紫色光合细菌Thermochromatium Tepidum捕光天线复合物2的激发态动力学
2010-12-12于龙江艾希成王征宇张建平
杨 帆 于龙江 王 鹏 艾希成 王征宇 张建平,*
(1集成光电子学国家重点联合实验室吉林大学实验区,吉林大学电子科学与工程学院,长春 130012; 2中国人民大学化学系,北京 100872; 3茨城大学理学部,水户市文京2-1-1 310-8512,日本)
The discovery of the high-resolution crystallographic structures of the light-harvesting complex 2(LH2)from photosynthetic purple bacteria Rhodopseudomonas(Rps.)acidophila[1-2]and Rhodospirillum(Rs.)molischianum[3]has stimulated extensive experimental and theoretical investigations aimed at its structure-function relationship.In the photosynthetic membrane of purple bacteria,a core light-harvesting complex 1(LH1)encircling a photosynthetic reaction center(RC)is surrounded by a pool of peripheral LH2s(see Ref.[4]and references therein). Light energy harvested by the antenna complexes(LH1,LH2)is transferred in form of electronic excitation to the RC with extremely high efficiency(>90%)to initiate the primary charge separation[5].The pigment-protein assembly of LH2 consists of 8 and 9repeatingsubunitsforRs.molischianum[3]andRps.acidophila[1], respectively.In the LH2 complexes,the subunits form circular aggregates with 8-or 9-fold symmetry,each subunit consists of a pair of α and β trans-membrane polypeptides,which bind 2 bacteriochlorophyll a(BChl)molecules with Qyabsorption at 850 nm(B850),1 BChl at 800 nm(B800),as well as 1 carotenoid (Car)molecule with slightly twisted all-trans configuration that is sandwiched between the pair of B850s and passes by the B800.The light-harvesting function of bacterial antenna is based on a series of ultrafast excitation energy transfer(EET)reactions proceeding in the time scales of 0.1-50 ps and in a cascading and downhill manner,e.g.,Car-to-BChl and B800-to-B850 within a LH2 complex[6],LH2-to-LH1 and LH1-to-RC[7].
For the aforementioned LH2 complexes with known crystallographic structures,the molecular and electronic structures and the excited-state properties of BChls and Cars have been examined in great details(see Refs.[6-9]for reviews).Differing from the pigment-protein assemblies of these LH2s consisting of relatively simple constituents of trans-membrane polypeptides and pigment cofactors,the LH2 complexes from other bacterial species,such as Rps.palustris and Thermochromatium(Tch.) tepidum,contain multi-composition polypeptides and/or Cars. Recently,the LH2 complex from Rps.palustris has been identified with a 0.75 nm resolution to be an 8-fold symmetric octamer,and each α,β-subunit most likely contains 4 BChls with unique structural orientation and spectroscopic properties[10].In addition,the organization and the macroscopic structures of bacterial antenna complexes in intracytoplasmic membrane(ICM) has recently been revealed by the use of atomic force microscopy[11-12].Despite these advances,high-resolution crystallographic structures remain unknown for the LH2s with relatively complicated constituents of α,β-polypeptides and/or pigments. Accordingly,the excitation dynamics of these LH2s,such as those from Rps.palustris or Tch.tepidum etc.,have not been investigated in details.
Purple photosynthetic bacterium Tch.tepidum is a moderate thermophile growing in an optimal temperature range of 48-50℃,which was firstly found from Mammoth Hot Springs in the Yellowstone National Park[13].Its pigment-protein complexes exhibit notably higher thermal stability than the mesophilic counterparts such as Allochromatium (Ach.)vinosum and Rb.sphaeroides growing under~30℃[14].We have recently studied the excited-state dynamics,and the origins of the long-wavelength Qyabsorption and the thermal stability of the LH1-RC core complex from Tch.tepidum[15-16].The LH2 of Tch.tepidum,whose crystallographic structure is unknown,is different from those of Rps.acidophila or Rs.molischianum in apoprotein and pigment compositions.It contains three different types of α and β polypeptide pairs[17],as well as three major carotenoid compositions categorized by the number of C=C conjugated double bonds(NC=C),namely,rhodopin and lycopene(NC=C=11),anhydrorhodovibrin (NC=C=12),and spirilloxanthin and(OH)-spirilloxanthin(NC=C=13)[18].
This work is intended to examine the detailed EET dynamics of the isolated LH2 complex from Tch.tepidum.Surfactants with different ionic properties,i.e.,nonionic n-dodecyl-β-D-maltoside(DDM)and zwitterionic lauryldimethylamine oxide (LDAO),are used to prepare the LH2 complex,and the ultrafast EET processes,including Car-to-Car,Car-to-BChl,and BChl-to-BChl,are examined.The implications of the excitation dynamics are discussed in terms of the light-harvesting and the photoprotection roles of the LH2 complex.
1 Materials and methods
1.1 Sample preparation
Tch.tepidum was cultured anaerobically at 48℃for 7 d.The cells were disrupted at 4℃by ultrasonication,and the obtained chromatophores were suspended in 20 mmol·L-1Tris-HCl buffer(pH 8.5)at a concentration of OD850nm~50 cm-1(OD:optical density).After being solubilized with 0.35%(φ,volume fraction)LDAO(Kao Corp.,Japan)for 60 min in the dark,the suspension was centrifuged(145400×g,4℃,100 min),and the supernatant was collected as crude LH2.The crude LH2 preparation was further purified by using DEAE-cellulose(Whatman DE52)column chromatography in the presence of 0.05%LDAO or 0.05%DDM,for the LDAO or the DDM preparations.The UV-visible absorption spectra were recorded on a Cray 50 absorption spectrometer(Varian).Fluorescence spectra were measured on a LS-55 luminescence spectrophotometer(Perkin Elmer).
1.2 Femtosecond time-resolved spectroscopy
The femtosecond time-resolved absorption apparatus with a temporal resolution of~150 fs is similar to that described in Refs.[15,19].Briefly,an optical parametric amplifier(OPA-800 CF-1,Spectra Physics)pumped bya regenerative amplifier(SPTF-100F-1KHPR,Spectra Physics)provided the actinic laser pulses (~120 fs,full width at half maximum),which was sent to the sample cell(optical path length,1 mm)with an excitation photon densities of 1013-1014photons·cm-2·pulse-1.White light continuum probe was generated from a 3-mm thick sapphire plate, and was detected after interrogating the excited sample by a liquid-nitrogen cooled CCD detector(Spec-10:400B/LN)attached to an imaging spectrograph(SpectraPro 2300i,USA).Timeresolved spectra were corrected against group velocity dispersion.To ensure that each laser shot excites fresh sample,the laser system was ran at the repetition rate of 100 Hz,and the sample cell was kept on shifting back and forth across the overlapped actinic and probe beams.The optical density of a LH2 sample at an actinic wavelength was adjusted to 0.2-0.45 mm-1. All of the time-resolved measurements were carried out at room temperature(23℃).Computer programs for the kinetics analysis were compiled based on Matlab (Mathworks)and Mathcad (MathSoft).
2 Results and discussion
2.1 Steady-state electronic absorption of the LH2 preparations

Fig.1 UV-visible electronic absorption spectra of the DDM (red)and the DDM(black)preparations of isolated LH2 complexes from Tch.tepidumDashed lines are the corresponding fluorescence excitation spectra recorded at the observing wavelength of 900 nm.Spectra are normalized to the Qx absorption maximum at 590 nm.Arrows point to the excitation wavelengths for time-resolved spectroscopy.
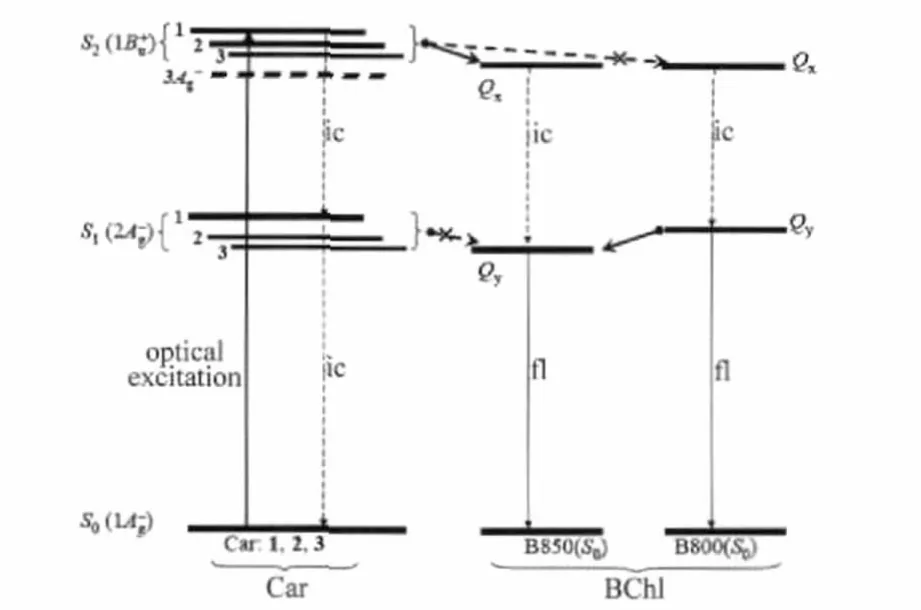
Fig.2 Schematic excited-state energy diagram and singlet excitation energy transfer scheme of the isolated LH2 complex form Tch.tepidumNumerals 1,2,and 3 represent the Car compositions 1,2 and 3 having NC=Cvalues of 11,12,and 13,respectively.The relative position of the exited-state manifolds of Car and BChl are scaled according to their state energies[8]. The recently found intermediate state,3A-g,of anhydrorhodovibrin[21] (composition 2)is also shown.Vertical arrows represent the optical excitation and the relaxation processes,i.e.,internal conversion(ic)and fluorescence emission(fl). Horizontal arrows indicate the EET processes discussed in this paper.
The broadband absorption covering 450-570 nm in Fig.1 originates from the strongly optically allowedtransition of Car(Fig.2).The Car absorption spectrum of the LDAO preparation shows no significant difference from that of the DDM preparation except the slightly more prominent vibronic structures.It is documented that the absorptive transitionis rather susceptible to the Car surroundings[20]. Therefore,the insignificant change of Car absorption spectra suggests that Cars in LH2 experience only subtle environmental changes from the DDM preparation to the LDAO one.
The LH2 complex of Tch.tepidum contains three major carotenoid compositions:composition 1 includes rhodopin (66.5%)and lycopene(2.2%)with NC=C=11;composition 2 is anhydrorhodovibrin(8.7%)with NC=C=12;composition 3 consists of spirilloxanthin(20.4%)and OH-spirilloxanthin(4.4%) with NC=C=13.It is well known that,theabsorption of Car shifts systematically to longer wavelength upon increasing NC=C.Specifically,the transition energy holds a linear relationship with 1/(2NC=C+1)[21-22].Therefore,the absorption spectra of different Car compositions overlap each other,which blurs the vibronic structures with respect to the LH2 containing single type of Car,e.g.,that of Rb.sphaeroides 2.4.1 contains spheroidene(NC=C=10)[23].By the use of fluorescence excitation spectroscopy,we have determined the overall Car-to-BChl singlet energy transfer efficiency,<30%,which is in general agreement with those reported for Cars having NC=Cvalues of 11-13 in bacterial light-harvesting complexes[24].Here,we note that for the LH2 complex containing multi-composition Cars,it is presently unknown that whether the Car heterogeneity associates to an individual LH2 or to different LH2s,an intriguing issue remaining to be clarified.
In the near-infrared region of Qyabsorption(Fig.1,750-900 nm),the spectrum of the DDM preparation resembles the LH2 spectra of Rs.molischianum,Rps.acidophila,or Rb.sphaeroides 2.4.1 both in maximal wavelength(~850 nm)and in B850-to-B800 absorption ratio(A850/A800=1.2-1.7).However,the spec-trum of the LDAO preparation differs significantly from that of the DDM preparation,i.e.,the B850(Qy)band blue shits for~6 nm,and the A850/A800ratio decreases to 0.9 with reference to that of the DDM preparation,1.5.The spectral variation is apparently induced by the detergents:For the specific case of Tch. tepidum,the nonionic DDM is gentle and,therefore,facilitates preserving the native pigment-protein assembly of LH2,whereas the zwitterionic LDAO is relatively harsh,and hence induces subtle structural change.On the other hand,the excitonic interaction among the B850 molecules is very sensitive to the geometry of the B850 circular aggregate.Furthermore,specific and nonspecific interactions between B850 and its surroundings also largely influence the Qyabsorption[25-27].Therefore,even subtle structural variation of the LH2 assembly can induce appreciable spectral change of the B850(Qy)band.At the present stage,for the LH2 complex of Tch.tepidum lacking of structural details,a complete theoretical account for the electronic absorption spectrum seems unfeasible.
2.2 Car-to-BChl excitation energy transfer
2.2.1 Excited state dynamics
Selective excitation of Car allows the processes of Car-to-BChl EET to be followed by probing the spectral dynamics and the population kinetics.As seen in Fig.3(a)for the DDM preparation,immediately following the pulsed excitation(0.0 ps),bleaching of Car ground state absorption appears as a negative signal (450-570 nm).The Car bleaching is accompanied by the characteristic Sn←S1absorption to the longer wavelength side(570-650 nm),which subsequently reaches the maximum at 0.3 ps. This rise phase of excited state absorption is due to the S2-to-S1internal conversion(ic,time constant derived by fitting the 600 nm kinetics in Fig.3(b),~140 fs).Importantly,the Sn←S1absorption maximized at 600 nm is attributed to anhydrorhodovibrin (NC=C=12)judging from its transition energy[28].
In the near-infrared region(Fig.3(a),800-1000 nm),the exciton absorption(800-840 nm)together with the B850 bleaching (840-950 nm)are observed,both of which are mainly induced by the ultrafast Car(S2)-to-B850(Qx)EET.Note that,at 0.0 ps the B850(Qy)exciton absorption does not appear despite a sizable B850 bleaching.The delay of exciton absorption is explained below:the ultrafast Car(S2)-to-B850(Qx)EET promptly depopulates the ground state B850 and,subsequently,the B850(Qy) population is built up via the ic process of Qx-to-Qy(Fig.2).Thus the rise of B850(Qy)exciton absorption takes place with a time constant comparable to that of the Qx-to-Qyic,~220 fs,as derived from the 830 nm kinetics in Fig.3(b),in well agreement with that determined for the LH2 complex of Rb.sphaeroides G1C,~300fs[29].Inthesamescenario,theabsence ofB800 bleaching in Fig.3(a)proves that the Car-to-B800 EET path is inactive, which is most likely due to the unfavorable orientation of the transition dipole moments between Car and B800.

Fig.3 (a,c)Femtosecond time-resolved spectra at indicated delay times,and(b,d)the corresponding kinetics at indicated probe wavelengths for the DDM(a,b)and the LDAO(c,d)preparations of LH2 complexes from Tch.tepidumSolid lines in(b,d)are fitting curves.Excitation wavelength was 530 nm.
2.2.2 Ultrafast formation of triplet excited state Car
From the decaying phase of the spectral dynamics(Fig.3(a), 0.3 ps→20.0 ps),it is seen that,when the Sn←S1absorption decays out at 20 ps,an absorption band at~565 nm remains.This transient with a decay time constant of microseconds,as derived from the 565 nm kinetics in Fig.3(b),can be safely attributed to the Tn←T1absorption of Car.Note that the Tn←T1transients appears in subpicosecond delay times,which cannot be explained by the BChl-to-Car triplet EET proceeding on a time scale of~10 ns as governed by Dexter′s electron-exchange mechanism[30]. The ultrafast formation of3Car*in subpicosecond time scale had been previously observed for the LH complex from Rs.rubrum containing the long chain Car,spirilloxanthin(NC=C=13)[31]. Since the covalent 21A-gexcitation intrinsically consists of a pair of double-excited and spin-correlated triplet excitationsthe ultrafast3Car*formation is ascribed to the singlet homofission reaction within Car conjugated backbone,i.e.,CarJudging from the Tn←T1transition energy,this triplet absorption spectrum is best attributed to the Car having NC=C=12(composition 2,anhydrorhodovibrin). Similar observation has recently been reported for the LH2 from Rps.palustris[32].
2.2.3 Car-to-BChl singlet excitation energy transfer
Both the S1and the S2states of Car are capable of mediating the Car-to-BChl singlet EET,e.g.,the LH2 complex of Rb. sphaeroides G1C containing a singlet Car composition of neurosporene(NC=C=9)shows an overall Car-to-BChl EET efficiency of 95%,the partitions of efficiency are 60%-74%for the Car (S2)-to-BChl(Qx)path and 24%-38%for Car(S1)-to-BChl(Qy), and the corresponding EET time constants are~100 fs and~1.42 ps,respectively[29].For energetic reasons,the Car(S1)-to-BChl (Qy)path is rather inefficient for Cars having NC=C=11,and it is even closed for those having NC=C=12 or 13.However,the Car (S2)-to-BChl(Qx)path is valid for most of the photosynthetic Cars(NC=C=9-13)in bacterial antenna complexes[28,33].In the LH2 complex of Tch.tepidum,the Car-to-BChl EET is predominantly mediated by the S2state,whereas the Car(S1)-to-BChl(Qy) path is inactive.This conclusion drawn on the basis of spectral dynamics is also supported by the population kinetics:(i)the S1-state lifetime of anhydrorhodovibrin in n-hexane(2.2 ps[34])in absence of energy acceptor is not shortened compared to that of the same Car bound in LH2(~2.4 ps,derived from the 600 nm kinetics in Fig.3(b));(ii)from the B850 bleaching kinetics at 860 nm,only a rise phase in a time scale of~100 fs is derived,i.e., no picosecond rising component correlating to the S1-state depopulation could be identified.
The spectral dynamics and population kinetics in Fig.3(c,d) for the LDAO preparation are similar to those of the DDM preparation.An apparent difference is seen by comparing the decay phases in Fig.3(a)and Fig.3(c):the rapid decay phase of the B850 bleaching for the DDM preparation is not seen for the LDAO preparation.This rapid decay with a time constant of~270 fs,as derived from the 860 nm kinetics in Fig.3(b),is due to singlet annihilation among the B850 excitations.
Taken together the above ultrafast spectroscopic results,we propose the scheme of Car-to-BChl EET as illustrated in Fig.2 for the LH2 of Tch.tepidum.It is important to point out that,despite the possible structural variation between the DDM and the LDAO preparations,the Car-to-BChl EET schemes are similar, suggesting that the accessory light-harvesting function of Car is rather robust against the structural variation of LH2.
2.3 Car-to-Car excitation energy transfer
Up to now,the subject of Car-to-Car singlet EET in bacterial light-harvesting complex has not been examined in any details, although Car-to-Car triplet EET at cryogenic temperature has recently been suggested for the LH2 of Rps.palustris[35].The difficulty of spectroscopic detection of Car-to-Car EET process stems from the fact that Car molecules in LH2 are identical and, therefore,hardly to be spectroscopically differentiated.The multicomposition Cars in the LH2 complex of Tch.tepidum may provide a possibility to tackle this problem.
2.3.1 Excited state dynamics
Despite sever overlap among the ground state absorption of different Cars(Fig.1),it is expect that excitation to the shorter wavelength side of the Car absorption band can preferentially excite shorter chain Cars owing to their highertransition energy.Fig.4 shows the representative transient spectra recorded under different excitation wavelengths.(i)Δt=0.0 ps. The ΔOD spectra show negative Car bleaching accompanied by the broadband Sn←S1absorption to the longer wavelength side, and no essential difference is seen from the transients at different excitation wavelengths.(ii)Δt=0.5 ps.The Sn←S1absorption reaches the maxima as a result of S2-to-S1ic proceeding in a time scale of~140 fs.For the LDAO preparation(Fig.4(b)),the shoulder absorption peaked at~570 nm becomes more prominent for longer excitation wavelength.This is an indication of Tn←T1absorption associated to the ultrafast3Car*formation.Such excitation wavelength dependence is not obvious for the DDM preparation.(iii)Δt=10.0 ps.The Tn←T1absorption peaked at~560 nm is observed irrespective to the excitation wavelengths.
2.3.2 Inactivity of Car(T1)-to-Car(T1)EET
Interestingly,although all of the three Car compositions were excited,only the Tn←T1absorption of anhydrorhodovibrin is observed(Figs.4(a,b),Δt=10.0 ps).Since direct Car-to-Car triplet EET must be extremely inefficient owing to the large intermolecular distance of~1 nm as known from the crystallographic structure of Rps.acidophila[1],the Tn←T1absorption of rhodopin, lycopene,or spirilloxanthin,if any,would be observed at different transition wavelengths.Here,it is difficult to understand why only the Tn←T1absorption of anhydrorhodovibrin is observed. Besides possible structural origins awaiting for detailed LH2 structural information of Tch.tepidum,we propose the following mechanism based on the unique electronic structure of this particular Car.Recent resonance Raman excitation profile studies of Car has revealed the presence of thestate beneath the well-knownstate[21](Fig.2).Theenergy separa-tion,~1700 cm-1,is comparable to a vibrational quanta of C=C double bond stretching,therefore,theic via state mixing must be extremely efficient.Importantly,because thestate is~500 cm-1below the Qxstate,directsinglet EET is energetically unfavorable.Taken together,the EET path of Car()-to-BChl(Qx)must be very inefficient for anhydrorhodovibrin owing to the rapid deactivation of theexcitation via efficientic.As the result,thepopulation is predominantly converted via cascading ic processes to theand further to thestate,which eventually show up as the Sn←S1and the Tn←T1absorption,respectively.

Fig.4 Comparison of the spectral dynamics of the LH2 complexes following selective optical excitation of Car at 475,500 and 530 nm(a)DDM preparation,(b)LDAO preparation.The spectra at delay times(Δt)of 0.5 and 10.0 ps are normalized.
2.3.3 Inactivity of Car(S1)-to-Car(S1)EET
As shown in Fig.5(a,b),at the probing wavelength of 600 nm,population kinetics under the excitation wavelengths of 475 and 530 nm shows almost identical decay time constant of (2.44±0.03)and(2.63±0.02)ps for the DDM and LDAO preparation,respectively.Since 475 nm laser pulses preferentially excite shorter chain Cars(NC=C=11)having a longer S1-state lifetime of~4 ps,the associated Sn←S1absorption,if any,would appear at the characteristic maximal wavelength of 580 nm[23].The above kinetics similarity implies that the EET path of Car(S1)-to-Car(S1) is inactive,which may be understood in view of the followings. (i)Because of the large Car-Car intermolecular distance(~1 nm)and the rather small transition dipole moment of the S1state (<3.33×10-30C·m)[33],the strength of transition dipole coupling between a pair of Cars must be extremely low and,consequently,the S1-state mediated Car-to-Car EET is extremely inefficient. (ii)The ultrafast Car(S2)-to-B850(Qx)EET proceeding on~100 fs together with the ultrafast Car(S2)-to-Car(S2)EET(vide infra) are competitive to the S2→S1ic process(~140 fs),thus population to the S1state of shorter chain Cars(NC=C=11)must be inefficient.
2.3.4 Car(S2)-to-Car(S2)EET
Fluorescence excitation spectroscopy has revealed that,for either DDM or LDAO preparations,the overall efficiency of Carto-BChl EET at 475 nm(~11%)is much lower than that at 530 nm(~28%),which are in contrast to the general trend that the EET efficiency is higher for shorter chain Cars.In addition,only the Sn←S1and the Tn←T1absorption of anhydrorhodovibrin (NC=C=12)are observed despite the much higher composition of the Cars with NC=C=11(~67%)compared to that of anhydrorhodovibrin(~8%).These observations strongly suggest the presence of S2-state mediated EET from the shorter chain rhodopin and/or lycopene to the longer chain anhydrorhodovibrin,which is further supported by the kinetics results:at the probing wavelength of 560 nm,the relative amplitude of the3Car*species (anhydrorhodovibrin)is higher under shorter wavelength excitation at 475 nm(Fig.5(a,b)).Under the 475 nm excitation,direct excitation of anhydrorhodovibrin is expected to be less probable,however,an unusually high population of3Car*(anhydrorhodovibrin)is observed,and those of shorter chain Cars(rhodopin and lycopene)expected to appear at short-er transition wavelengths are not seen.This is readily accounted for by the S2-state mediated ultrafast Car-to-Car EET,which together with the ultrafast Car(S2)-to-BChl(Qx)EET efficiently deplete the S2-state and hence the S1-state population of Cars (NC=C=11).The presence of ultrafast Car(S2)-to-Car(S2)EET also holds the structural basis:thetransition carries a transition dipole moment as large as 5.00×10-30C·m[33].In addition,the Car back-bones are nearly paralleling as known from the existing crystallographic LH2 structures.Therefore,strong transition dipole coupling between Cars is expected in spite of the~1 nm separation.It is then in-ferred from the Car(S2)-to-Car (S2)EET that Cars having NC=C=11 and NC=C=12 coexist in a LH2 complex.Here,we note that,the minor Car composition anhydrorhodovibrin in LH2 seems an efficient trap of excitation energy rather than a light harvester.
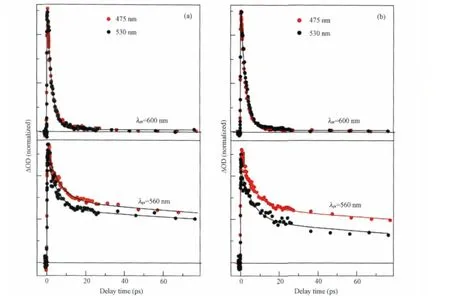
Fig.5 Comparison of the population kinetics under the indicated excitation wavelengths for (a)DDM and(b)LDAO preparations of LH2 complexes from Tch.tepidumThe kinetics are plotted from the spectral dynamics shown in Fig.4 at the indicated probing wavelengths(λpr).Solid lines are fitting curves.
2.4 B800-to-B850 excitation energy transfer
Fig.6 shows the bleaching recovery kinetics of B800 and B850 probed at 810 and 860 nm,respectively,as well as the kinetics of B850(Qy)exciton absorption probed at 830 nm.The instantaneous bleaching of B800 induced by pulsed excitation is followed by a slow recovery phase,which closely correlates to the rise phases of the kinetics at 830 or 860 nm.Such decay-to-rise correlation substantiates the process of B800(Qy)-to-B850 (Qy)EET(Fig.2).
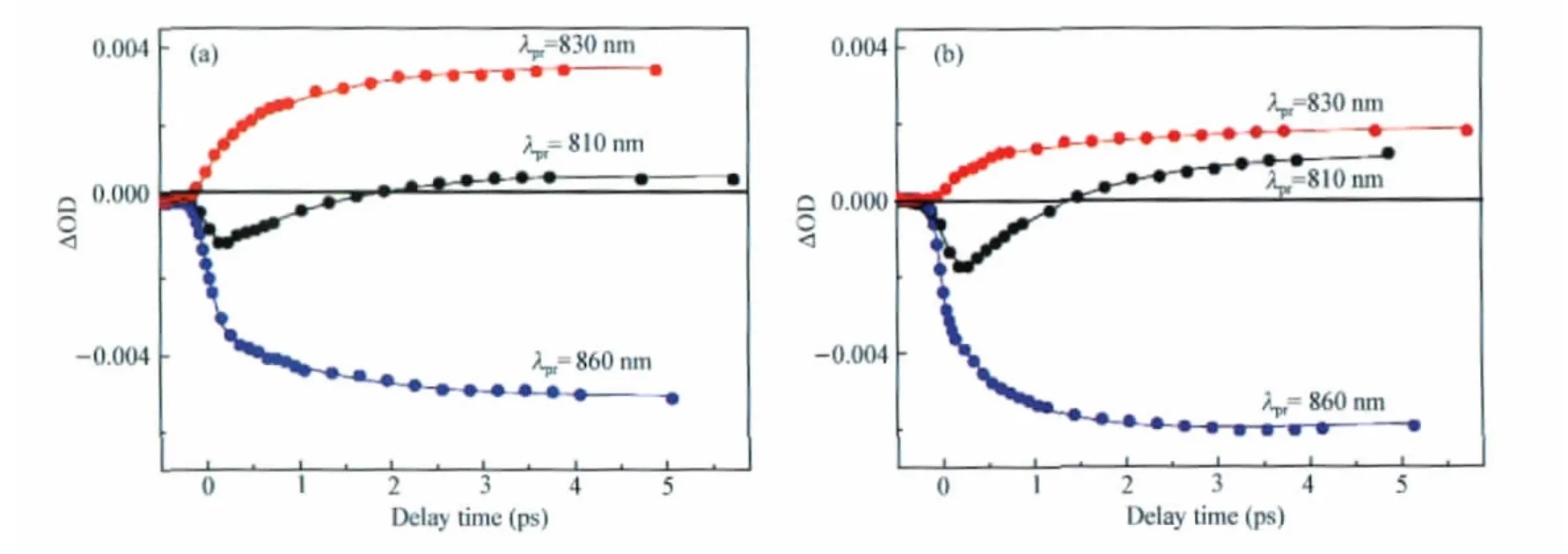
Fig.6 Kinetics at the indicated probing wavelengths for(a)DDM and(b)LDAO preparations of LH2 complexes from Tch.tepidumSolid lines are fitting curves.Excitation wavelength was 785 nm.
Global fit of the three kinetics traces with bi-or tri-exponential model functions revealed the EET time constants,i.e.,~1.2 ps for the DDM preparation and~1.7 ps for the LDAO one.The value of 1.2 ps for the DDM preparation is~50%prolonged compared to the B800(Qy)-to-B850(Qy)EET time constants for the LH2s containing single Car composition from Rps.acidophila,Rs.molischianum,and Rb.sphaeroides(~0.8 ps)[36],suggesting that the mutual orientation of B800 and B850 in the LH2 of Tch.tepidum differs considerably from those in the LH2s from the above mesophilic species.Fitting the individual kinetics at 830 and 860 nm yielded identical rise time constant,i.e.,~1.2 ps for the DDM preparation and~0.8 ps for the LDAO one, whereas fitting the specific traces at 810 nm yielded a decay time constant of~1.1 ps for either DDM or LDAO preparation. Thus the B800(Qy)-to-B850(Qy)EET time constants for the DDM preparation derived from different fitting procedures are the same and,therefore,the value of~1.2 ps is considered to be robust.Here we note that the B800(Qy)-to-B850(Qy)EET time constant for the LDAO preparation of LH2 from Tch.tepidum had previously been reported to be 0.7-0.9 ps[37].How-ever,the detergent condition for LH2 preparation in Ref.[37]differs significantly from the present case.Nonetheless,the considerable difference in the time constants of B800(Qy)-to-B850(Qy)EET between the DDM and the LDAO preparations sug-gests some change in the mutual orientation of B800 and B850.Despite the relatively slow B800(Qy)-to-B850(Qy)EET,the EET efficiency of either of the LH2 preparations is close to unity because of the~1 ns Qy-state lifetime of BChl in LH2[38].These results imply that the light-harvesting functionality of LH2 is rather robust against its structural variation.
2.5 Ultrafast band shift of Car in response to the BChl excitation
First,we consider the ultrafast Car band shift for the DDM preparation(Fig.7(a,b)):(i)B850 excitations(Fig.7(a)).Car band shift is seen as weak derivative type signal residing on the broad background of excited state absorption of BChl.A spectral bump(marked with asterisk)appears in the 0.5 ps transient to the red side of the 0-0 vibronic band of the ground state absorption.In the 10.0 ps transient this bump decays out and another weak one remains at the shorter wavelength side.(ii)B800 excitation(Fig.7(b)).The transient at 0.5 ps is characterized by the asterisked bump accompanied by another one to its shorter wavelength side.The transient at 10.0 ps closely resembles the transient of B850 excitation at the same delay time.This is understandable since at 10.0 ps the B800 excitation is gone and only the B850 excitation remains.Here,Car can be regarded as a spectator of the ultrafast BChl dynamics,e.g.,B800(Qy)-to-B850 (Qy)EET and relaxation among the manifold of B850 excitonic states.
The dynamics of Car band shift observed for the LDAO preparation(Fig.7(c,d))is similar to those for the DDM preparation(Fig.7(a,b))except that the asterisked bump observed for the DDM preparation is missing.This difference originates most likely from the subtle structural alteration in the LH2 assembly as induced by different detergents.Specifically,with reference to the 0-0 vibronic absorption band at 530 nm,the~10 nm red shift of the asterisked bump observed for the DDM preparation may arise from the Car composition 3 with NC=C=13,which preferentially senses the B850 excitation because of somehow favorable Car-BChl orientation.This speculation draws support from the difference between DDM and LDAO preparations in their B850 excitonic coupling strengths(Section 2.1).Here,it is worthy of noting that,besides other possible physiological roles,the photoprotection function for longer chain Cars,such as anhydrorhodovibrin and spirilloxanthin,is considered to be more important than light harvesting[42].Therefore,it is also physiologically reasonable to assume that spirilloxanthin is closer to BChls compared to other Car compositions.
3 Conclusions
We have examined,by the use of femtosecond time-resolved absorption spectroscopy,the detailed excitation dynamics of the isolated LH2 complex of Tch.tepidum prepared with the surfactants DDM or LDAO having different ionic properties.The spectral dynamics and the population kinetics of these LH2 preparations reveal efficient S2-state mediated Car-to-Car and Car-to-BChl singlet EET in a time scale of~100 fs,as well as the Qy-state mediated B800-to-B850 singlet EET for the DDM preparation in a time constant of~1.2 ps.These ultrafast EET processes suggest that the Cars with NC=C=11 and 12 coexist in a LH2 complex,and that the B800-B850 mutual orientation in the LH2 of Tch.tepidum differs considerably from those in the LH2s from the extensively investigated mesophilic species,e.g., Rps.acidophila and Rs.molischianum,etc.Anhydrorhodovibrin (NC=C=12)as a minor Car composition is found to act as an efficient trap of excitation energy,i.e.,itsexcitation,not transferring to BChl,relaxes rapidly to the intermediate statesthat either form triplet excitation or further relax to the ground state.The excitation-trap mechanism of anhydrorhodovibrin is considered to be important for the photoprotection function of Tch.tepidum.Furthermore,based on the ultrafast Car band shift in response to selective BChl excitation, we suggest that(OH-)spirilloxanthin locates in closer proximity to BChl compared to other Car compositions.Our results may facilitate to understand the light-harvesting and photoprotection mechanisms of Tch.tepidum as a thermophilic purple bacterium living under harsh natural conditions.
1 McDermott,G.;Prince,S.M.;Freer,A.A.;Hawthornthwaite-Lawless,A.M.;Papiz,M.Z.;Cogdell,R.J.;Isaacs,N.W.Nature, 1995,374:517
2 Papiz,M.Z.;Prince,S.M.;Howard,T.;Cogdell,R.J.;Isaacs,N. W.J.Mol.Biol.,2003,326:1523
3 Koepke,J.;Hu,X.;Muenke,C.;Schulten,K.;Michel,H.Structure, 1996,4:581
4 Sturgis,J.N.;Tucker,J.D.;Olsen,J.D.;Hunter,C.N.;Niederman, R.A.Biochemistry,2009,48:3679
5 Hu,X.;Damjanovic,A.;Ritz,T.;Schulten,K.Proc.Natl.Acad. Sci.U.S.A.,1998,95:5935
6 Sundström,V.;Pullerits,T.;van Grondelle,R.J.Phys.Chem.B, 1999,103:2327
7 Cogdell,R.J.;Gardiner,A.T.;Roszak,A.W.;Law,C.J.;Southall, J.;Isaacs,N.W.Photosynth.Res.,2004,81:207
8 Polívka,T.;Sundström,V.Chem.Rev.,2004,104:2021
9 Ritz,T.;Damjanovic,A.;Schulten,K.;Zhang,J.P.;Koyama,Y. Photosynth.Res.,2000,66:125
10 Hartigan,N.;Tharia,H.A.;Sweeney,F.;Lawless,A.M.;Papiz,M. Z.Biophys.J.,2002,82:963
11 Bahatyrova,S.;Frese,R.N.;Siebert,C.A.;Olsen,J.D.;van der Werf,K.O.;van Grondelle,R.;Niederman,R.A.;Bullough,P.A.; Otto,C.;Hunter,C.N.Nature,2004,430:1058
12 Scheuring,S.;Lévy,D.;Rigaud,J.L.Biochim.Biophys.Acta, 2005,1712:109
13 Madigan,M.T.Science,1984,225:313
14 Kimura,Y.;Hirano,Y.;Yu,L.J.;Suzuki,H.;Kobayashi,M.; Wang,Z.Y.J.Biol.Chem.,2008,283:13867
15 Ma,F.;Kimura,Y.,Zhao,X.H.;Wu,Y.S.;Wang,P.;Fu,L.M.; Wang,Z.Y.;Zhang,J.P.Biophys.J.,2008,95:3349
16 Ma,F.;Kimura,Y.;Yu,L.J.;Wang,P.;Ai,X.C.;Wang,Z.Y.; Zhang,J.P.FEBS J.,2009,276:1739
17 Sekine,F.;Horiguchi,K.;Kashino,Y.;Yu,L.J.;Wang,Z.Y.Gene sequences and characterization of light-harvesting complex 2 from Thermochromatium tepidum.(In preparation.For more information,please contact Prof.Wang,Z.Y.via wang@mx. ibaraki.ac.jp)
18 Suzuki,H.;Hirano,Y.;Kimura,Y.;Takaichi,S.;Kobayashi,M.; Miki,K.;Wang,Z.Y.Biochim.Biophys.Acta,2007,1767:1057
19 Han,R.M.;Wu,Y.S.;Feng,J.;Ai,X.C.;Zhang,J.P.;Skibsted, L.H.Photochem.Photobiol.,2004,80:326
20 Wang,Y.L.;Hu,X.C.J.Am.Chem.Soc.,2002,124:8445
21 Furuichi,K.;Sashima,T.;Koyama,Y.Chem.Phys.Lett.,2002, 356:547
22 Tavan,P.;Schulten,K.Phys.Rev.B,1987,36:4337
23 Rondonuwu,F.S.;Yokoyama,K.;Fujii,R.;Koyama,Y.;Cogdell, R.J.;Watanabe,Y.Chem.Phys.Lett.,2004,390:314
24 Frank,H.A.;Cogdell,R.J.The photochemistry and function of carotenoids in photosynthesis//Young,A.,Britton,G.Carotenoids in photosynthesis.London:Chapman&Hall,1993:252-326
25 He,Z.;Sundström,V.;Pullerits,T.J.Phys.Chem.B,2002,106: 11606
26 Linnanto,J.;Korppi-Tommola,J.E.I.;Helenius,V.M.J.Phys. Chem.B,1999,103:8739
27 Zerlauskiene,O.;Trinkunas,G.;Gall,A.;Robert,B.;Urboniene, V.;Valkunas,L.J.Phys.Chem.B,2008,112:15883
28 Akahane,J.;Rondonuwu,F.S.;Fiedor,L.;Watanabe Y.;Koyama Y.Chem.Phys.Lett.,2004,393:184
29 Zhang,J.P.;Inaba,T.;Watanabe,Y.;Koyama,Y.Chem.Phys. Lett.,2001,340:484
30 Dexter,D.L.J.Chem.Phys.,1953,21:836
31 Gradinaru,C.C.;Kennis,J.T.M.;Papagiannakis,E.;van Stokkum,I.H.M.;Cogdell,R.J.;Fleming,G.R.;Niederman,R. A.;van Grondelle,R.Proc.Natl.Acad.Sci.U.S.A.,2001,98: 2364
32 Zhao,X.H.;Liang,J.;Ma,F.;Su,W.J.;Wang,P.;Fu,L.M.;Ai, X.C.;Zhang,J.P.Chem.J.Chin.Univ.,2008,29:149 [赵晓辉,梁 俊,马 菲,苏文杰,王 鹏,付立民,艾希成,张建平.高等学校化学学报,2008,29:149]
33 Zhang,J.P.;Fujii,R.;Qian,P.;Inaba,T.;Mizoguchi,T.;Koyama, Y.;Onaka,K.;Watanabe Y.J.Phys.Chem.B,2000,104:3683
34 Fujii,R.;Inaba,T.;Watanabe,Y.;Koyama,Y.;Zhang,J.P.Chem. Phys.Lett.,2003,369:165
35 Feng,J.;Wang,Q.;Wu,Y.S.;Ai,X.C.;Zhang,X.J.;Huang,Y. G.;Zhang,X.K.;Zhang J.P.Photosynth.Res.,2004,82:83
36 Pullerits,T.;Chachisvilis,M.;Sundström,V.J.Phys.Chem., 1996,100:10787
37 Kennis,J.T.M.;Streltsov,A.M.;Vulto,S.I.E.;Aartsma,T.J.; Nozawa,T.;Amesz,J.J.Phys.Chem.B,1997,101:7827
38 Chen,X.H.;Zhang,L.;Weng,Y.X.;Du,L.C.;Ye,M.P.;Yang., G.Z.;Fujii,R.;Rondonuwu,F.S.;Koyama,Y.;Wu,Y.S.;Zhang, J.P.Biophys.J.,2005,88:4262
39 Herek,J.L.;Polívka,T.;Pullerits,T.;Fowler,G.J.S.;Hunter,C. N.;Sundström,V.Biochemistry,1998,37:7057
40 Herek,J.L.;Wendling,M.;He,Z.;Polívka,T.;Garcia-Asua,G.; Cogdell,R.J.;Hunter,C.N.;van Grondelle,R.;Sundström,V.; Pullerits,T.J.Phys.Chem.B,2004,108:10398
41 Zhang,J.P.;Nagae,H.;Qian,P.;Limantara,L.;Fujii,R.; Watanabe,Y.;Koyama,Y.J.Phys.Chem.B,2001,105:7312
42 Qian,P.;Saiki,K.;Mizoguchi,T.;Hara,K.;Sashima,T.;Fujii,R.; Koyama,Y.Photochem.Photobiol.,2001,74:444
