机械蜻蜓悬停时的气动力实验研究
2011-04-15姚大鹏申功炘朱百六
姚大鹏,申功炘,朱百六,郭 鹏,陆 远
(1.北京航空航天大学国家流体力学研究所,北京 100083;2.中国航天空气动力技术研究院,北京 100074)
0 引 言
近年来,微型飞行器(MAV,Micro Air Vehicles或Micro Aerial Vehicles)成为航空领域研究的热门,可看作是自然界的MAV的昆虫自然成为了我们研究的重要对象。在过去的几十年中,昆虫的升力机理渐渐开始为人们所知[1-4]。对于蜻蜓这类双翼昆虫的研究虽然相对而言比较初步,但也取得了一定进展。
近期,数值模拟方面[5-7]取得了一些结论,比如前后翼相互作用不大且有害于气动力[5];实验方面,英国小组[8]使用烟线法对活体蜻蜓的流场进行观测,德国小组[9]对机电蜻蜓翼模型进行二维PIV测量和测力。从以上的工作可以看出,测力结果可以比较直观地反应蜻蜓前后翼干扰的特点;然而,只有结合流场信息,干扰的机制才能得到进一步的解释。
笔者对蜻蜓悬停时双翼采取不同拍动模式产生的气动力进行了定量测量,并结合流场信息分析蜻蜓的升力机理。
1 实验设备和模拟方法
1.1 蜻蜓的运动学和形态学特征
研究对象的蜻蜓品种为Aeschna juncea,外形如图1所示,两翼前后排列,前翼翼根略靠上,两翼的拍动平面平行且与两翼翼根连线垂直。
蜻蜓翼的拍动一般有 3个自由度:(1)拍动(Translation)、(2)扭转(Rotation)和(3)拍动面偏离(Stroke plane deviation)。由于第3个自由度的运动对结果影响较小,实验不考虑拍动面的偏离。
图2显示了蜻蜓身体水平(OX向)时左侧单翼的运动,翼轴在拍动平面F(与水平面 H夹角β=60°)内拍动,整个翼绕这个距离前缘1/4c的轴转动。
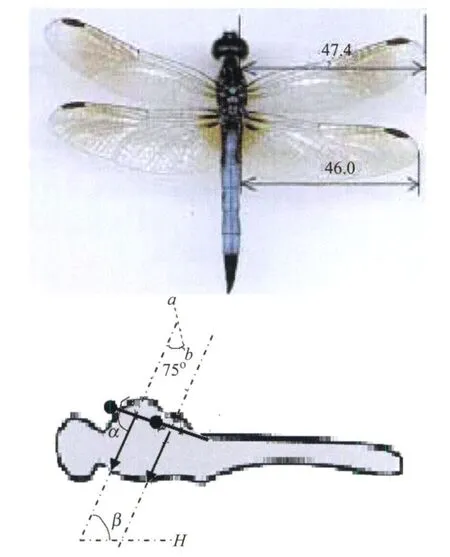
图1 蜻蜓形态特征Fig.1 Appearance of dragonfly
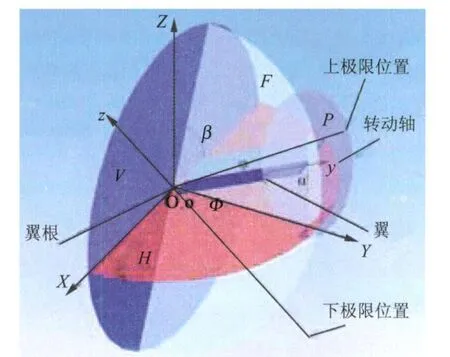
图2 实际蜻蜓的运动示意图Fig.2 Sketch of dragonfly motion
下拍开始时,翼处在拍动的上极限位置,翼平面与拍动平面成75°夹角,以此位置为原点,以下给出两个自由度运动的角位移函数。
拍动方位角φ

扭转的运动函数为

双翼拍动的情况下两翼会存在相位差[11],即产生了不同的拍动模式,该试验观察的有:同相,相差1/4周期和相差1/2周期(即反相)。
1.2 模型机构
由于实验为悬停飞行和设计斜向拍动机构困难,所以实验模型采用类似于Maybury和 Lehmann实验[9]中两翼上下布局的形式,即将相当于将蜻蜓身体在俯仰方向上仰转 60°,使得前翼在上,后翼在下,且两翼拍动平面与水平面平行。如图3所示,蜻蜓模型的拍动轴1竖直插入水中,两翼分别在轴2和3所处的水平平面内来回地拍动(两翼共轴但可分开控制),扭转轴2和3水平,相距5/4c,前后翼在拍动的同时各自绕扭转轴转动。这就是本机构模拟的两个自由度的运动。
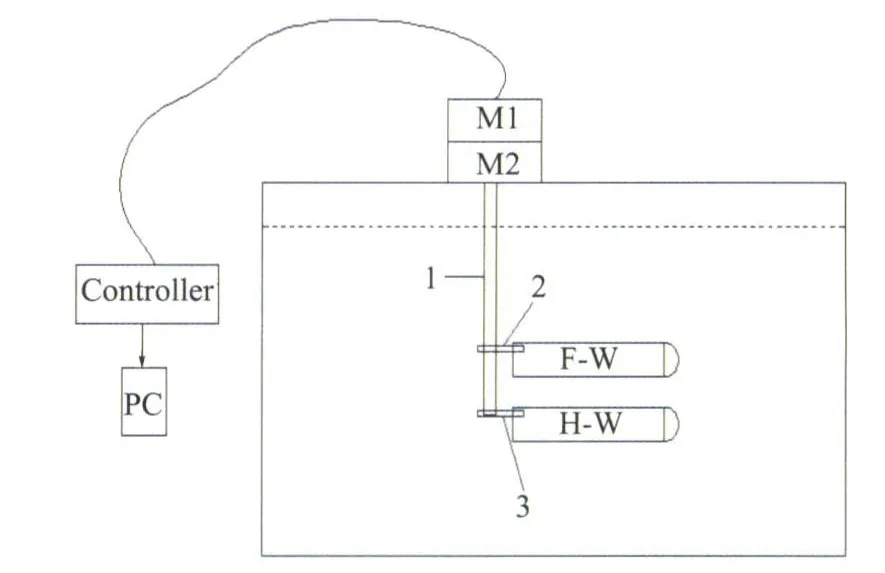
图3 前翼-后翼的布局(正视)Fig.3 Schematic of dragonfly model
机构置于1m×1.2m×1m(长×宽×高)的水缸中。而为克服传动机构和电机的防水问题,将主要传动机构和电机置于水面以上,并采用四连杆机构将电机的输出传到水下翼的转轴处,并可以通过更换不同长度的连杆调整前后翼之间的距离。
在观测所得的Aeschna juncea翼形数据[10]的基础上对蜻蜓模型翼作了如下简化:(1)前后两翼几何形状完全相同(实际上蜻蜓后翼的面积比前翼的大);(2)翼面形状为翼梢处为半圆的矩形;(3)不考虑翼面的变形,假设翼为刚性平板。同时为了使模型翼具有质量轻和抗弯曲的特性,测力用模型翼采用碳纤维制作,尺寸数据如表1。
为结合分析流场信息与所测气动力的关系,实验平台还安装有染色液流动显示系统。染色液从模型翼前缘根部流出,用于显示拍动产生的前缘涡核线。
1.3 相似准则

表1 模型翼的几何尺寸Table 1 The statistics of a model wing
2 测力实验步骤与数据处理
测力实验使用特殊设计三分量水下天平和江苏联能公司的YE6267数据采集系统进行测量及采集,模型翼的拍动频率为0.5Hz,Re=9750,数据采集频率为200Hz,所以每个拍动周期对每个分量要采400个数据点。由于模型翼在拍动中所受到的力包括重力、浮力、惯性力等,实验通过测量模型静止所受的力得到重力和浮力,并测量铜棒在拍动中所受的力得到近似的惯性力,具体的实验步骤如下:
(1)在空气测量中单翼在一个周期内的40个相位点静止时受到的力Sa;
(2)测量铜棒在空气中拍动所受到的力Da;
(3)在水中测量单翼在一个周期内的40个相位点静止时受到的力Sw;
(4)测量翼在水中拍动时受到的力Dw。
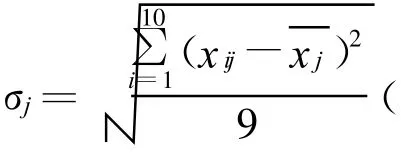
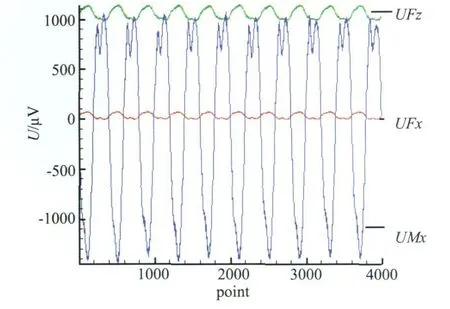
图4 所选取的10个周期点原始动态数据Fig.4 Original data in 10 stroke-cycles

表2 10周期天平测量数据的平均标准偏差Table 2 Mean values of standard error of each component
2.1 消除重力和浮力
模型静态时的测量值代表重力和浮力,把动态数据与静态数据相减的过程中自然可以消除这部分干扰。但翼在一个周期内不同相位时的姿态不同,天平的静态值也就不同,因此在一个周期内等间隔的取了40个相位,测量其静态值Sw,并线性插值到400个点(即一个周期内动态值的个数),然后将对应相位的动态值Dw与静态值Sw相减得到重力浮力以外的力。
2.2 力的轴系转换
由于拍动时天平是随模型翼一同运动的,因此天平直接所测的力Fz和Fx是相对翼的随体坐标系的力,首先将其转化为模型所在的地轴坐标的水平力H和竖直力V
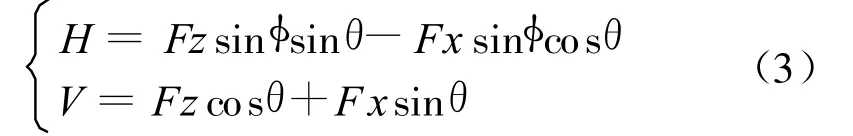
其中θ为扭转角(θ=α+5/6π),φ为拍动角。而模型蜻蜓的体轴坐标系和地轴坐标系有夹角β,因此相对蜻蜓体轴的升力L和阻力D有

变换前后的力曲线见图5。
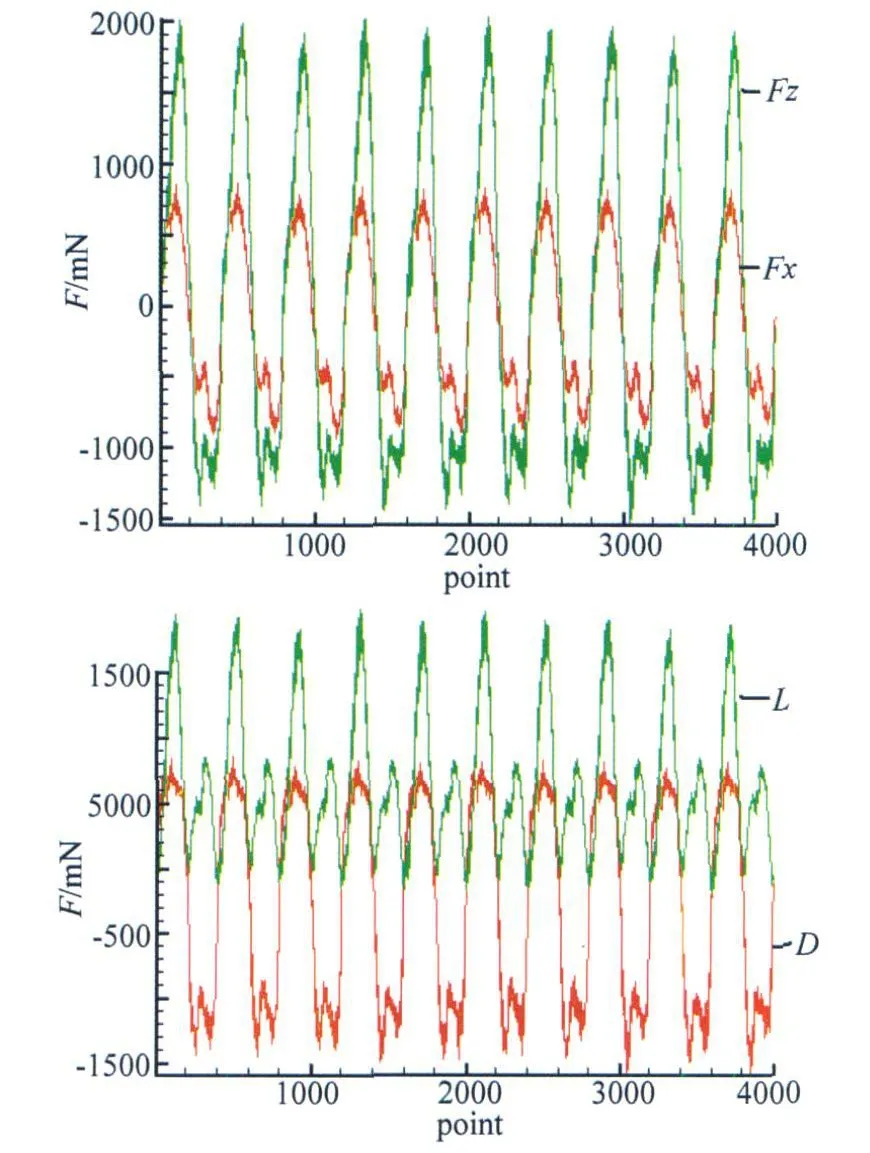
图5 坐标系变换前后力曲线Fig.5 Forces in different coordinates
2.3 消除惯性力
为了测得翼拍动的惯性力G,首先在空气中进行了实验,并采用了一根质量、质心位置都和翼相同的铜棒代替模型翼,由于截面积小,且处于空气中,拍动产生的气动力Q可略去不计,那么有G=Da-Sa。以后在水中用模型翼拍动时,其所受气动力Q=Dw-Sw-G。图6给出了一个周期内(单翼)气动力和惯性力的曲线,该曲线由10周期平均得来,并经过滤波平滑。由图中可以看出,惯性力对测力结果的影响虽然比较小,但也并不能忽略不计。
3 气动力测量结果
3.1 单翼拍动
图7给出了单翼在一个周期内的升力和阻力变化曲线,图中实线代表升力系数,虚线代表阻力系数,时间零点时,翼处于上拍的极限位置,即紧接着将进行下拍运动。
升力在下拍和上拍过程中各有一个峰值。整个周期而言升力基本都为正值,且下拍时产生的升力要大于上拍时产生的升力,周期平均升力系数为1.7926。
阻力在下拍时为正,上拍时为负,虽然对于悬停中的蜻蜓而言,整个周期阻力基本抵消掉,但明显可以看出上拍时的负阻力比较大,即总体而言蜻蜓会受到一个向前的推力。整个周期的平均阻力系数为-0.8738。这可能是实验所选取的运动函数与实际有所出入所致。
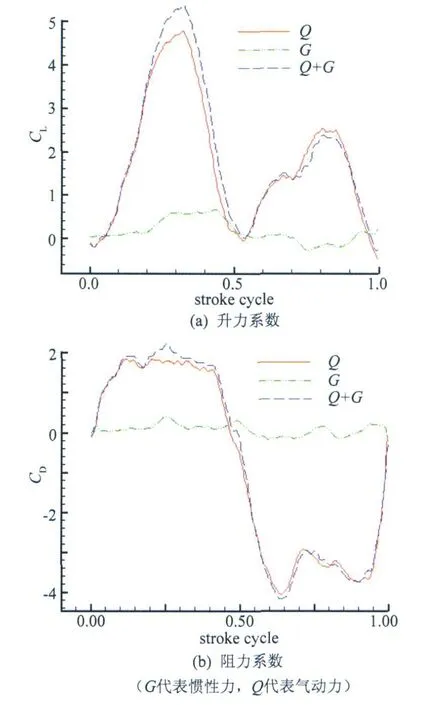
图6 一个周期内气动力和惯性力引起的阻力升力变化曲线Fig.6 Lift and drag caused by aerodynamic force and inertia force
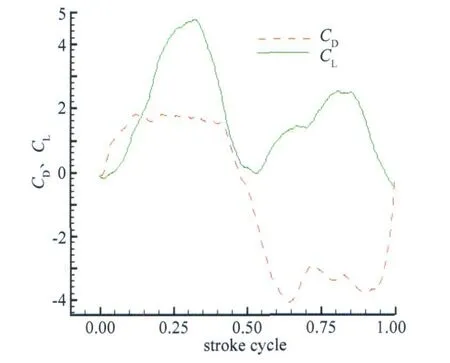
图7 单翼在一个周期内的升力和阻力系数变化Fig.7 Lift and drag averaged in 1 stroke cycle
3.2 双翼的测量结果
为方便比较,在此将各种拍动模式下前翼和后翼的气动力系数曲线放到相同的相位上作一下比较。前后翼在各种拍动模式下的升力和阻力系数如图8。
(1)前翼
升力方面,双翼拍动时下拍的升力峰值都有所下降,但升力的上升普遍比单翼要早。反相拍动时上拍的升力极值有明显减小。
前翼在各种拍动模式下,下拍时阻力曲线基本保持一致,同相拍动的阻力最大,反相和后翼领先1/4周期相对单翼变化不多。上拍过程则和单翼相差较大,但共同点在于上拍前半段会出现一个波动,并且较晚达到负向极值,各种拍动模式中仍是同相拍动拥有最大的负向极值。整个周期而言,前翼在双翼拍动时推力大大减小。

图8 前后翼不同拍动模式下气动系数比较Fig.8 Force coefficient in different flapping mode
(2)后翼
升力方面,只有同相拍动时升力上升有所提前,其它的拍动模式下反而有所滞后。双翼反相拍动时后翼升力峰值无论上拍和下拍都有明显下降,其它拍动模式峰值却没有减小,后翼领先前翼1/4周期拍动时,上拍的升力峰值反而有所增加。
在下拍阶段,反相拍动的阻力有明显减小,后翼领先前翼1/4周期也有一定减小,同相拍动则和单翼相似,上拍阶段后翼阻力变换不像前翼那么明显,同相和后翼领先1/4周期拍动时负向极值有所增大,反相和后翼领先1/4周期拍动出现明显波动,出现时间点都是前后翼交错的时候,很明显这是翼间相互干扰所至。
4 结果分析和比较
各拍动模式下前后翼的平均气动力系数如表3。
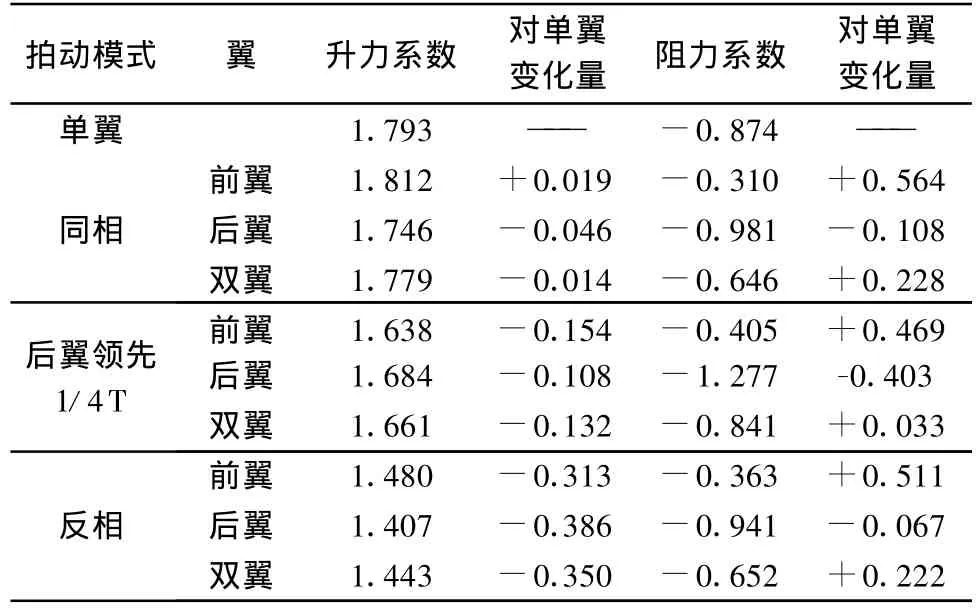
表3 周期平均气动力系数Table 3 Mean values of force coefficient in 1 stroke cycle
总体看来,双翼同相拍动时具有最大的升力,其中前翼的升力系数甚至超过单翼,之前 Reavis和Luttges[12]所进行的活体蜻蜓测力实验也表明同相拍动模式能提供最大的升力。后翼领先1/4周期的模式具有最大的推力,这种拍动模式可能比较适合前飞,这与一系列研究和观察的结论相符合[13]。反相拍动阻力和升力都比较小,这种受力较小的拍动模式被普遍认为适合于巡航。
Maybury等人在2004年做过类似测力试验[9],但其采用的运动参数与本试验不同,上拍与下拍对称,因此所得结果也是对称的。而本试验采用非对称拍动周期,得到更多的升力和推力,说明这种拍动方式更接近实际蜻蜓飞行所用拍动方式。
目前所得到的测力结果已经给出了一个周期内模型翼上升力和阻力的变化,但这还不能详尽地揭示蜻蜓双翼拍动相互干扰的机理。因此,对下拍阶段的前缘涡演化进行了流动显示。
实验平台包括染色液流动显示系统,染色液从模型翼前缘根部流出,用于显示拍动产生的前缘涡核线。单翼前缘涡,在下拍开始阶段卷起,随后沿展向发展,最后破裂。所观察到的现象与文献[14]一致。
对于双翼的流动显示各种情况,这里给出以涡核线最明显的0.4T时刻(如图9)进行说明。

图9 双翼各种拍动模式下0.4T时的前缘涡核Fig.9 Flow visualization of leading-edge vortex at 0.4T in different flapping mode
在后翼领先前翼1/4周期时,后翼的前缘涡核线有所加强,但就测力的结果而言,其升力并不是最大,这或许和其发展较迟有关,就升力峰值而言它与单翼的情况差不多,这可用单翼机制来进行解释。
两翼同相下拍时(俯转结束后的纯平动阶段),前后翼都与水平面平行,并处于同一个平面内,前翼后缘和后翼前缘相距仅1/4c,这种情况更像是前后翼组成了一个小展弦比的平板翼,使得前翼的前缘涡能更好地附着在翼面上发展。无论计算还是实验,这种模式下的前翼升力都是各拍动模式下最高的。
反相拍动时的后翼前缘涡比较松散,无论升力和阻力都明显比其它拍动模式要小。
5 结 论
实验对蜻蜓悬停时两翼所受气动力进行了测量,并得出以下结论:
(1)蜻蜓翼(单翼)拍动所受到竖直方向上的升力会在下拍和上拍中段稍后一些的时刻达到两次极值,下拍升力极值大约是上拍的两倍,整个周期来看翼上获得的升力主要来自下拍阶段。水平方向上,下拍产生阻力,而上拍产生推力,整个周期而言蜻蜓受到一定推力,水平方向力不平衡的原因可能是我们所选取的运动曲线与实际有出入有关;
(2)双翼拍动时,同相拍动产生了最大升力;后翼领先前翼1/4周期的升力次之,但拥有最大的推力,说明这种拍动模式适合于机动飞行;反向拍动的升力和阻力都是最小,被认为适合于省力的巡航飞行。双翼在有相位差拍动时,后翼与前翼相遇的时刻气动力会产生波动,反相上拍中的后翼升力和阻力都有明显降低。
[1] ELLINGTON C P,VANDER B C,WILLMOTT A P.Leading-edge vortices in insect flight[J].Nature,1996,384:626-630.
[2] DICKINSON M H,LEHMENN F O,SANE S P.Wing rotation and the aerodynamic basis of insect flight[J].Science,1999,284:954-1960.
[3] SUN M,TANG J.Unsteady aerodynamic force generation by a model fruit fly wing in flapping motion[J].Journal of Experimental Biology,2002,205:55-70.
[4] DUDDLY R.Biomechanics of insect flight[M].Princeton Press,2000.
[5] SUN M,LAN S L.A computational study of the aerodynamic forces and power requirements of dragonfly(aeshna juncea)hovering[J].Journal of Experimental Biology,2004,207:1887-1901.
[6] WANG J K,SUN M.A computational study of the aerodynamics and forewing-hindwing interaction of a model dragonfly in forward flight[J].Journal of Experimental Biology,2005,208:3785-3804.
[7] ISOGAI K,FUJISHIRO S,SAITOH T,et al.Unsteady three-dimensional viscous flow simulation of a dragonfly hovering[J].AIAA J,2004,42(10):2053-2059.
[8] THOMAS A L R,TAYLOR G K,SRYGLEY R B,et al.Dragonfly flight:Free-flight and tetheredflow visualizations reveal a diverse array of unsteady lift-generating mechanisms,controlled primarily via angle of attack[J].Journal of Experimental Biology,2004,207:4299-4323.
[9] M AYBURY W J,LEHM ANN F O.The fluid dynamics of flight control by kinematic phase lag variation between two robotic insect wings[J].Journal of Experimental Biology,2004,207:4707-4726.
[10]NORBERG R A.The pterostigma of insect wings an inertial regulator of wing pitch[J].Journal of Comparative Physiology,1972,81:9-22.
[11]ALEXANDER D E.Unusual phase relationships between the forewings and hindwings in flying dragonflies[J].Journal of Experimental Biology,1984,109:379-383.
[12]SOMPS C,LUTTGES M.Dragonfly flight:Novel uses of unsteady separation flow[J].Science,1985,228:1326-1328.
[13]ALEXANDER D E.Unusual phase relationships between the forewings and hindwings in flying dragonflies[J].Journal of Experimental Biology,1984,109:379-383.
[14]LU Yuan,SHEN Gong-xin,SU Wen-han.Flow visualization of dragonfly hovering via an electromechanical model[J].AIAA J,2007,45(3):615-623.
