哈鱼生长速度的因素非常复杂,有遗传因素和栖息地环境等因素共同影响。3.2.2 群体结构小型化 2010年乌苏里江秋大麻哈鱼生殖洄游群体平均体长、体质量与历史资料相比呈减小趋势,这也顺延了20世纪60年代以来的总体下降趋势(图4)。一方面是因为群体年龄结构的降低导致群体结构小型化;另一方面就是生活环境导致鱼类个体小型化,由于缺乏历史资料,只能对此简单探讨。
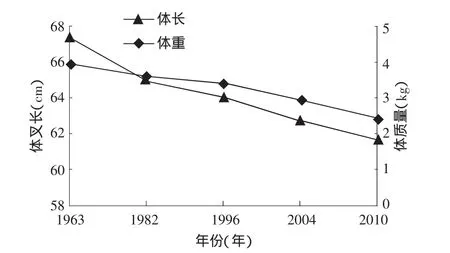
图4 乌苏里江大麻哈鱼不同年份的平均叉长和体质量
国内外对太平洋大麻哈鱼体长小型化现象也早有报道[16-18]。针对鱼类小型化现象的原因,有研究发现和环境有相关关系,如温度、盐度等。但是成熟大麻哈鱼体长的连续下降不能完全用环境变化来解释,捕捞选择性所引起种群遗传结构的变化而造成的成熟群体小型化也是不容忽视的[7,19-20],捕捞活动引起的高死亡率和个体大小的选择性是引起鱼类个体大小遗传进化的两个因素[21-22]。随着太平洋大麻哈增殖放流活动力度的不断增强,大麻哈鱼群体数量的不断增加,由此带来种间竞争压力的增大也是导致鱼体小型化的因素之一。
据了解,中国在20世纪60年代曾在乌苏里江对大麻哈鱼进行过增殖放流,之后基本上停止了这项活动,而俄罗斯对大麻哈鱼放流也主要集中在黑龙江流域。因此,目前中国境内乌苏里江大麻哈鱼基本上是天然种群。同时由于乌苏里江大麻哈鱼种群数量相对较少,在海洋中和其他大麻哈鱼种群地理分布不同[,18],种间压力相对较小。因此,种间竞争不应是大麻哈鱼小型化的主要原因。而中国大麻哈鱼的捕捞强度一直保持在较高水平,捕捞死亡率大于可开发率从而造成渔业资源的衰退,同时捕捞网具对个体大的大麻哈鱼选择性较高,小个体鱼类逃逸的可能性要大于大个体种类。因此,在中国捕捞选择性应该是造成大麻哈鱼小型化的主要原因。
综上所述,大麻哈鱼成熟期体长、体质量、年龄的变化是环境因子、生物间的相互作用、选择性捕捞的综合作用的结果。在生态系统中,鱼类小型化、成熟年龄变化可以说是对周围环境的一种响应,由于生态系统复杂性,很难确切说明这些生物学特征变化的具体原因。因此,这方的研究需渔业生态研究学者的共同努力。
[1] Shigehiko U,Tomonori A,Penelope A,et al.Origin and Distribution of Chum Salmon in the Bering Sea during the Early Fall of 2002:Estimates by Allozyme Analysis[R].National Salmon Resources Center,Toyohira-ku,Sapporo 062-0922,Japan,2004.
[2] Chen JP,Sun D J,Dong C Z,et al.Genetic analysis of four wild chum salmon Oncorhynchus keta populations in China based on microsatellitemarkers[J].Environmental Biology of Fishes.2005,(73):181-188.
[3] Hideji T,Yasuaki T,Yasuhiko N.Behavioural thermoregulation of chum salmon during homingmigration in coastalwaters[J].The Journal of Experimental Biology.2000,(203):1825-1833.
[4] Manu E.Observations of spawning behaviour in Salmoninae:Salmo,Oncorhynchus and Salvelinus[J].Fish Biology and Fisheries.2005,(15):1-21.
[5] Manu E.Two examplesof fixed behavioural patterns in salmonines female false spawning and male digging[J].Japan Ethological Society and Springer-Verlag.2007,(25):63-70.
[6] Brian SB,David W W,John H H.A review of size trends among North PacificA review of size trends among North Pacific salmon(Oncorhynchus spp.)[J].Canadian Journal of Fisheries and Aquatic Sciences.1996,(53):455-465.
[7] Richard L.Fishing selection and phenotypic evolution[J].Journal of Marine Science.2000,(57):659-668.
[8] Shoko H M,Kentaro M,Hiroyuki S.Growth of chum salmon(Oncorhynchus keta)correlated with sea-surface salinity in the North Pacific[J].Journal of Marine Science.2001,(58):1335-1339.
[9] Ishida Y,Welch DW,Ogura M.Potential influence of North Pacific sea-surface temperatures on increased production of chum salmon(Oncorhynchus keta)from Japan[J].Climate change and northern fish populations.1995,(121):271-275.
[10] Ishida Y,Ito S,Kaeriyama M,et al.Recent changes in age and size of chum salmon (Oncorhynchus keta)in the North Pacific Ocean and possible causes[J].Fisheries and Aquatic Sciences.1993,(50):290-295.
[11] Hyunju S,Hideaki K,Masahide K.Spatiotemporal change in growth of two populations of Asian chum salmon in relation to intraspecific interaction[J].Fish Sci.2009,(75):957-966.
[12] Hyunju S,Hideaki K,Masahide K.Long-term climate-related changes in somatic growth and population dynamics of Hokkaido chum salmon[J].Environ Biol Fish.2011,(90):131-142.
[13] 董崇智,赵春刚,王 金,等.乌苏里江大麻哈鱼的溯河生殖群体结构[J].中国水产科学,1999,6(3):5-9.
[14] 赵春刚,潘伟志,陈 军,等.2004年乌苏里江大麻哈鱼生殖群体结构研究[J].水产学杂志,2005,18(2):59-64.
[15] 唐富江,刘 伟,徐凤龙,等.黑龙江水系(中国)秋大麻哈鱼回归群体变动分析[J].上海海洋大学学报,2010,19(1):74-79.
[16] Kaev A.Dynamics of some biological indexes of Oncorhynchus keta in connection with the formation of its numbers[J].Journal of ichthyology.1999,39(88):642-651.
[17] John H H,Margaret SH.Size decline and older age atmaturity of two chum salmon(Oncorhynchus keta)stocks in western North America[J].Fish Aquature.1995,(121):245-260.
[18] Shogo M,Shunpei S,Moongeun Y,et al.Nonrandom distribution of chum salmon stocks in the Bering Sea and the North Pacific Ocean estimated using mitochondrial DNA microarray[J].Fish Science.2009,(75):359-367.
[19] Ricker W E.Changes in the average size and average age of Pacific salmon[J].Journal of Fisheries and Aquatic Sciences.1981(38):1636-1656.
[20] David O C,Stephan BM.Sustaining Fisheries Yields Over Evolutionary Time Scales[J].Science.2002,297(5578):94-96.
[21] Terry D B,Clyde B M.A genetic analysis of body size in pink salmon(Oncorhynchus gorbuscha)[J].Genome.1988,(30):31-35.
[22] Kentaro M,Masa-aki F.Why age and size at maturity have changed in pacific salmon[J].Marine Ecology Progress Series.2007,(335):289-294.
[23] Jorgensen T.Long-term changes in age at sexual maturity of Northeast Arctic cod(Gadusmorhua L.)[J].Conseil International pour l'Exploration de la Mer.1990,(46):235-248.
[24] Morita K,Morita S,Fukuwaka M,et al.Rule of age and size at maturity of chum salmon(Oncorhynchus keta):implications of recent trends among Oncorhynchus spp.[J].Canadian Journal of Fisheries and Aquatic Sciences.2005,(62):2752-2759.
