条斑星鲽幼鱼变态期间核酸及蛋白的变化
2010-10-23佟雪红徐世宏刘清华肖志忠马道远
佟雪红, 徐世宏 刘清华 李 军 肖志忠 马道远
(1. 中国科学院 海洋研究所, 山东 青岛 266071; 2. 中国科学院 研究生院, 北京 100039)
条斑星鲽幼鱼变态期间核酸及蛋白的变化
佟雪红1,2, 徐世宏1, 刘清华1, 李 军1, 肖志忠1, 马道远1
(1. 中国科学院 海洋研究所, 山东 青岛 266071; 2. 中国科学院 研究生院, 北京 100039)
本试验测定了条斑星鲽(Verasper moseri)幼鱼变态过程中DNA、RNA、蛋白及其比值的变化, 作为评价幼鱼变态阶段生长潜能的指标。结果表明, 试验期间培育水温为16.5~18.0°C, DNA、RNA及蛋白的变化都带有发育阶段特异性。DNA含量在34~42日龄增加缓慢, 然后快速增加直至 44日龄, 在45日龄和46日龄分别保持下降和上升趋势。蛋白含量和RNA含量的变化趋势相似, 在试验期间保持快速增长的趋势。RNA/DNA比值从试验开始至35日龄呈上升状态, 36日龄时下降, 38日龄时达到最高值5.01, 然后显著下降(P<0.05), 在试验结束时达到最低值1.76。蛋白/DNA在40日龄时达到最高值58.64, 在 46日龄时达到最低值 21.28。RNA、DNA及蛋白含量跟全长和体质量有明显的线性关系(P<0.05)。RNA/DNA与全长和体质量的关系比蛋白/DNA与全长和体质量的关系密切。蛋白/DNA比值的变化趋势跟 RNA/DNA比值类似, 但前者的变化滞后于后者, 表明 RNA/DNA比值是评价条斑星鲽生理状况的更有效的生理指标。
条斑星鲽(Verasper moseri); 幼鱼; DNA; RNA; 蛋白
鱼类在仔稚鱼阶段对外界环境的变化很敏感,此阶段以快速的生长率和剧烈的形态改变为标志[1~3]。鱼类的生长可以通过体长或者体质量来度量,而后者的变化可以体现在仔稚鱼核酸和蛋白组成的变化方面[3]。由于仔鱼的形体较小, 精确测定仔稚鱼形态指标比较困难, 因此传统的通过体长或者体质量衡量成鱼生长的方法并不适用于仔稚鱼[4,5], 加之有些鱼类在仔鱼阶段生长缓慢, 降低了应用体长或体质量来评价生长的可行性[6]。所以, 借助于生化指标来度量鱼类的生长是非常必要的。
近年来, 利用形态学、组织学及生化分析等方法评价养殖和野生鱼类仔稚鱼阶段生理状况的研究逐渐开展[7~9]。在通过传统方法不能度量出鱼类生长的变化时, 生化指标能够准确地检测到鱼类生长的细微变化和食物分布的波动[10]。最近, 仔稚鱼的生长潜能多采用以核酸为基础的指标(例如: RNA/DNA)进行度量, RNA/DNA比值反映了鱼类蛋白的合成能力[11~16]。采用以核酸为基础的指标来评价生长的基础在于动物细胞中DNA的量是恒定的, RNA的量跟蛋白合成的速度紧密相连[2]。由于仔鱼的生长依赖于蛋白的合成, 而且以核酸为基础的参数对仔稚鱼的摄食变化反映灵敏, 因此以核酸为基础的指标是评价仔稚鱼生长的有效参数[17~19]。国内, 赵玲等[20]对鲢(Hypophthalmichehys molitrix)、鳙(Aristichthys nobilis)、团头鲂(Megalobrama amblycephala)的研究,罗莉等[21]对草鱼(Ctenopharyngodon idellus)的研究以及梁萌青等[22]鲀对红鳍东方 (Takifugu rubripes)的研究均表明, RNA/DNA 比值是一个非常灵敏地反映鱼类生长的指标。另外, RNA/DNA 比值还可以推断天然水域中鱼类幼体的营养状况以及是否处于饥饿状态[23]。RNA/DNA 比值还被用来作为评价鱼用配合饲料优劣的指标和衡量水环境的指标, 能在一定条件下预测污染与否以及水环境是否有利于鱼类的生长[24,25]。因此, RNA/DNA比值在水产动物的生产和研究中是一个非常重要的指标。
条斑星鲽(Verasper moseri)俗称“花豹子”、“花边爪”, 属鲽形目(Pleuronectiformes)、鲽科(Pleuronectidae)、星鲽属(Verasper), 主要分布于日本茨城县以北到鄂霍茨克海以南海域, 中国黄渤海海域亦有少量分布, 为冷温性大型底栖鱼类。变态期往往伴随着营养危机和高的死亡率, 是鲆鲽鱼类早期发育阶段的关键期, 并决定着年产量和经济收益[26~28]。因此, 评价此阶段仔稚鱼的生理状况和生长潜能是非常必要的。本试验的目的在于探索条斑星鲽变态阶段核酸和总蛋白的变化规律, 明确生化指标跟生长的相关关系, 从而建立生化指标评价仔稚鱼生理状况的检测技术。
1 材料和方法
1.1 仔稚鱼的培育
实验所用条斑星鲽鱼苗取自山东烟台东方海洋科技股份有限公司。样品为孵化后 34~46日龄的鱼苗。仔稚鱼在16 m3的水泥池中流水培育, 培育时的水质条件是: 水温为 16.5~18.0 °C, 溶氧为 7.0~8.5 mg/L, 盐度为30~32, pH为7.7~8.0。试验期间投喂人工配合饵料。
1.2 样品采集和保存
试验自鱼苗孵化后34 d开始至46 d结束。2 d取样1次, 8: 00定点取样45尾, 其中15尾先用丁香酚麻醉, 再用冰冷的去离子水冲洗并快速冻于液氮中保存, 用于测定 RNA、DNA及蛋白。其余的 30尾用于测定全长、体长及体质量。
1.3 核酸及蛋白的测定
参照Buckley[29]和Kuropat等[30]的方法, 采用整体匀浆法提取条斑星鲽仔稚鱼的核酸和蛋白。用GeneQuantpro紫外分光光度计测定各自的 A260值,并计算RNA 和DNA 的含量。依据Bradford[31]的方法测定蛋白。
1.4 数据分析
试验所得数据表示为平均值±标准差。采用SPSS 11.0 for Windows统计软件中 One-way ANOVA分别分析核酸及蛋白在不同生长时期的差异显著性, 当P<0.05时认为差异显著。
2 结果
2.1 条斑星鲽的生长变化
条斑星鲽变态期间体长和体质量的变化见图1。鱼苗34日龄时开始变态, 44日龄时变态结束。变态开始时体质量和体长分别是23.5mg、1.1cm。体质量的增长程度高于体长的增长程度。变态高峰期时, 体长和体质量增长迅速; 变态结束时, 体长和体质量的增长速度变慢。
2.2 核酸及蛋白的变化
本论文采用两种单位(质量比和含量)分别来阐述 DNA、RNA及蛋白的变化规律。质量比表示为µg/mg 湿质量, 指的是每毫克体质量下DNA、RNA及蛋白的量; 含量表示为µg/尾, 指的是每尾鱼所具有的DNA、RNA及蛋白的量。
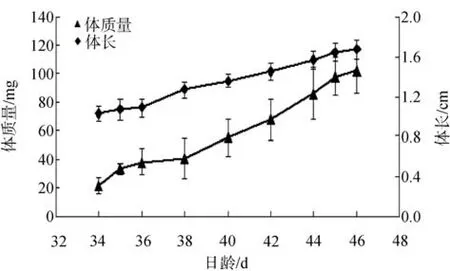
图1 条斑星鲽仔稚鱼体质量和体长的变化Fig. 1 Changes in body weight and body length of barfin flounder larvae and juveniles

图2 条斑星鲽仔稚鱼DNA, RNA和蛋白质量比的变化平均值后字母不同者表示差异显著, 图3、图4同Fig. 2 Changes in DNA, RNA and protein concentrations of barfin flounder larvae and juvenilesMeans followed by different letters are significantly different
DNA质量比在32至42日龄期间保持恒定, 在44日龄时显著上升(P<0.05), 然后呈下降趋势, 在46日龄时数值升至最高(图2)。RNA质量比从实验开始至变态高峰期增长快速, 之后逐渐下降。蛋白质量比在40日龄之前保持波动状态, 然后维持在较高的稳定水平。
DNA、RNA及蛋白含量在试验期间均保持快速增长的趋势, 在 40~44日龄期间增加显著(P<0.05),但在45日龄时缓慢下降(图3)。

图3 条斑星鲽仔稚鱼DNA, RNA和蛋白含量的变化Fig. 3 Changes in DNA, RNA and protein contents of barfin flounder larvae and juvenilesMeans followed by different letters are significantly different
2.3 RNA/DNA和蛋白/DNA的变化
RNA/DNA比值在试验期间保持波动趋势(图4),试验开始至35日龄该比值呈上升状态, 36日龄时下降, 在 38日龄时达到最高值 5.01, 然后显著下降(P<0.05), 在试验结束时达到最低值1.76。蛋白/DNA变化趋势跟RNA/DNA变化趋势类似, 在40日龄时达到最高值58.64, 在试验结束时达到最低值21.28。

图4 条斑星鲽仔稚鱼蛋白/DNA和RNA/DNA的变化Fig. 4 Changes in protein/DNA and RNA/DNA ratios of barfin flounder larvae and juvenilesMeans followed by different letters are significantly different
2.4 DNA、RNA、蛋白及其比值与全长和体质量的关系
本实验中, 以核酸为基础的指标与全长和体质量的关系见图5和表1。DNA、RNA及蛋白含量跟全长有明显的线性关系,R2分别是 0.772、0.921、0.906 (P<0.05)。DNA、RNA及蛋白含量跟体质量也有明显的线性关系,R2分别是 0.851、0.987、0.985(P<0.05)。RNA/DNA与全长和体质量的线性关系R2分别是0.465、0.531; 蛋白/DNA与全长和体质量的线性关系R2分别是0.337、0.377; RNA/DNA与全长和体质量的关系比蛋白/DNA与全长和体质量的关系密切。蛋白增长率与全长和体质量无明显的线性关系,R2分别为0.06、0.025。
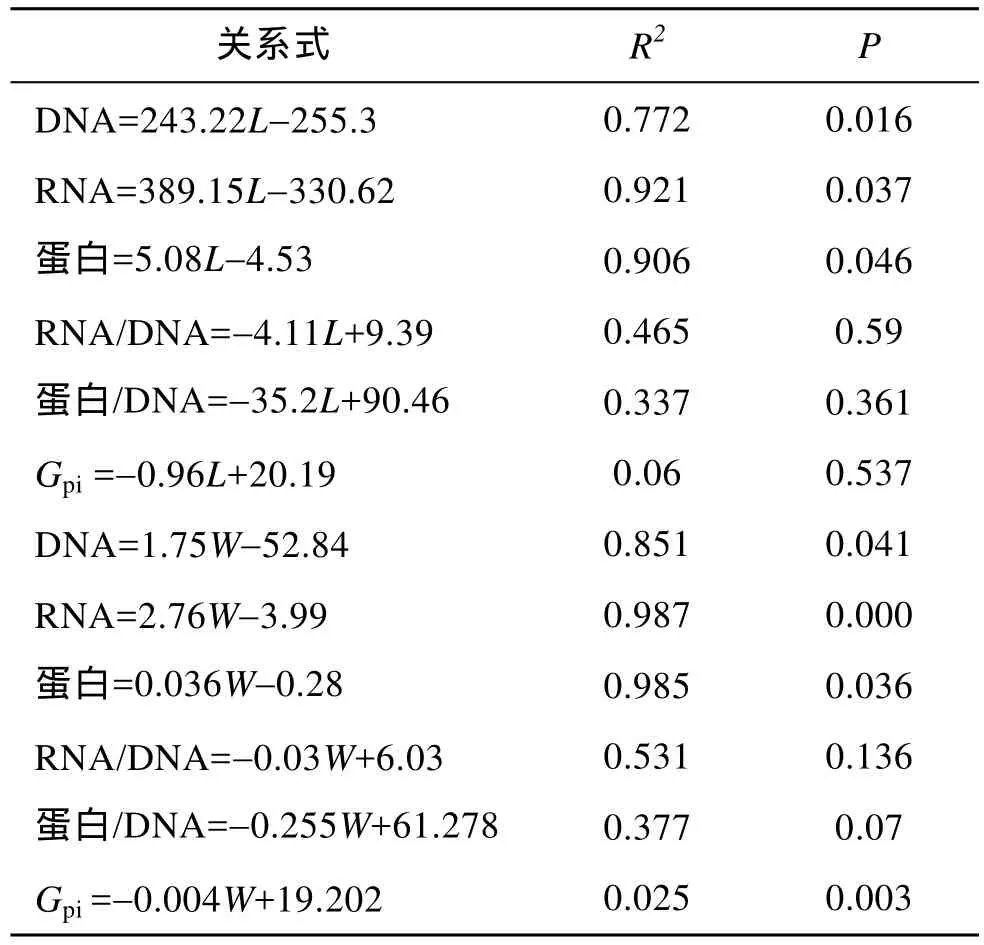
表1 蛋白含量、以核酸为基础的指标及蛋白增长率与条斑星鲽仔稚鱼全长和体质量的回归关系Tab. 1 Regression equations of protein, the nucleic acids-based indices or Gpi with total length and body weight of barfin flounder larvae and juveniles
3 讨论
3.1 条斑星鲽变态期间核酸及蛋白的变化
核酸在鱼类的生长和发育过程中起着重要作用。DNA的量反映了鱼体的细胞数量, 而RNA的量则跟细胞中蛋白的合成直接相关[2]。本试验中, RNA质量比在 38日龄之前呈上升趋势, 然后下降, 而DNA质量比在此期间几乎没有变化, 表明从变态开始至变态高峰期, 条斑星鲽的生长以细胞增大为主;鱼类变态是一个耗能的过程, 所以, 变态后期 RNA量会降低[27]。RNA和蛋白含量在条斑星鲽变态期间呈快速增长的趋势, 这可能与鲆鲽类变态期间内脏器官的发育和完善密切相关[2,32]。本实验中蛋白含量的变化趋势跟RNA含量的变化趋势类似, 说明RNA含量变化反映了鱼体蛋白合成的多少。
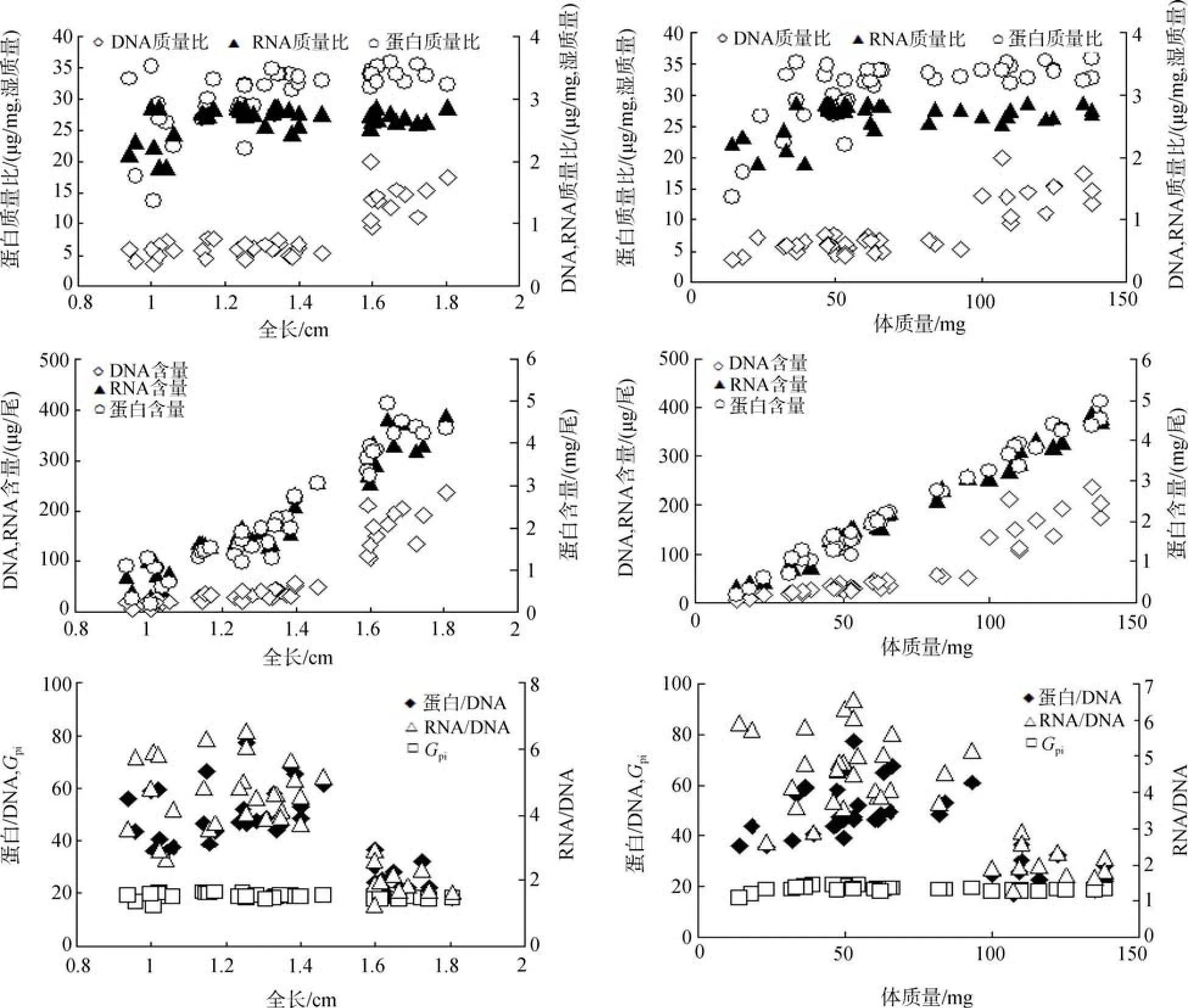
图5 条斑星鲽仔稚鱼全长和体质量与蛋白、RNA、DNA、RNA/DNA、Gpi及蛋白/DNA的关系Fig. 5 Relationships of protein, RNA, DNA, RNA/DNA, Gpi or protein/DNA with total length and body weight of barfin flounder larvae and juveniles
DNA含量是生物体内细胞数目的指示因子[32]。42日龄时 DNA含量的急剧增长可能预示了此时条斑星鲽鱼苗比较脆弱[33], 对外界反映很敏感, 该时期条斑星鲽正经历由浮游状态到底栖状态的改变。本实验中以核酸为基础的指标从细胞水平上反映了条斑星鲽的生长特性: 条斑星鲽变态期间的生长以细胞增大为主, 而在变态后的生长以细胞增殖为主。与在牙鲆中得出的试验结果类似[1], 本试验中 DNA和RNA含量的变化跟发育阶段和仔稚鱼大小密切相关。另外, 鲆鲽类变态期间, 内在器官经历分化和成熟的过程[34], 因此, 本试验中RNA和DNA含量的变化跟此发育阶段器官的分化增殖也有一定联系。
3.2 条斑星鲽变态期间 RNA/DNA比值的变化
RNA/DNA是生物体中细胞代谢强度的指示指标, 因此常被用来评价鱼类的生理状况[35]。本实验中RNA/DNA呈波动性变化, 主要是由于 RNA含量波动和DNA含量相对稳定的缘故。该比值在36日龄时急剧下降, 原因在于此时 RNA量的下降和 DNA量的增长, 由此进一步说明此时细胞增殖的速度要高于蛋白合成的速度[1]。在38日龄时, RNA/DNA比值上升至最高峰, 表明此时蛋白合成的速度快于细胞增殖的速度, 合成的蛋白可能跟变态高峰时器官的分化增殖有关。RNA/DNA比值在变态后显著下降,表明细胞中蛋白合成的降低和细胞数目的增多, 这可能跟此时期条斑星鲽稚鱼生长方式由浮游状态转变为底栖生活所伴随的能量消耗有关。本研究中,RNA/DNA比值随鱼体增大呈下降趋势, Kimura等[35]在评价日本沙丁鱼初次摄食时的营养状况时, 以及Clemmesen[36]在分析饵料分布对青鱼 RNA/DNA 比值的影响时, 均得出了类似结果。
对牙鲆进行的相关研究显示, RNA/DNA有一个关键的临界值为2.49[1,37], RNA/DNA低于此临界值的牙鲆仔稚鱼处于“亚健康”状态或者饥饿状态。本研究中, 条斑星鲽从38日龄开始RNA/DNA比值急剧下降, 而且在44日龄后该比值均维持在2左右,这可以从条斑星鲽生活习性的改变上得到解释, 条斑星鲽变态后将由浮游生活转为伏底生活, 栖息环境的转变可能会影响条斑星鲽的摄食并进一步导致生理状况和营养水平的急剧变化, 从而使得 RNA/DNA下降至临界值周围。因此, 刚伏底的一段时间往往是条斑星鲽的死亡高发期。
3.3 条斑星鲽变态期间蛋白/DNA比值的变化
蛋白/DNA是生物体中细胞大小或者细胞质量的指标, 因为蛋白占据了细胞的绝大部分质量[32]。38日龄时蛋白/DNA比值明显下降, 主要是由于变态高峰期时的巨大能量消耗和器官分化所需要的细胞快速增殖所致。而在40日龄该比值达到最高峰, 原因在于生物体快速合成了大量蛋白以应对伏底期的营养危机。条斑星鲽转入伏底后, 生物体将大量耗能,表现为蛋白/DNA比值的急剧下降。Fukuda等[32]发现, 施氏鲽(Pleuronectes schrenki)在变态后蛋白/DNA比值也呈下降趋势。蛋白/DNA跟RNA/DNA的变化趋势相似, 但在时间上相对滞后, 说明RNA/DNA是评价条斑星鲽生理状况和生长的更有效的指标。
以上结果表明, 条斑星鲽变态期间的生长以细胞增大为主, 而在变态后的生长以细胞增殖为主,以核酸为基础的指标可以从细胞水平上评价条斑星鲽仔稚鱼的生长。
[1] Gwak W S, Tanaka M. Changes in RNA, DNA and protein contents of laboratory-reared Japanese flounder Paralichthys olivaceus during metamorphosis and settlement [J]. Fisheries Sci, 2002, 68: 27-33.
[2] Tanaka Y, Gwak W S, Tanaka M, et al. Ontogenetic changes in RNA, DNA and protein contents of laboratory-reared Pacific bluefin tuna Thunnus orientalis[J].Fisheries Sci, 2007, 73: 378-384.
[3] Park S U, Lim H K, Han H S. Changes in RNA ⁄DNA ratio and growth of slime flounder, Microstomus achne,larvae until metamorphosis[J]. J Appl Ichthyol, 2008, 24:50-54.
[4] Fraser A J, Sargent J R. Lipid class and fatty acid composition as indicators of the nutritional condition of larval Atlantic herring [J]. Am Fish Soc Symp, 1987, 2:129-143.
[5] Westerman M E, Holt G J. RNA: DNA ratio during the critical period and early larval growth of the red drum Sciaenops ocellatus [J]. Mar Biol, 1994, 121: 1-9.
[6] Mercaldo-Allen R, Kuropat C, Caldarone E M. A model to estimate growth in young-of-the-year tautog,Tautoga onitis, based on RNA: DNA ratio and seawater temperature[J]. J Exp Mar Biol Ecol, 2006, 329:187-195.
[7] Buckley L J. Relationships between RNA-DNA ratios,prey density, and growth rate in Atlantic cod (Gardus morhua) larvae[J]. J Fish Res Bd Can, 1979,36: 1 497-1 502.
[8] Yin M C, Blaxter J H S. Morphological changes during growth and starvation of larval cod Gadus morhua and flounder Platichthys flesus[J]. J Exp Mar Biol Ecol,1986, 104: 215-228.
[9] Clemmesen C M. An RNA and DNA fluorescent technique to evaluate the nutritional condition of individual marine fish larvae[J]. Meeresforsch, 1988, 32:134-143.
[10] Haines T A. An evaluation of RNA-DNA ratio as a measure of long-term growth in fish populations[J]. J Fish Res Bd Can, 1973, 30: 195-199.
[11] Mathers E M, Houlihan D F, Cunningham M J. Nucleic acid concentrations and enzyme activities as correlates of growth rate of the saithe Pollachius virens:growth-rate estimates of open-sea fish[J]. Mar Biol, 1992,112: 363-369.
[12] Buckley L J, Calderone E, Ong T L. RNA-DNA ratio and other nucleic acid-base indicators for growth and condition of marine fishes[J]. Hydrobiology, 1999, 401:265-277.
[13] Fukuda M, Sako H, Shigeta T, et al. Relationship between growth and biochemical indices in laboratory-reared juvenile Japanese flounder (Paralichthys olivaceus) and its application to wild fish[J]. Mar Biol,2001, 138: 47-55.
[14] Peck M A, Buckley L J, Caldarone E M, et al. Effects of food consumption and temperature on growth rate and biochemical-based indicators of growth in early juvenile Atlantic cod Gadus morhua and haddock Melanogrammus aeglefinus [J]. Mar Ecol Prog Ser, 2003,251: 233-243.
[15] Yamashita Y, Tominaga O, Takami H, et al. Comparison of growth, feeding and cortisol level in Platichthys bicolaratus juveniles between estuarine and nearshore grounds[J]. J Fish Biol, 2003, 63: 617-630.
[16] Vinagre B C, Fonseca V, Maia A, et al. Habitat specific growth rates and condition indices for the sympatric soles Solea solea (Linnaeus, 1758) and Solea senegalensis Kaup 1858, in the Tagus estuary, Portugal,based on otolith daily increments and RNA-DNA ratio[J]. J Appl Ichthyol, 2008, 24: 163-169.
[17] Varnavskiy V S, Varnavskaya N V, Kalinin S V, et al.RNA: DNA index as an indicator of growth rate of coho salmon (Oncorhynchus kisutch) during early marine life[J]. J Ichthyol, 1992, 32: 10-19.
[18] Bisbal G A, Bengtson D A. Description of the starving condition in summer flounder, Paralichthys dentatus,early life history stages[J]. Fish Bull, 1995, 93:217-230.
[19] Gwak W S, Tanaka Y, Tominaga O, et al. Field evaluation by RNA: DNA ratios on post-release nutritional status of released and wild juvenile Japanese flounder Paralichthys olivaceus juveniles[J]. J Exp Mar Biol Ecol, 2003a, 293: 107-124.
[20] 赵玲, 周洪琪, 陆海祺, 等. 对草鱼、鲢、鳙、团头鲂鱼种阶段的 RNA/DNA的初步研究[J]. 上海水产大学学报, 1995, 4(3): 199-203.
[21] 罗莉, 文华, 王琳, 等. 牛磺酸对草鱼生长、品质、消化酶和代谢酶活性的影响[J]. 动物营养学报, 2006,18 (3): 166-171.
[22] 梁萌青, 王成刚, 陈超, 等. 几种添加剂对红鳍东方鲀的促生长效果与 RNA/DNA 关系[J]. 海洋水产研究, 2001, 22 (2): 38- 41.
[23] Islam M S, Tanaka M. Nutritional condition, starvation status and growth of early juvenile Japanese sea bass(Lateolabrax japonicus) related to prey distribution and feeding in the nursery ground[J]. J Exp Mar Biol Ecol,2005, 323: 172-183.
[24] 赵振山, 赵可椒, 张益明, 等. 用 RNA/DNA 比值评定鲤鱼的生长及其配合饲料的营养价值[J]. 水产学报, 1994, 18 (4): 257-264.
[25] 甘居利, 金有坤. 用鱼肌核酸指示水体污染的初步研究[J]. 农业环境保护, 1997, 16 (2): 68-73.
[26] Thorisson K. Is metamorphosis a critical interval in the early life of marine fishes [J]. Environ Biol Fish, 1994,40: 23-36.
[27] Tanaka M, Kawai S, Seikai T, et al. Development of the digestive organ system in Japanese flounder in relation to metamorphosis and settlement [J]. Mar Fresh Behav Physiol, 1996, 28: 19-31.
[28] Amara R, Galois R. Nutritional condition of metamorphosing sole: spatial and temporal analyses [J].J Fish Biol, 2004, 64: 72-88.
[29] Buckley L J, Bulow F J. Techniques for the estimation of RNA, DNA, and protein in fish[A]. Summerfelt R C,Hall G E. The Age and Growth of Fish[C]. Ames, IA:Iowa State University Press, 1987. 345-354.
[30] Kuropat C, Mercaldo-Allen R, Caldarone E, et al.Evaluation of RNA concentration as an indicator of growth in young-of-the-year winter flounder Pseudopleuronectes americanus and tautog Tautogaonitis[J].Mar Ecol Prog Ser, 2002, 230: 265-274.
[31] Bradford M M. A rapid sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein–dye binding[J]. Anal Biochem,1976, 72: 248-254.
[32] Fukuda M, Yano Y, Nakano H, et al. Protein and nucleic acid changes during early development stages of cresthead flounder[J]. Bull Jpn Soc Sci Fish, 1986, 52:951-955.
[33] Malzahn A M, Clemmesen C, Rosenthal H. Temperature effects on growth and nucleic acids in laboratory-reared larval coregonid fish[J]. Mar Ecol Prog Ser, 2003, 259: 285-293.
[34] Youson J H. First Metamorphosis[A]. Hoar W S, Randall D J. Fish Physiology[C]. New York: Academic Press, 1988. 135-196.
[35] Kimura R, Watanabe Y, Zenitani H. Nutritional condition of first-feeding larvae of Japanese sardine in the coastal and oceanic waters along the Kuroshio Current[J]. J Mar Sci, 2000, 57: 240-248.
[36] Clemmesen C. The effect of food availability, age or size on the RNA/DNA ratio of individually measured herring larvae: laboratory calibration[J]. Mar Biol,1994, 118: 377-382.
[37] Gwak W S, Tanaka M. Developmental change in RNA/DNA ratios of fed and starved laboratory-reared Japanese flounder larvae and juveniles, and its application to assessment of nutritional condition for wild fish[J]. J Fish Biol, 2001, 59: 902-915.
Changes in nucleic acids and protein indices during metamorphosis of barfin flounder Verasper moseri
TONG Xue-hong1,2, XU Shi-hong1, LIU Qing-hua1, LI Jun1, XIAO Zhi-zhong1,MA Dao-yuan1
(1. Institute of Oceanology, the Chinese Academy of Sciences, Qingdao 266071, China ; 2. Graduate University of Chinese Academy of Sciences, Beijing 100039, China)
Mar., 15, 2010
barfin flounder (Verasper moseri); juvenile ; DNA ; RNA ; protein
Changes in DNA, RNA, protein and their ratios of barfin flounder Verasper moseri during metamorphosis were determined, and were used to evaluate the growth potential of barfin flounder at this stage. Water temperature fluctuated between 16.5°C and 18.0°C. Patterns of DNA, RNA and protein parameters all showed developmental stages-specific traits. DNA content remained slight increase until 42 days after hatching (DAH) and showed abrupt rise towards 44 DAH followed by a drop and a rise around 45 and 46 DAH, respectively. The changes in protein content coincided well with those of RNA content; and they both kept obvious increases throughout the experimental period. RNA/DNA ratio increased up to 35 DAH, but decreased at 36 DAH, and reached the climax of 5.01 at 38 DAH, which was followed by a significant decline with the minimum value of 1.76 at the end of the experiment. Protein/DNA reached the maximum value of 58.64 at 40 DAH and the minimum value of 21.28 at 46 DAH. DNA, RNA and protein contents all showed significant correlations with total length and body weight of barfin flounder (P<0.05). RNA/DNA had stronger correlations with total length and body weight of barfin flounder than protein/DNA. Protein/DNA showed a similar pattern to RNA/DNA, but the changes in protein/DNA lagged behind those in RNA/DNA, suggesting that RNA/DNA ratio is a more sensitive index than protein/DNA ratio in evaluating physiological conditions of barfin flounder.
Q955
A
1000-3096(2010)05-0041-07
2010-03-15;
2010-03-23
现代农业产业技术体系建设专项资金资助项目(nycytx-50);国家863“十一五”计划资助项目(2006AA10A404)
佟雪红(1979-), 女, 山东潍坊人, 博士研究生, 研究方向:海洋生物学, 电话: 0532-82898716, E-mail: txh113005@163.com; 李军,通信作者, 研究员, E-mail: junli@ms.qdio.ac.cn
(本文编辑: 谭雪静)
