Bats and marsupials as indicators of endemism in the Yungas forest of Argentina
2010-12-25MarSandovalClaudiaSzumikRubBarquez
María L Sandoval, Claudia A Szumik, Rubén M Barquez,*
[1. PIDBA (Research Program of Argentinean Biodiversity), Natural Sciences Faculty and Miguel Lillo Institute, National University of Tucuman, Miguel Lillo 205, San Miguel de Tucumán, CP 4000, Tucumán, Argentina; andCONICET (National Council of Scientific and Technical Researches),Argentina; 2. INSUE (Superior Institute of Entomology “Dr. Abraham Willink”), Natural Sciences Faculty and Miguel Lillo Institute, NationalUniversity of Tucuman, Miguel Lillo 205, San Miguel de Tucumán, CP 4000, Tucumán, Argentina; andCONICET (National Council of Scientific and Technical Researches), Argentina]
Bats and marsupials as indicators of endemism in the Yungas forest of Argentina
María L Sandoval1, Claudia A Szumik2, Rubén M Barquez1,*
[1.PIDBA(Research Program of Argentinean Biodiversity),Natural Sciences Faculty and Miguel Lillo Institute, National University of Tucuman, Miguel Lillo205,San Miguel de Tucumán, CP4000,Tucumán, Argentina; andCONICET(National Council of Scientific and Technical Researches),Argentina; 2.INSUE(Superior Institute of Entomology“Dr. Abraham Willink”),Natural Sciences Faculty and Miguel Lillo Institute, National
University of Tucuman, Miguel Lillo205,San Miguel de Tucumán, CP4000,Tucumán, Argentina; andCONICET(National Council of Scientific and Technical Researches),Argentina]
Several studies have characterized the Yungas as a separate biogeographic unit, mainly based in floristic components. However, these characterizations were mainly qualitative and did not include faunal groups. The Yungas have been assumed as a region with rich floral and faunal diversity, but without testing how well they are described by animal distributions. Our study consists of a formal analysis of endemism based on distribution of small mammals in the southernmost portion of the Yungas. This area is biogeographically very interesting because the Yungas are comprised of discontinuous fragments of forests that extend into temperate arid and semiarid habitats. As a first approximation, we contrasted a group of volant species (bats) versus a group of non-volant species (marsupials). Our results show that small mammals are efficient indicators of endemism in Yungas. Eighty percent of the species of small marsupials included in the analysis supported the identified areas as being zones of endemism. Regarding bats, almost 55 percent of the species supported a designation of endemism. The results also show that the areas we considered are congruent with the botanical definition of the Yungas of northwestern Argentina and their subdivisions, an assumption that had not been previously tested with a formal quantitative method. We also found that non-volant species are better indicators of endemism than volant ones at regional scales, but volant species are better indicators than was previously thought.
Andean rainforests; Chiroptera; Didelphimorphia; Optimality criterion; Small mammals
It is well known that mammals are important as biogeographic indicators and their species, or higher taxonomic levels (genera, families, and orders), have been important in defining such areas at different scales.The Biogeographic Regions of the world have endemic orders, families, and subfamilies, while Subregions and minor divisions have endemic genera and species.Koopman (1976, 1981) and Proches (2005) found that bats are important for defining faunal regions, while other authors (Willig & Mares, 1989; Corbet & Hill,1992) suggested that bats are not useful for defining biogeographic zones or their limits within lowland tropical forest. Proches (2005), in his analysis of bat distributions at global scales, obtained regional groupings largely consistent with the zoogeographic regionalization model and the phytogeographic data are consistent with the zoological data. His results are coincident with that of Koopman (1976, 1981), who had already considered endemics among bats as defining faunal provinces of South America. On the other hand,Corbet & Hill (1992) pointed out that the distributions of non-flying mammals are more tied to the regional limits than are the distributions of flying mammals. They observed than the Australian and Asian faunas of volant species are more mixed than the terrestrial mammals and any limit is arbitrary. Similarly, Willig & Mares (1989)obtained little congruence between phytogeographic zones and distributional limits of bat species in Venezuela, so they argued that the distribution of bats were not limited to particular phytogeographic zones;moreover, they argued that no associations of bats can be used to define biogeographic zones.
Descriptions of the Yungas have been essentially qualitative (Cabrera & Willink, 1973; Cabrera, 1976;Vervoorst, 1979) and based on floristic components (see Ribichich, 2002). For animals the Yungas were then assumed to be a natural region, but they did not test the congruence of species distributions of animals or plants with the putative regions. Moreover, the large latitudinal extension and the great altitudinal development of the mountains of northwestern Argentina region provide great environmental heterogeneity, leading several authors (e.g. Cabrera & Willink, 1973; Brown, 1995a, b;Hueck, 1978; Ojeda, 1999) to consider it as a nonhomogenous unit, and to assume that its fauna was composed by species from the surrounding regions.However, other authors suggest that in the Yungas there are taxa that are more typical and common there than in marginal areas, although they may not be exclusive(Cabrera, 1976; Morrone, 2000, 2001).
The distribution of animals and plants in a region is the result of a complex interplay of historical,evolutionary, and ecological factors acting over different time and spatial scales (Hortal et al, 2010; Levin, 1992).A number of species with different dispersal capabilities which have similar distributional areas share a common history, besides similar ecological requirements. To share a common history implies that a number of taxa have been affected by the same historical processes that have operated for millions of years and that they have responded in similar ways. The congruent distributional areas of these taxa form an area of endemism. According to Platnick (1991) an area of endemism is defined by the areas of consistent distribution of two or more species spatially restricted to it. This definition implies sympatry,although not necessarily an exact congruence of the distributional areas of the species. Various delineations of areas of endemism over the years (Harold & Moi,1994; Morrone, 1994; Linder, 2001; Szumik et al, 2002)are based on the definition established by Platnick (1991)(Szumik & Roig-Juñent, 2005), but only Szumik et al(2002) formally compared the degree of superposition between the distributional areas of species with the application of an optimality criterion. This comparison is fundamental considering that sympatry is the basic requirement to establish areas of endemism (Szumik &Roig-Juñent, 2005).
The biogeographic assessment of a region is a starting point to formulate strategies for conservation of biodiversity. Consequently, in order to propose and develop programs of conservation and management, it is important to have quantitative descriptions of the different components and levels of biodiversity of a particular region (Ojeda, 1999). In this sense the areas of endemism are crucial (Vane-Wright et al, 1991; Linder,1995; Peterson and Watson, 1998). Previous studies dealing with identification of areas of importance for the conservation of mammals in Northwestern Argentina(NWA) that use quantitative methods over data of occurrence do exist (Ojeda et al, 2003; Tabeni et al,2004). Also, the areas of endemism are considered the fundamental units in the study of historical biogeography,because they are the entities to be compared in the biogeographic analyses. Finally, the identification of areas of endemism can be useful as additional evidence for studies that involve species of uncertain taxonomic status, and to predict the occurrence of species in areas where they have not been recorded (Díaz Gómez, 2007).
We have applied an explicit method to mammal distributions based on grids in order to identify areas of endemism. As a first approximation, we have contrasted selected small mammals, volant (bats) versus a group of non-volant (marsupials), from the Andean Subtropical Forests of NWA. We have evaluated whether small mammals in general are useful indicators of areas of endemism, whether the areas recovered are congruent with perceived natural areas (as botanically defined), and whether non-volant species are better indicators than volant one at the regional (meso) scale.
1 Materials and Methods
1.1 Study area
The study area (Figs. 1, 2) includes the provinces of Jujuy, Salta, Tucuman, Catamarca, and Santiago del Estero in northern Argentina. This region is a complex mosaic of habitats that intermingle, where large phytogeographic units converge, and some reach their boundaries of occurrence (Cabrera, 1976). The Yungas phytogeographic province (Cabrera & Willink, 1973;Cabrera, 1976) forms a narrow slash from Venezuela south to NWA, along more than 4,000 km, from 9º north latitude, and gradually disappearing at 28º south latitude in the province of Catamarca (Cabrera & Willink, 1973;Hueck, 1978). The southernmost part of the Yungas forests is recognized as a distinctive biogeographic and ecological unit (Cabrera, 1976). Moreover, Ojeda et al(2008) emphasized the distinctive character of this fraction of the Yungas, located in Argentina between 22ºand 28º south latitude (from the border line with Bolivia,at 22º south latitude, to the centre of Catamarca; Cabrera,1976), defining it as a “peninsula” because of its position in relation to the habitat matrix that surrounds it. This portion of the Yungas is then unique and biogeographically very interesting. The entire zone consists of an intergradation of Andean and pre-Andean mountains and lowland desert valleys and fragments of forests that protrude into temperate arid and semiarid habitats. In Argentina, the Yungas are bordered to the east by the phytogeographic province of Chaco and to the west by the Monte and Prepuna provinces and in some sectors by the Puna province (Cabrera, 1976). Altitude varies from 300 to 3,000 m and three altitudinal districts can be differentiated (Meyer, 1963; Cabrera, 1976; Brown,1995a, b; Prado, 1995). Additionally three latitudinal sectors were proposed that were associated with rainfall and orography: North, Centre, and South (Brown &Ramadori, 1989; Morales et al, 1995; Morales, 1996;Brown et al, 2001).
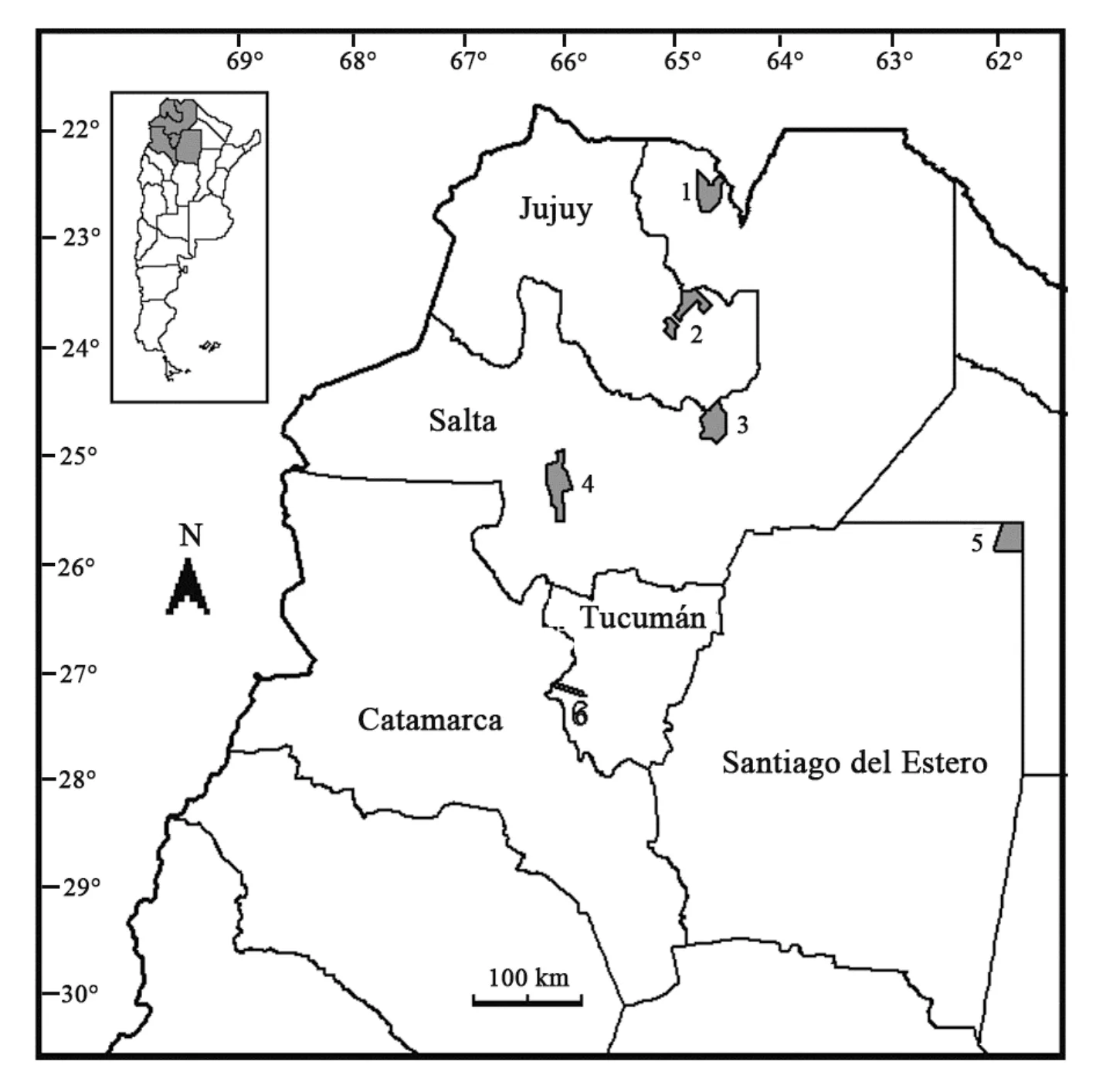
Fig. 1 Map of northwestern Argentina, showing the provinces of Jujuy, Salta, Tucuman, Catamarca, and Santiago del Estero The insert shows the relative position of the provinces within Argentina. National Parks are indicated with numbers 1 to 6 as follow: 1)Parque Nacional Baritú (Salta province); 2) Parque Nacional Calilegua (Jujuy province); 3) Parque Nacional El Rey (Salta province); 4)Parque Nacional Los Cardones (Salta province); 5) Parque Nacional Copo (Santiago del Estero province); 6) Parque Nacional Campo de los Alisos (Tucuman province).

Fig. 2 Map of northwestern Argentina showing the localities where bats and small marsupials were recorded (black points),as well as the phytogeographic province of the Yungas (solid gray fill)
1.2 Data
Advances in the knowledge of small mammals of NWA are well documented for the last several decades allowing the achievement of comprehensive biogeographic analysis. Taxonomic arrangement follows Barquez et al (2006). Taxonomic identifications were made by comparisons of morphology and external and cranial morphometrics with reference material, type descriptions of species, and published taxonomic revisions of particular species or species groups. Forty two species of bats have been cited for the region, 39 of which are in the Yungas (Barquez et al, 1993; Barquez et al, 1999a, b; Barquez & Díaz, 2001; Barquez, 2006; Díaz& Barquez, 2007). Marsupials are represented by 11 species, nine in the Yungas (Flores et al, 2000; Díaz et al,2002; Flores, 2003, 2006; Jayat & Miotti, 2006; Flores et al, 2007). The list of species and their distributional data were obtained from field surveys, and from literature and systematic collections. One thousand, one hundred and eighty nine (1189) records of 52 species of NWA were obtained, including the 42 species of bats and ten of the 11 small marsupials from the five NWA provinces of Jujuy, Salta, Tucuman, Catamarca, and Santiago del Estero (Fig. 2). Five species were registered at only one site (Appendix 1). Some georeferences were taken in the field with Global Positioning System (GPS) at the time of collection while others were obtained from gazetteers,maps, or satellite images.
1.3 Methodology
The methodology proposed by Szumik et al (2002),later modified by Szumik and Goloboff (2004), was used to identifying the areas of endemism determined by the distribution of bats and small marsupials in NWA. The method consisted in dividing the study region into cells,and counting the number of species that can be considered endemic to a particular set of cells. The sets with the highest number of endemic species were preferred (Szumik et al, 2002; Ruggiero & Ezcurra, 2003;Szumik & Goloboff, 2004). The method was implemented in two computer programs, NDM/VNDM v.2.5 (Goloboff, 2005).
In this study an analysis of endemicity was carried out by using a heuristic search. It was established that the sets had to be found by adding/eliminating one cell at a time, and that these sets had to be defined by two or more endemic species. Also, their endemicity value ought to be higher than or equal to that of two species with a perfect homopatrid distribution. Cell sets partially overlapping with more than 50 percent of species in common were ruled out, obviously retaining those with the highest endemicity value.
Given that there are no criteria to select an optimal grid and that there are patterns of quite different sizes,the data were analyzed using various different grids and fill options. We analyzed the database of the georeferenced records by using grids with cells of different shapes (square, horizontally, and vertically rectangular) and sizes (Tab. 1). The method considers three categories of records: absent, present (including inferred), and assumed (as present). A species may not have been recorded in a cell but its presence may be suspected or assumed. On the other hand, a species absent from one cell but present in surrounding cells can be inferred as present in that cell. The assumed and inferred records can be filled by NDM with a simple algorithm (Szumik & Goloboff, 2004). The program offers to the researchers the possibility to set the filler radius size as a percentage of the cell size. The radius sizes used for assumed and inferred presences when analyzing the database with the different grids considered are given in Tab. 1.
The origin of the grid was arbitrary defined and located atx=31.700 andy=20.530. For the mapping of the resulting areas, political and phytogeographic maps of NWA were used. The areas of endemism and the record points of the species analyzed were graphed on the maps using the Global Mapper v8.03 program.
2 Results
Marsupials have been useful at characterizing areas of endemism, and eight out of ten species had positive endemism scores at different spatial scales (Tab. 3).Regarding bats, our results show that 23 out of the 42 species were good indicators and permit the definition of areas clearly coincident with the Yungas of Argentina with striking accuracy (Tab. 3).

Tab. 1 Search parameters for the analysis with grid of cells of different shapes and different sizes(expressed in degrees)

Tab. 2 Summarized results obtained in the analysis of grids with cells of different shapes and sizes (expressed in degrees)and considering different values of filler for assumed and inferred presences
Several areas of endemism, remarkably coincident with the Yungas in almost all their extension, or with combinations of their latitudinal sectors, were obtained(Tab. 2; Fig. 3). It is worth mentioning that these results were almost identical, and stable, with all the search parameters used. The following areas were obtained:
A) This generalized area has been treated as a“Península” (Ojeda et al, 2008). It represents all of the areas of endemism obtained from the Yungas of NWA along their entire latitudinal extension (Fig. 3A), and it was obtained several times, with grids of square,horizontally, and vertically rectangular cells, with scores that range widely between 2.20 and 9.13 (Tab. 2).
All coalescent areas of endemism are included within this generalized area, even if some have individual limits which are slightly different from each other. The obtained areas that can be grouped here cover almost all the surface of the Yungas in NWA. They range from 22º S latitude in the north of Salta province,through the east of the province of Jujuy, centre of Salta,and centre and west of the province of Tucuman, south to approximately 28.5º south latitude in the province of Catamarca (Fig. 3A). None of these individual coalescent areas includes the southern portions of the Yungas formation (i.e., the Mountain Ranges of Ambato and Ancasti, in Catamarca) (Fig. 2).
While some of the obtained areas of endemism are contiguous, other have disjunctions in central Salta,between 24.5° and 25.5° south latitude (Fig. 3A), that may be related to at least two factors of importance. First,an important discontinuity is located between the northcentral and south-central mountain ranges of Salta province, resulting in an important discontinuity of the forests formation in the region (Fig. 2). Second, but related to the first, the amount of records for the area is quite low if compared with the known records for other sectors of the Yungas in Argentina, except the southernmost sectors of the Yungas in the province of Catamarca (Fig. 2).
The score of endemicity was given by 13 species of bats (Artibeus planirostris,Chrotopterus auritus,Eptesicus diminutus,Histiotus laephotis,Lasiurus blossevillii,Myotis albescens,M. keaysi,M. riparius,Nyctinomops laticaudatus,N. macrotis,Sturnira erythromos,S. lilium, andS. oporaphilum) and four marsupials (Micoureus constantiae,Thylamys cinderella,T. pallidior, andT. sponsorius) (Tab. 3, in part).
The highest individual endemicity values for bats(greater than 0.67) were found inA. planirostris,C.auritus,L. blossevillii,M. keaysi,S. erythromos,S. lilium,andS. oporaphilum.Artibeus planirostrisandC. auritusare mostly found in the Yungas in NWA, although Chacoan records also exist. However, these species have high endemicity values because the localities of the Dry Chaco are very close, and closely related to the Yungas.L. blossevilliiis a widely distributed species in Argentina,and most of its records in the northwest correspond to the Yungas, with just a few in the Dry Chaco. In Argentina,M. keaysiandS. erythromoshave been recorded only in the provinces of Jujuy, Salta, and Tucuman, with most of the records belonging to the Yungas, and just some to the Dry Chaco.Sturnira liliumis a very abundant species all over the entire north of the country and in the northwest inhabits primarily in the Yungas, and secondarily in the
Dry Chaco and in the Monte Desert; other species of the genus,S. oporaphilum, is exclusive of the Yungas in Argentina. Among marsupials,M. constantiae,T.cinderella, andT. sponsoriushave high endemicity values, which is consistent with previous distributional analyses (Flores, 2006; Flores et al, 2007) that conclude that these species are exclusive of the Yungas.

Tab. 3 Characterizers species of the three generalized areas of endemism obtained in the analysis with grids of squared cells of different sizes (expressed in degrees) and considering different filling values for assumed and inferred presences (not shown)
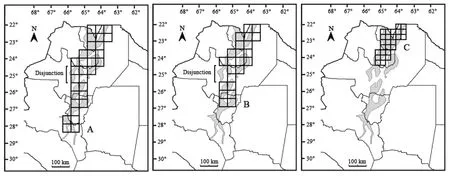
Fig. 3 Map of northwestern Argentina showing the first (A), the second (B) and the third(C) generalized areas of endemism obtained
B) This area represents all the obtained areas of endemism that includes the Yungas of NWA but do not include their southernmost portion in the provinces of Tucuman and Catamarca (Fig. 3B). This area was obtained a few times, with grids of square cells, with scores that range between 2.12 and 5.69 (Tab. 2).
As in A (the previous case) in this generalized area,all of the obtained areas of endemism, that occupy the same zone, are included. They run from the 22° south latitude in the north of Salta province, through the east of Jujuy province, and centre of Salta, to approximately 27°south latitude in the province of Tucuman (Fig. 3B).Some of obtained areas are contiguous, while others show disjunctions in the centre of the province of Salta,between 24.5° and 25.5° south latitude.
The species that contribute to the score of endemicity are six bat species (A. planirostris,Cynomops planirostris,Eumops patagonicus,Molossus rufus,Noctilio leporinus, andS. oporaphilum) and three marsupials (M. constantiae,T. cinderella, andThylamyssp.) (Tab. 3).
In Argentina,C. planirostrishas been recorded only in the provinces of Jujuy and Salta; its endemicity value is high (=0.78) because all of the confirmed records are from the Yungas, although this species can probably inhabit also the Dry Chaco (Barquez, 2006).E.patagonicusis widely distributed in Argentina, and have a very low individual value (=0.18) because in NWA its records are both in the Yungas as in the Dry Chaco, but without any predominance of records in the Yungas.Molossus rufusis distributed in central and northern Argentina; in the NWA it has been recorded mostly in the Yungas, with some records in the Dry Chaco. The records ofN. leporinusin NWA are also mostly found in localities of the Yungas forests and very limited individuals are from the Dry Chaco. The only three known records ofThylamyssp. also belongs to the Yungas region.
C) This third generalized area represents all the obtained areas of endemism that includes the northernmost portions of the Yungas in NWA (Fig. 3C),which are the north of Salta and the east of Jujuy provinces. The retrieved areas that can be grouped in this category runs from the 22° south latitude in the north of Salta, south to the 24.5° south latitude in the province of Jujuy (Fig. 3C).
The area was obtained with all the combinations of search parameters, with grids of square, horizontally, and vertically rectangular cells, and with scores that range between 2.01 and 5.88 (Tab. 2). The species that contribute to the score of endemicity are eight bats(Anoura caudifer,C. planirostris,Diaemus youngi,Glossophaga soricina,Micronycteris microtis, M. rufus,Pygoderma bilabiatum, andTonatia bidens) and three marsupials (Cryptonanus chacoensis,C. ignitus, andThylamys venustus) (partially shown in Tab. 3).Anoura caudiferis a species exclusive of the Yungas in Argentina, with records in the provinces of Jujuy and Salta.Diaemus youngihas been recorded in the forests of Misiones and in the Yungas of Jujuy and Salta in NWA.Glossophaga soricinahas a slightly wider distribution,covering provinces of the northeast and centre of the country. In NWA all of the records correspond to the Yungas. Until the implementation of this analysis,Micronycterismicrotishas been recorded in only one locality of the Yungas in the province of Salta.Pygoderma bilabiatumhas been recorded in the provinces of Corrientes and Misiones in the northeast,and in Jujuy and Salta in the NWA where all the records correspond to the Yungas.Tonatia bidenshas been recorded in the forests of Misiones and in the Yungas of Jujuy in NWA. Among marsupials,C. ignitusis known only by one specimen from the type locality, in the province of Jujuy, located in the Yungas.Thylamys venustusis known only from Yungas localities of the province of Salta.
3 Discussion
This study consists in a formal analysis of endemism based on distributional records of mammals,and focused on species of bats and small marsupials from northwestern Argentina. We decided to apply an optimality criterion (Szumik et al, 2002; Szumik &Goloboff, 2004). On the other hand, the parsimony analysis of endemicity (PAE) (Rosen, 1988; Morrone,1994) has been widely used to identify areas of endemism for different taxa in several regions (Escalante et al, 2009; and references therein). PAE uses a parsimony algorithm over a matrix of grid cells(equivalent to taxa in a classic phylogenetic analysis) by taxa (equivalent to characters in a classic phylogenetic analysis) in order to obtain an area cladogram. In such cladogram, clades are considered areas of endemism.Despite its wide use, PAE provide a worst resolution than NDM and allow the identification of fewer areas of endemism and fewer diagnostic species (Casagranda &Taher, 2007; Carine et al, 2009; Escalante et al, 2009).
Eighty percent of the species of small marsupials included in the analysis scored positively to the identified areas of endemism (eight -C. chacoensis,C.ignitus,Micoureus constantiae,Thylamys cinderella,T.pallidior,T. sponsorius,T. sp¸ andT. venustus- of ten species); this is in concordance with the fact that each species has a particular and restricted ecological distribution. Regarding bats,a prioriit is usually considered that they may not be considered important indicators of endemicity, because of their ability to disperse by flying. Contrary to predictions, our results show that almost 55 percent of the species of bats gave score to these areas (23 out of the 42 species). Therefore,our results show that small mammals are efficient indicators of areas of endemism; they also show that those areas are congruent with the botanical definition of the Yungas of NWA and their subdivisions, an assumption that was not previously tested with any formal quantitative method; we also found that nonvolant species are better indicators than volant at regional scale, but volant are better indicators than it was previously thought.
According to Koopman (1982), in Argentina there is a massive, relatively homogeneous zoogeographic subregion, the Patagonian subregion, which includes almost all of the bats of Argentina, with an intrusion of the Eastern Brazilian Highlands subregion into northeastern Argentina. Barquez et al (1999b) have argued that this assumption is no longer acceptable because, according to their data, it was observed that the bat fauna of Argentina is more complex than suggested,but not sufficiently rich to delineate faunal regions. Other authors also believed that bats are not of great utility in delineating faunal regions (see Willig & Mares, 1989). In contrast to these studies, our results are markedly different and somewhat similar to what it was found in other regions, for example those reported by Andersen(Andersen, 1912) who as early as in the 1910 was able to determine solid biogeographic patterns on the basis of detailed distributions of bats in the Old World Tropics.Then we suggest that bats could be considered as important faunal elements to characterize areas of endemism, together with a diversity of terrestrial small mammals.
In this study the areas of endemism were based on the points of occurrence of the species, and only records associated to museum voucher specimens, or highly reliable references, were included. It should be considered that, with respect to literature records, the identity of the species must be tentative. Also geographic biases may exist in relation to distributional points as they were not randomly recorded but distributed unevenly along roads, or in easily accessible areas. Other problems are the ecological biases associated with some aspects of the biology of the species that make it unlikely to be documented. In this sense, Ojeda et al (2003) and Tabeni et al (2004) have noted that the correct identification of priority areas for conservation may be potentially affected by the confused taxonomy of some groups, as well as by the unequal sampling effort in biomes, the obsolescence of the databases, or by changes of the landscape caused by human activities. This is also true with respect to the identification of areas of endemism. Fortunately, there are some alternatives to avoid these problems. The most commonly used alternatives are two: To apply the optimality criterion to data consisting in potential distributional models and to use the fill option in software NDM (see program documentation). Both techniques led to finding areas of endemism previously unidentified, and provided new species to diagnose the areas (Escalante et al, 2009). But both may fail because they may overpredict the distributional areas of the species. Moreover, modelling species distributions can involve the same biases as the use of points of occurrence because it makes assumptions about the primary data (uniform and random sampling, equal probability of capture of all species present at a site) that are not always true. Also, as noted by Tabeni et al (2004), using georeferenced records of occurrence optimizes the quality of analysis as opposed to the use of species distribution maps, which tend to overestimate species diversity. Biogeographic analyses based in occurrence points can be quite good despite existing some geographic and ecological biases in primary data, if a sufficiently large number of points is available. As Ojeda et al (2003) sustain, although maps of distributional areas of species are useful at larger spatial scales, at smaller spatial scales the analysis needs more detailed data. Moreover, the mammal fauna of NWA seems to be fairly well known for the total area(Ojeda et al 2003), thus allowing the implementation of reliable analysis.
In the present analysis, we have identified areas of endemism along the entire latitudinal extension of the Yungas in Argentina, as well as areas that correspond just to portions of the whole formation. This is in accordance with the pattern of latitudinal impoverishment proposed by Ojeda & Mares (1989), and in close correspondence with vegetation patterns. In particular, the Northern sector of the Yungas of NWA(sensu Brown & Ramadori, 1989) (Fig. 3C) was obtained with all the parameters (grid of cells with different shapes and sizes and with different filling values of assumed and inferred presences). Our analysis allows recognizing the Northern sector as the best defined sector because of the presence of unique species not found further south. This is not surprising because, as indicated by Ojeda & Mares (1989), the Northern sector has the higher species richness. Morales (1996) reported that the tree floras of the Central and Southern sectors are impoverished versions of the Northern sector. Our data support such impoverished pattern based on small marsupials and bats distribution, which is reflected in the nestedness of Northern areas with respect to the Yungas as a whole, as well as in the failure to recover the Southern and Central sectors of the Yungas as independent areas of endemism. The nested pattern of endemism is possible and frequent to observe. It consists in one or more areas contained within a larger one. This is the case of the Northern Yungas area of endemism.The species that give score to the Northern Yungas are different to those that give score to the larger areas of endemism that include it. All of them have distribution areas that overlap in part but do not define the same areas.For example, the nectar-feeding bats give score to the Northern Yungas area of endemism but not to the others.These tropical species occur here and no further to the south likely because of resource limitations. The southern limits ofGlossophagaandAnouramay reflect the change of degree of seasonality between the forested fragments (Ojeda & Mares, 1989). In a similar way,several other species that are found only in the North define a small area that is the one with higher diversity(Brown & Ramadori, 1989; Ojeda & Mares, 1989). In an attempt to explain this high diversity, Brown et al (2001)suggested that the Bermejo River Upper Basin in Argentina and adjacent areas in Bolivia possibly acted as a biodiversity refuge during the Pleistocene.
The main hypothesis of vicariance biogeography postulates that the emergence of barriers separate simultaneously the distribution of several taxa, producing distributional patterns that can be explained as a result of a common history. The appearance of a distributional range, with regard to continuity or discontinuity, may be only an effect of the scale of the study. A real separation requires of a real barrier to the dispersal of individuals belonging to populations separated by it (Espinosa Organista et al, 2001). For example, the location of transitional lines among biogeographic regions, has often been a reason of controversy, because of the lack of well defined climatic, environmental, or biotic barriers(current or past) among regions, and/or because different groups of organisms used to identify the barriers,respond (or responded in the past) differently to the presence of those climatic, environmental, and biotic factors. In the case of the Argentine Yungas, they are completely surrounded by semi-arid and arid environments, with very different and contrasting climatic conditions, which could be acting as very important and efficient barriers. It is well known that abrupt changes in the fauna are coincident with similar topographic, climatic, and floristic changes. In the Yungas, the most important geographic barriers could be,first of all, the different environmental conditions that limit, in elevation, the distribution of organisms; in addition, and limiting their latitudinal distribution, there are series of geographic barriers, as the great valleys,between the principal mountain chains. The intermontane valleys, with different and often contrasting climatic conditions, act as barriers for the dispersion of some species. The transitional forests, a natural connection among latitudinal sectors, have practically disappeared,contributing to accentuate the isolation between them. In addition, barriers can be of various types, not only topographical; for example Ojeda & Mares (1989)proposed that low temperatures may be acting as barriers to the dispersal of some tropical species in the Yungas of northwestern Argentina.
The establishment of protected areas requires the implementation of efficient methods that allow the selection of biodiversity hotspots, or priority areas for conservationin situ(Williams et al, 1996; Tabeni et al,2004). Hotspots are defined as those areas that individually contain the highest species richness, or the highest concentration of endemic, rare or endangered species (Williams et al, 1996; Tabeni et al, 2004). Mares(1992) pointed out that taxonomic diversity and number of endemic taxa are high in the Western Montane Forests,in which the Yungas are included. Tabeni et al (2004)found that both the methods to identify hot spots through the analysis of richness, as well as those employing the analysis of endangered species, have led to the selection of priority areas for conservation in the Yungas. Also,Ojeda et al (2003) have found that the species richest grid cells in relation to total richness, to mega-species and to endemic species, were located in the Yungas. In fact, the most important protected areas in northwestern Argentina are located in the Yungas (Fig. 1). Our results also highlight the value of the Yungas in terms of its endemic species and, if the location of the protected areas proposed by other authors (Ojeda et al, 2003;Tabeni et al, 2004) is compared with ours, the coincidence is clear. Nevertheless, additional studies are needed for the Monte and Chaco biomes (Ojeda et al,2003).
Acknowledgements:Several specimens in the Colección Mamíferos Lillo were collected in the last decades by the members of the PIDBA during field trips partially supported by grants from ANPCyT (Agencia Nacional de Promoción Científica y Tecnológica),CIUNT (Consejo de Investigaciones, Universidad Nacional de Tucumán) and CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas) with funds given to RMB. We would like to thank Michael A.Mares for assistance with the language. MLS acknowledges Norberto P. Giannini for the critical lecture of the manuscript and to CONICET for her fellowship.
Andersen K. 1912. Catalogue of the Chiroptera in the Collection of the British Museum. Volume 1: Megachiroptera [M]. 2nded. London:British Museum of Natural History.
Barquez RM. 2006. Orden chiroptera blumenbach, 1779 [M]// Barquez RM, Díaz MM, Ojeda RA. Mamíferos de Argentina, Sistemática y Distribución. Mendoza: SAREM. 56-86.
Barquez RM, Braun JK, Mares MA, Jayat JP, Flores DA. 1999a. First record for Argentina for a bat in the genusMicronycteris[J].Mammalia, 63: 368-372.
Barquez RM, DÍAZ MM. 2001. Bats of the argentine Yungas: A systematic y distributional análisis [J].Acta Zool Mex:n.s., 82: 1-81.
Barquez RM, Giannini NP, Mares MA. 1993. Guide to the Bats of Argentina [M]. Norman: Oklahoma Museum of Natural History.
Barquez RM, Mares MA, Braun JK. 1999b. The Bats of Argentina [M].Lubbock: Special Publications Texas Tech University and Oklahoma Museum of Natural History.
Brown AD. 1995a. Fitogeografía y conservación de las selvas de montaña del noroeste de Argentina [M]//Churchill SP, Balsley H,Forero E, Luteyn JL. Biodiversity and Conservation of Neotropical Montane Forests. New York: The New York Botanical Garden, 663-672.
Brown AD. 1995b. Las selvas de montaña del noroeste de Argentina:problemas ambientales e importancia de su conservación[M]//Brown AD, Grau HR. Investigación, Conservación y Desarrollo en Selvas Subtropicales de Montaña. Tucumán:Proyecto de Desarrollo Agroforestal, LIEY, 9-18.
Brown AD, Grau HR, Malizia LR, Grau A. 2001. Argentina [M] //Kappelle M, Brown AD. Bosques Nublados del Neotrópico.Heredia: INBio, 623-659.
Brown AD, y Ramadori ED. 1989. Patrón de distribución, diversidad y características ecológicas de especies arbóreas de las selvas y bosques montanos del noroeste de la Argentina[J] Anales VI Congreso Forestal Argentino, 177-181.
Cabrera AL. 1976. Regiones fitogeográficas argentinas [M]// Kugler WF.Enciclopedia Argentina de Agricultura y Jardinería,: Tomo 2. 2nded Buenos Aires: ACME Editorial,. 1-85.
Cabrera AL, Willink A. 1973. Biogeografía de América Latina [M]//Monogr. Cient. Serie Biología n 13. Washington DC: Secretaría General de la OEA.
Carine MA, Humphries CJ, Guma IR, Reyes-Betancort JA, Santos Guerra A. 2009. Areas and algorithms: Evaluating numerical approaches for the delimitation of areas of endemism in the Canary Islands archipelago [J].J Biogeogr, 36: 593-611.
Casagranda D, Taher L. 2007. Elementos bióticos, endemismo y parsimonia: estudio de casos de distribuciones hipotéticas [J].Darwiniana, 45: 37-38
Corbet GB, Hill JE. 1992. The Mammals of the Indomalayan Region:A Systematic Review [M]. Oxford: Natural History Museum Publications and Oxford University Press.
Díaz MM, Barquez RM. 2007. The wild mammals of Jujuy province,Argentina: Systematics and Distribution [M]//Kelt DA, Lessa EP,Salazar-Bravo J, Patton JL. The Quintessential Naturalist,Honoring the Life and Legacy of Oliver P. Pearson. Los Ángeles:University of California Press, 417-578.
Díaz MM, Flores DA, Barquez RM. 2002. A new species of gracile mouse opossum, genusGracilinanus(Didelphimorphia,Didelphidae), from Argentina [J].J Mammal, 83: 824-833.
Díaz Gómez JM. 2007. Endemism inLiolaemus(Iguania: Liolaemidae)from the argentinian Puna [J].South America J Herpetol, 2: 59-68.Escalante T, Szumik S, Morrone JJ. 2009. Areas of endemism of Mexican mammals: Reanalysis applying the optimality criterion[J].Biol J Linn Soc, 98: 468-478.
Espinosa Organista D, Aguilar Zuñiga C, Escalante Espinosa T. 2001.Endemismo, áreas de endemismo y regionalización biogeográfica[M]//Llorente Bousquets J, Morrone JJ. Introducción a la Biogeografía en Latinoamérica: Teorías, Métodos y Aplicaciones.México DF: Las Prensas de Ciencias, UNAM, 31-37.
Flores DA. 2003. Estudio sistemático y zoogeográfico de los marsupiales argentinos (Mammalia, Marsupialia) [D]. PhD Dissertation. Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Tucumán.
Flores DA. 2006. Orden Didelphimorphia Gill, 1872 [M]//Barquez RM,Díaz MM, Ojeda RA. Mamíferos de Argentina, Sistemática y Distribución. Mendoza: SAREM, 31-45.
Flores DA, Díaz MM, Barquez RM. 2000. Mouse opossums(Didelphimorphia, Didelphidae) of northwestern Argentina:Systematics and distribution [J].Mamm Biol, 65: 321-339.
Flores DA, Díaz MM, Barquez RM. 2007. Systematics and distribution of marsupials in Argentina: A review [M]//Kelt DA, Lessa EP,Salazar-Bravo J, Patton JL. The Quintessential Naturalist,Honoring the Life and Legacy of Oliver P. Pearson. Los Ángeles:University of California Press, 579-669.
Goloboff P. 2005. NDM/VNDM v.2.5. Programs for identification of areas of endemism. Programs and documentation avalaible[EB/OL]. www.zmuc.dk/public/phylogeny/endemism.
Harold AS, Moi RD. 1994. Areas of endemism: Definition and recognition criteria [J].Syst. Biol, 43: 261-266.
Hortal J, Roura-Pascual N, Sanders NJ, Rahbek C. 2010.Understanding (insect) species distributions across spatial scales[J].Ecography, 33: 51-53.
Hueck K. 1978. Los Bosques de Sudamérica. Ecología, Composición e Importancia Económica [M]. Berlin: Soc. Alemana de Cooperación Técnica (GTZ).
Jayat JP, Miotti MD. 2006. Primer registro deMonodelphis kunsi(Didelphimorphia, Didelphidae) para Argentina [J].Mast Neotr,12: 253-256.
Koopman KF. 1976. Zoogeography [M]//Baker RJ, Jones JK, Jr.,Carter DC. Biology of Bats of the New World Family Phyllostomidae: Part 1. Lubock: Special Publications, the Museum, Texas Tech University, 10. 39-47.
Koopman KF. 1981. The distributional patterns of new world nectarfeeding bats [J].Ann Missouri Bot Gard, 48: 352-369.
Koopman KF. 1982. Biogeography of bats of South America[M]//Mares MA, Genoways HH. Mammalian Biology in South America. Pennsylvania: Special Publications Series, Pymatuning Laboratory of Ecology, University of Pittsburgh, 273-302.
Levin SA. 1992. The problem of pattern and scale in ecology: The Robert H. MacArthur award lecture [J].Ecology, 73: 1943-1967.
Linder HP. 1995. Setting conservation priorities: The importance of endemism and phylogeny in the southern african orchid genusHerschelia[J].Conserv Biol, 9: 585:595.
Linder HP. 2001. On areas of endemism, with an example from the african Restionaceae [J].Syst Biol, 50: 892-912.
Mares MA. 1992. Neotropical mammals and the myth of Amazonian biodiversity [J].Science, 255: 976-979.
Meyer T. 1963. Estudios sobre la selva tucumana. La selva de mirtáceas de Las Pavas [J].Oper Lilloana, 10: 1-144.
Morales JM. 1996. Comunidades arbóreas no saturadas en las Yungas de Argentina [J].Ecol Aust, 6: 17-22.
Morales JM, Sirombra M, Brown AD. 1995. Riqueza de árboles en las Yungas argentinas [M]//Brown AD, Grau HR. Investigación,Conservación y Desarrollo en Selvas Subtropicales de Montaña.Tucumán: Proyecto de Desarrollo agroforestal, LIEY, 163-174.
Morrone JJ. 1994. On the identification of areas of endemism [J].Syst Biol, 43: 438-441.
Morrone JJ. 2000. A new regional biogeography of the Amazonian subregion mainly based on animal taxa [J].An Inst Biol Univ Nac Auton Mex:Ser Zool, 71: 99-123.
Morrone JJ. 2001. Biogeografía de América Latina y El Caribe:Volume 3. [M]. Zaragoza: M&T–Manuales & Tesis SEA.
Ojeda RA. 1999. Biodiversidad y conservación de mamíferos de la interfase tropical-templada de la Argentina [M]//Matteucci SD,Solbrig OT, Morillo J, Halffter G. Biodiversidad y Uso de la Tierra: Conceptos y Ejemplos de Latinoamérica. Buenos Aires:
Eudeba, Universidad de Buenos Aires, 443-462.
Ojeda RA, Barquez RM, Stadler J, Brandl R. 2008. Decline of mammal species diversity along the Yungas forest of Argentina [J].Biotropica, 40: 515-521.
Ojeda RA, Mares MA. 1989. A Biogeographic Analysis of the Mammals of Salta Province, Argentina. Patterns of Species Assemblage in the Neotropics [M]. Lubbock: Special Publications,the Museum, Texas Tech University 27. 1-66.
Ojeda RA, Stadler J, Brandl R. 2003. Diversity of mammals in the tropical-temperate Neotropics: Hotspots on a regional scale [J].Biodiver Conserv, 12: 1431-1444.
Peterson AT, Watson DM. 1998. Problems with areal definitions of endemism: The effects of spatial scaling [J].Divers Distribut, 4:189-194.
Platnick N. 1991. On areas of endemism [J].Aust Syst Bot, 4: 11-12.
Prado DE. 1995. Selva pedemontana: contexto regional y lista florística de un ecosistema en peligro [M]//Brown AD, Grau HR.Investigación, Conservación y Desarrollo en Selvas Subtropicales de Montaña. Tucumán: Proyecto de Desarrollo agroforestal, LIEY,19-52.
Proches S. 2005. The world's biogeographical regions: Cluster analyses based on bat distributions [J].J Biogeogr, 32: 607-614.
Ribichich AM. 2002. El modelo clásico de la fitogeografía argentina:un análisis crítico [J].Interciencia, 27: 669-675.
Rosen BR. 1988. From fossils to earth history: Applied historical biogeography [M]//Myers AA, Giller P. Analytical Biogeography:An Integrated Approach to the Study of Animal and Plant Distributions. London: Chapman and Hall, 437-481.
Ruggiero A, Ezcurra C. 2003. Regiones y transiciones biogeográficas:complementariedad de los análisis en biogeografía histórica y ecológica [M]//Morrone JJ, Llorente Bousquets J. Una Perspectiva Latinoamericana de la Biogeografía. México: UNAM, 141-154.
Szumik C, Cuezzo F, Goloboff P, Chalup A. 2002. An optimality criterion to determine areas of endemism [J].Syst Biol, 51: 806-816.
Szumik C, Goloboff P. 2004. Areas of endemism. An improved optimality criterion [J].Syst Biol, 53: 968-977.
Szumik C, Roig-Juñent S. 2005. Criterio de optimalidad para áreas de endemismo: el caso de América del Sur Austral [M]//Morrone JJ,Llorente Bousquets J. Regionalización Biogeográfica en Iberoamérica y Tópicos Afines. México,;UNAM, 495-508.
Tabeni MS, Bender JB, Ojeda RA. 2004. Puntos calientes para la conservación de mamíferos en la provincia de Tucumán,Argentina [J].Mast Neotr, 11: 55-67.
Vane-Wright R, Humphries CJ, Williams PH. 1991. What to protectsystematics and the agony of choice [J].Conserv Biol, 55: 235-254.
Vervoorst F. 1979. La vegetación del noroeste argentino y su degradación [M]. Serie Conservación de la Naturaleza. Tucumán:Fundación Miguel Lillo.
Williams P, Gibbons D, Margules C, Rebelo A, Humphries C, Pressey R. 1996. A comparison of richness hotspots, rarity hotspots, and complementary areas for conserving diversity of British birds [J].Conserv Biol, 10: 155-174.
Willig MR, Mares MA. 1989. A comparison of bat assemblages from phytogeographic zones of Venezuela [M] // Morris DW,Abramsky Z, Fox BJ, Willig MR. Patterns in the Structure of Mammalian Communities. Lubbock: Spec. Publ., The Museum,Texas Tech University, 28. 59-67.
阿根廷Yungas雨林特有翼手类和有袋类
María L Sandoval1, Claudia A Szumik2, Rubén M Barquez1,*
[1.PIDBA(Research Program of Argentinean Biodiversity),Natural Sciences Faculty and Miguel Lillo Institute, National University of Tucuman, Miguel Lillo205,San Miguel de Tucumán, CP4000,Tucumán, Argentina; andCONICET(National Council of Scientific and Technical Researches),Argentina; 2.INSUE(Superior Institute of Entomology“Dr. Abraham Willink”),Natural Sciences Faculty and Miguel Lillo Institute, National
University of Tucuman, Miguel Lillo205,San Miguel de Tucumán, CP4000,Tucumán, Argentina; andCONICET(National Council of Scientific and Technical Researches),Argentina]
有学者根据植物组成等特性将 Yungas雨林定性为独立的生物地理单元,但这些特性主要是定性的且未包括其动物区系组成。Yungas雨林被认为是动植物种类分布丰富的多样性的区域。然而,尚未有研究评估其动物分布记述的状况。在生物地理学上 Yungas雨林使人充满兴趣,它函盖不连续分布的雨林且延伸至温带干旱和半干旱地带。该研究分析了 Yungas最南端地区小型哺乳动物特有种分布记录,对比了飞行物种(蝙蝠类)和非飞行物种(有袋类)的分布。结果显示小型哺乳动物特有种是 Yungas地区的有效指标;所分析的 80%小型有袋类物种和 55%蝙蝠类物种支持所鉴定的区域作为特有种区域。所研究的区域与阿根廷西北部及其以下的Yungas的植物学定义一致。该区域以前尚未正式用定量方法评测。结果还发现非飞行特有种较之飞行特有种更合适作为区域尺度上的特有种指标,作者认为飞行特物种作为特有种指标比以前所认为的要更好。
Andean雨林;翼手目;负鼠目;最优性准则;小型哺乳动物
Q958.155;Q959.822;Q959.833
A
0254-5853-(2010)06-0633-012
;2010-05-05;接受日期:2010-10-25
10.3724/SP.J.1141.2010.06633
date: 2010-05-05; Accepted date: 2010-10-25
*Corresponding author (通信作者), Email: rubenbarquez@arnet.com.ar
猜你喜欢
杂志排行

Zoological Research的其它文章
- Shape change in viable eggs of the collembolan Folsomia candida provides insight into the role of Wolbachia endosymbionts
- Digestive enzyme and alkaline phosphatase activities during the early stages of Silurus soldatovi development
- Isolation and characterization of Hainantoxin-II, a new neurotoxic peptide from the Chinese bird spider (Haplopelma hainanum)
- Morphological changes of silver and bighead carp in the Yangtze River over the past 50 years
- Positive influence of traditional culture and socioeconomic activity on conservation: A case study from the black-and-white snub-nosed monkey (Rhinopithecus bieti) in Tibet
- A new record of Dasyatid fish in China: Dasyatis laosensis