基于DNA染料EMA的PCR技术检测鉴别副溶血性弧菌死活细胞*
2010-01-13祝儒刚吕淑霞刘月萍张喆
祝儒刚,吕淑霞,刘月萍,张喆
1(沈阳农业大学食品科学院,辽宁沈阳,110866)2(沈阳农业大学生物科学技术学院,辽宁沈阳,110866)3(辽宁大学轻型产业学院,辽宁沈阳,110036)
副溶血弧菌(Vibrio parahaemolyticus,VP)是海洋和盐湖中分布的一种嗜盐性病原菌。分布在近海岸的海水、海底沉积物、鱼类、贝类中,适应于高浓度(6%)食盐的环境中生长。该菌是亚于霍乱弧菌的一种常见肠道致病菌,其污染会给海水养殖业带来严重损失,还会引起人类腹泻等肠道疾病及食物中毒,出现腹泻、腹部痉挛、恶心、呕吐和头痛等症状。自2001年以来,副溶血弧菌性食物中毒爆发已经成为中国最常见的食源性疾患[1-4]。
随着人们对食品安全性要求的不断提高,PCR技术以其特异性强、灵敏度高和快速准确等优点已经成为检测海产品中副溶血弧菌的最主要手段[5-6]。然而,此法的缺点是不能区分所检测到的副溶血弧菌是活细胞还是死细胞。近年来,人们尝试各种方法通过PCR技术仅仅检测样品中的活菌,例如mRNA的检测,以及利用能够与核酸共价结合的染料,这种染料能够选择性地渗透到死细胞内部与细胞内的DNA共价结合。在所有这些方法中,一种DNA插入型染料EMA(ethidium bromide monoazide),展现出了很大的应用潜力,能够克服所面临的困难。许多研究表明,EMA与PCR技术相结合,能够成功地选择性抑制样品中死细胞的DNA扩增,以便于更加精确地检测样品中活菌细胞的存在[7-10]。
EMA是一种荧光插入型的核酸结合染料,这种染料最显著的特点是当样品中死活细胞共同存在时,它能够选择性地与死细胞DNA共价结合。由于活菌细胞的细胞膜比较完整,EMA不能渗透到活菌细胞内部。然而,由于通常死细胞的细胞膜已经遭到破坏,EMA很容易渗透到细胞内部从而插入到细胞内DNA双螺旋上。在经过高强度的可见光曝光处理后,插入到DNA双螺旋上的EMA能够与DNA双螺旋发生共价交叉偶合作用,从而导致死细胞内部的DNA在接下来的PCR扩增过程中被抑制[11-12]。
本研究将EMA选择渗透性和PCR特异性和灵敏性相结合,从而建立一种快速、灵敏并能够有效检测和定量纯培养条件下副溶血弧菌死活细胞混合液中的活细胞的新方法。
1 材料与方法
1.1 菌种
实验用标准菌株,副溶血弧菌ATCC17802,本实验室保存。
1.2 仪器与设备
PCR仪(TC-512),紫外可见分光光度计(752N),卤钨灯(500W),BIO-RAD凝胶成像仪。
1.3 试剂及培养基
EMA(ethidium bromide monoazide,美国 Sigma公司),胰胨肉汤(蛋白胨2%,NaCl 4%,0.01%的结晶紫0.5%,pH值9.0)。
1.4 副溶血弧菌热致死时间确定
取37℃、150 r/min条件下过夜培养的副溶血弧菌菌悬液10 mL,10 000r/min离心10 min。菌体沉淀悬浮于10 mL 0.85%的生理盐水中,10 000 r/min离心10 min。菌体沉淀再悬浮在10 mL 0.85%的生理盐水中,再次 10 000 r/min离心 10 min。测定OD600nm并进行平板计数。
取0.5 mL上述菌悬液调整其浓度为2×108CFU/mL至1.5 mL离心管中,95℃水浴加热,每隔30 s取样涂平板,37℃过夜培养。
1.5 最佳曝光时间确定
用ddH2O配制0.1 mg/mL的EMA溶液,-20℃避光保存(EMA为致癌物质,注意操作安全)。分别取0.5 mL热致死副溶血弧菌菌悬液(2×108CFU/mL)于7个小离心管,加入EMA使每管EMA终浓度达到1.4 μg/mL。充分混匀后,将离心管黑暗中室温放置5 min。然后将离心管开盖放置冰上,距离灯管16 cm,分别曝光 0、1、3、5、10、15 和 20 min。同样方法取活菌菌悬液作为对照。
1.6 不抑制活细胞PCR扩增的最大EMA浓度确定
取EMA溶液(1 mg/mL)分别加入装有0.5 mL活细胞的副溶血弧菌菌悬液(2×108CFU/mL)的7支离心管中,使 EMA终浓度分别达到0 μg/mL、2 μg/mL、4 μg/mL、6 μg/mL、8 μg/mL、10 μg/mL 和 12 μg/mL。充分混匀后,将离心管黑暗中室温放置约5 min。然后将离心管开盖放置冰上,距离灯管16 cm,曝光20 min。
1.7 完全抑制死细胞PCR扩增的最小EMA浓度确定
将EMA(0.1 mg/mL)分别加入装有0.5 mL热致死副溶血弧菌菌悬液(2×108CFU/mL)的9支离心管中,使 EMA 终浓度分别达到 0、0.2、0.4、0.6、0.8、1.0、1.2、1.4 和 1.6 μg/mL。充分混匀后,将离心管黑暗中室温放置约5 min。然后将离心管开盖放置冰上,距离灯管16 cm,曝光20 min。
1.8 死活细胞混合菌悬液的EMA-PCR扩增
在完全一样的8只离心管中分别加入0.25 mL固定数量(5×107CFU)的热致死细胞菌悬液与0.25 mL 变化数量(5 ×107、5 ×106、5 ×105、2.5 ×105、5 ×104、2.5×104、5×103和 2.5 ×103CFU)的活细胞菌悬液(生理盐水漂洗),混合均匀。向菌悬液中加入0.1 mg/mL的EMA,使EMA终浓度达到1.4 μg/mL。将菌悬液在黑暗中室温放置5 min,然后将离心管开盖放置冰上,距离灯管16 cm曝光20 min。曝光后的混合菌悬液在10 000 r/min离心5 min,收集菌体。倒掉上清液后将菌体重悬在0.5 mL的生理盐水中,10 000 r/min再次离心5 min,收集菌体,倒掉上清液,再次重悬在0.5 mL生理盐水中,提取模板DNA。
1.9 DNA 模板制备[13-14]
用TZ裂解液(2.0%Triton X-100,2.5 mg/mL的叠氮化钠,0.1 mol/L Tris-HCl缓冲液,pH8.0)提取模板DNA。
0.5 mL的各种菌悬液与等体积的2×TZ裂解液混匀,沸水浴10 min后冷却到室温,8 000 r/min离心5 min去掉菌体碎片沉淀,取上清液直接用作PCR模板。
1.1 0 引物及PCR扩增
根据GenBank中已发表的副溶血弧菌tlh基因(基因序列号AY578148)序列,利用Primer 5.0软件设计一对PCR引物,上游引物(P1)为:5'-AAG CGG ATT ATG CAG AAG CA-3',下游引物(P2)为:5'-GCT ACT TTC TAG CAT TTT CTC TGC-3',扩增片段长度为449bp。优化PCR扩增条件为:25 μL的PCR反应体系中含 4 μL 模板,2.5 μL 的 10 × PCR Buffer,各10pmol的引物,2 μL 的 25 mmol/L MgCl2,2 μL 的2.5 mmol/L dNTP,0.2 μL 的 Taq酶,用灭菌水补足到25 μL体系。PCR反应条件为94℃ 5 min,29个循环,每个循环 94℃ 1 min、55℃ 1 min、72℃ 2 min,最后 72℃延伸 10 min[15-18]。
1.1 1 PCR产物检测及定量分析
取10 μL PCR产物在1.5%琼脂糖凝胶上进行电泳(100 V,1 h),凝胶成像系统观察结果并成像。利用NIH1.61图像分析软件定量分析混合液中各DNA条带的相对荧光强度,相对荧光强度取3次独立实验的平均值。
2 结果与分析
2.1 副溶血弧菌热致死时间
将原始菌悬液做梯度稀释,测定各梯度的OD600nm值,取其中一个梯度悬液涂平板,设3个重复,37℃过夜培养,计算每个梯度的菌浓度。得到活菌的菌浓度随OD600nm变化曲线,如图1所示。由图1可知,当 OD600nm为0.42时,菌浓度正好为2×108CFU/mL。
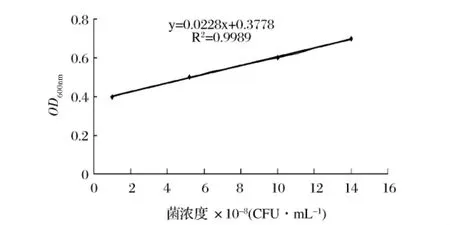
图1 菌浓度与OD600nm的标准曲线
95℃水浴处理副溶血弧菌菌悬液(OD600nm为0.42,2×108CFU/mL),每隔 30s取样涂平板,每个样品设3个重复,37℃过夜培养后观察计数(表2)。结果表明,随着热处理时间的增加,其平板菌落数逐渐减少,当热处理时间延长到9 min时,副溶血弧菌已失活,平板上没有菌落生长,平板计数为0。因此,为了确保所有副溶血弧菌完全被杀死,得到结果:将浓度为2×108CFU/mL的副溶血弧菌菌悬液在95℃水浴处理10 min,所有副溶血弧菌细胞都已经杀死,平板计数为0。

表1 不同加热时间处理的活细胞数
2.2 最佳曝光时间
热致死细胞和活细胞菌悬液(2×108CFU/mL)分别经EMA(终浓度1.4 μg/mL)处理后,曝光1~20 min,结果如图2和图3所示。
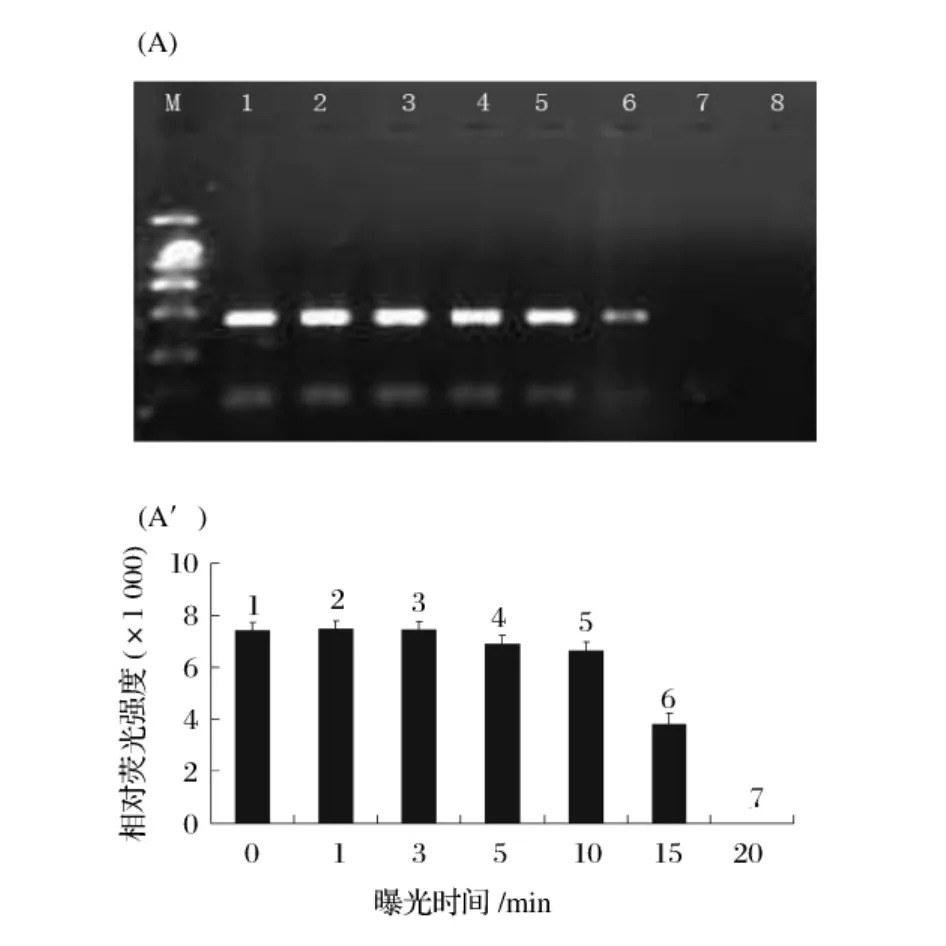
图2 EMA处理热致死细胞的最佳曝光时间优化(A):条带1~7分别为EMA浓度为1.4μg/mL时热致死细胞曝光 0、1、3、5、10、15 和 20 min 后的 PCR 凝胶电泳图。条带8为阴性对照(不加模板DNA)。M为DNA Marker。(A'):为(A)凝胶电泳图上相应DNA条带的相对荧光强度。

图3 EMA处理活细胞的最佳曝光时间优化(B):条带1~7分别为EMA浓度为1.4μg/mL时活细胞曝光0、1、3、5、10、15和 20 分钟后的 PCR 凝胶电泳图。条带8为阴性对照(不加模板DNA)。M为DNA Marker。(B'):为(B)凝胶电泳图上相应DNA条带的相对荧光强度。
随着曝光时间的延长,经EMA(终浓度1.4 μg/mL)处理后死细胞的PCR扩增条带逐渐变弱,当曝光时间延长到15 min,PCR扩增产物的荧光条带变得很淡,曝光时间延长到20 min时荧光条带完全消失(P<0.01)。而对照组活菌细胞则随着曝光时间的延长,荧光条带没有明显变化。因此,在整个试验中都以20 min为最佳EMA曝光时间。
2.3 不抑制活细胞PCR扩增的最大EMA浓度
EMA的不同添加量对活菌PCR扩增的抑制作用电泳图谱如图4。随着EMA浓度的逐渐增加,PCR扩增条带也逐渐变弱,表明EMA浓度足够大时对活菌的PCR扩增也有一定抑制作用。图谱分析可知,当EMA浓度小于等于2 μg/mL时,EMA对活细胞PCR扩增没有明显抑制作用(P>0.01)。然而,当EMA的添加量增加到4 μg/mL时,EMA对活细胞的PCR扩增有显著的抑制作用(P<0.01)。因此,在本实验中我们选择不抑制活细胞PCR扩增的最大EMA浓度为 2 μg/mL。
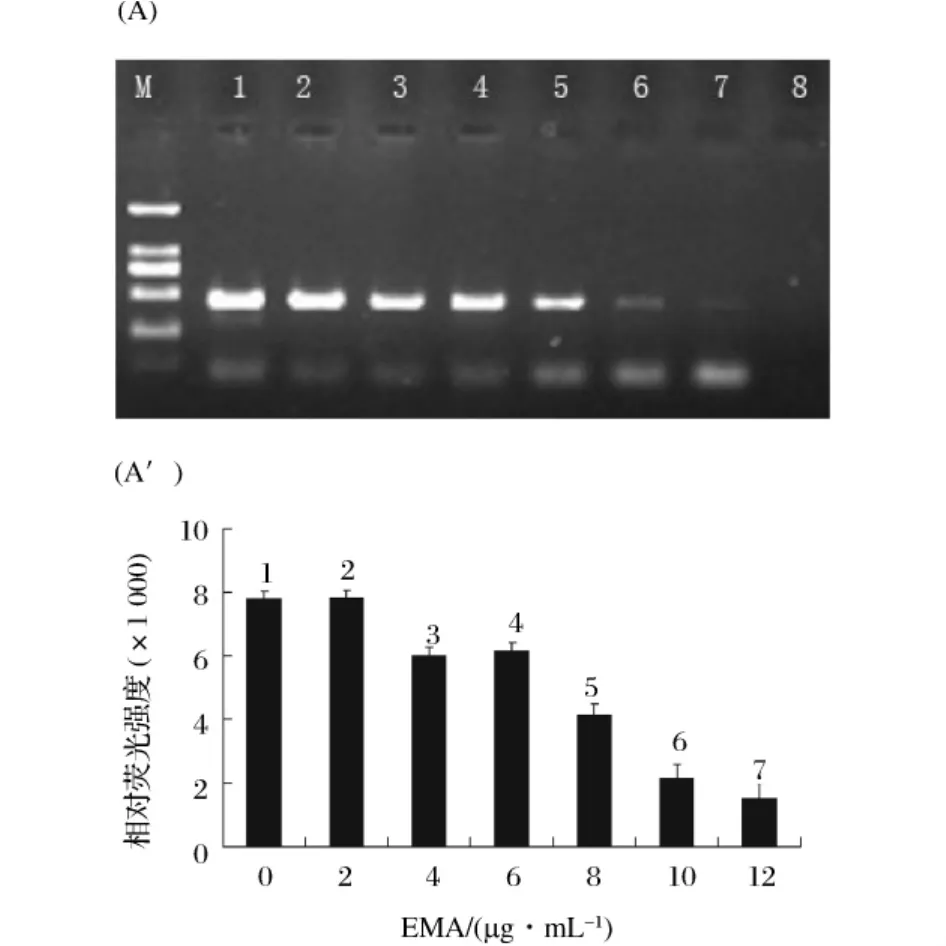
图4 不抑制活菌细胞DNA扩增的最大EMA浓度优化(A):不同EMA浓度下活菌细胞EMA-PCR产物的琼脂糖凝胶电泳图。M为DNA Marker。条带1~7所对应EMA 浓度分别为 0、2、4、6、8、10 和 12 μg/mL。第 8 泳道为阴性对照(不加模板DNA)。(A'):为(A)凝胶电泳图上相应DNA条带的相对荧光强度。
2.4 完全抑制死细胞PCR扩增的最小EMA浓度
随着EMA浓度的不断增加,PCR扩增条带的亮度越来越弱,当EMA的终浓度等于或者大于1.2 μg/mL时,电泳图谱上没有相应的条带出现,即热致死细胞的PCR扩增被完全抑制(P<0.01)(图5)。此时,1.2 μg/mL的EMA浓度小于可以抑制活细胞PCR扩增的EMA浓度2 μg/mL,因此,为了能够使死细胞的PCR彻底被EMA抑制,同时又不会抑制活细胞的PCR扩增,实验中选择EMA浓度1.4 μg/mL为完全抑制死细胞PCR扩增时EMA的最适浓度。此时,通过EMA-PCR完全能够区分海产品中副溶血弧菌死活细胞。
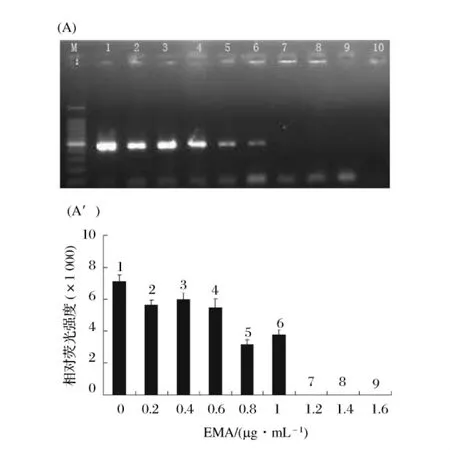
图5 完全抑制死菌细胞DNA扩增的最小EMA添加量的优化(A):不同EMA浓度下死菌细胞EMA-PCR产物的琼脂糖凝胶电泳图。M为DNA Marker。条带1~9所对应的EMA 浓度分别为 0、0.2、0.4、0.6、0.8、1.0、1.2、1.4 和1.6μg/mL。第10泳道为阴性对照(不加模板 DNA)。(A'):为(A)凝胶电泳图上相应DNA条带的相对荧光强度。
2.5 相对荧光强度与副溶血弧菌死活细胞混合液中活细胞对数的线性关系
定量的死菌细胞与变化量的活菌细胞的混合液中,死细胞DNA的PCR扩增被EMA(1.4 μg/mL)完全抑制,DNA条带的相对荧光强度随着活细胞数(2×105、2 × 104、2 × 103、1 × 103、2 × 102、1 × 102、2 ×101、和1×101CFU/PCR)的减少而逐渐减弱(图6A),检测限能达到10CFU/PCR。而且,由图6A'可知,在1×101到2×105CFU/PCR体系范围内,DNA条带的相对荧光强度与每个PCR反应体系中所存在的活细胞菌落数的对数值呈现线性关系,在此范围内可以相对荧光强度来对每个PCR体系中的活菌数进行定量。
3 结论
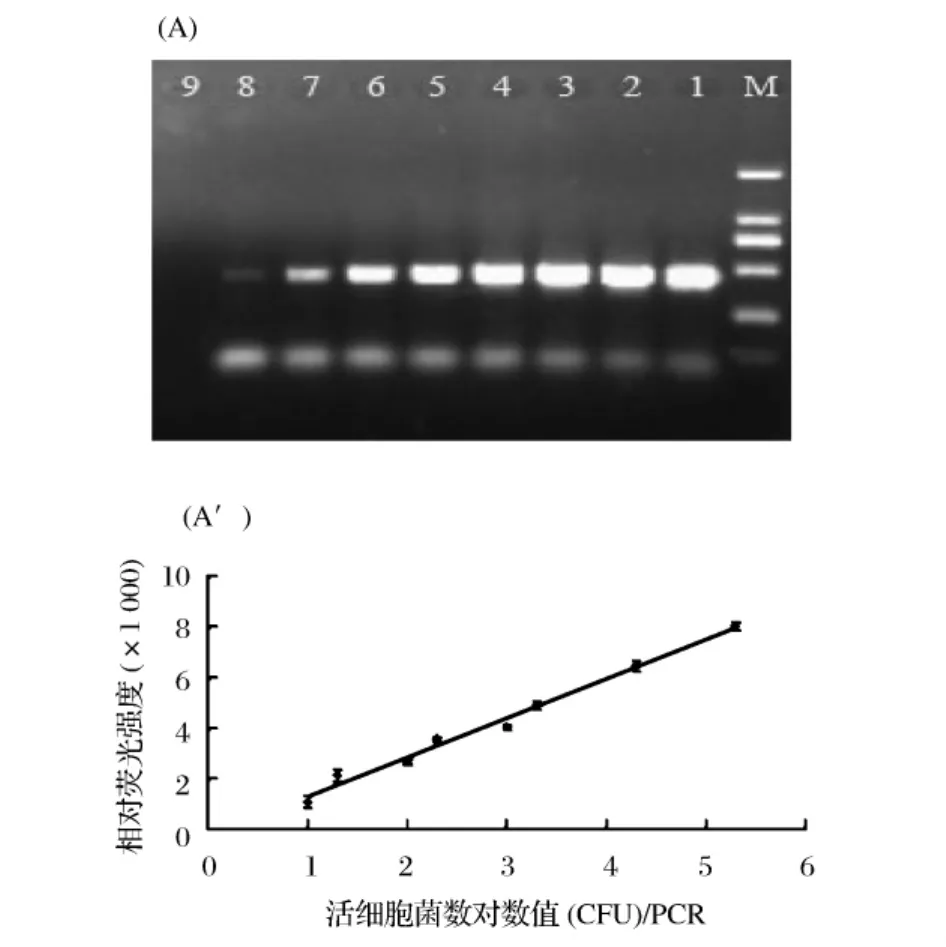
图6 DNA条带相对荧光强度与死活细胞混合液中活细胞对数值的关系(A):变化数量的活细胞(5×107、5 ×106、5 ×105、2.5 ×105、5×104、2.5 ×104、5 ×103和 2.5 ×103CFU)与固定数量(5×107CFU)死活细胞混合液的EMA-PCR扩增结果的琼脂糖凝胶电泳图,EMA终浓度均为1.4 μg/mL。1~8泳道中,每个PCR体系活细胞数分别为2×105、2×104、2 × 103、1 × 103、2 × 102、1 × 102、2 × 101、和 1 ×101CFU;第9泳道为阴性对照(不加模板DNA)。M为DNA marker。(A'):相对荧光强度与死活细胞混合液中活细胞对数值的线性关系。
本文将DNA染料EMA与传统PCR技术相结合,研究了纯培养副溶血弧菌死活细胞的鉴别方法。研究结果表明,副溶血弧菌中激活和光解EMA的最佳曝光时间为20 min,与类志贺邻单胞菌(Weimin Gu and Robert E.Levin,2008)、创伤弧菌(Wang and levin,2006)、埃希氏大肠杆菌、单核细胞增生性李斯特菌以及沙门氏菌(Nogva et al,2003)的最佳曝光时间基本相同。不抑制副溶血弧菌活细胞PCR扩增的最大EMA浓度是2 μg/mL,比类志贺邻单胞菌的5 μg/mL(Weimin Gu and Robert E.Levin,2008)以及创伤弧菌的3 μg/mL(Wang and levin,2006)都要低。完全抑制副溶血弧菌死细胞PCR扩增的最小EMA浓度为1.4 μg/mL,比类志贺邻单胞菌的0.75 μg/mL(Weimin Gu and Robert E.Levin,2008)要高,但是比创伤弧菌的2.5 μg/mL(Wang and levin,2006)要低,比大肠杆菌、单核细胞增生性李斯特菌以及沙门氏菌(Nogva et al,2003)的10 μg/mL显著偏低。通过这些数据比较表明,当用EMA-PCR方法区别不同种类微生物死活细胞的时候,完全抑制死细胞PCR扩增的最适EMA浓度以及能够影响活细胞PCR扩增的最大EMA浓度都需要重新优化[7-11]。
经EMA(1.4 μg/mL)处理定量的死菌细胞与变化量的活菌细胞的混合液,死细胞DNA的PCR扩增被完全抑制,在1×101~2×105CFU/PCR范围内,活细胞DNA扩增产物的相对荧光强度与每个PCR反应体系中所存在的活细胞菌落数的对数值呈现线性关系,此标准曲线可以用来定量任何来源的样品中的细菌数。
在本实验中,副溶血弧菌细胞热致死的条件是95℃水浴加热10 min。而Weimin Gu and Robert E.Levin(2008)的类志贺邻单胞菌的热致死条件是72℃水浴10 min,Wang and Levin(2006)创伤弧菌的热致死条件是 100℃条件下5 min,在 Nogva et al(2003)的研究中,埃希氏大肠杆菌、单核细胞增生性李斯特菌以及沙门氏菌被杀死在72℃或者100℃加热5 min,或者使用其他致死介质(96%的酒精,70%的异丙醇,或者0.1‰的杀藻胺)。可见,不同的活菌细胞致死方法也可能会影响到所需要的EMA的浓度[7]。
EMA-PCR是最近新出现的一种有效地区别微生物死活细胞的方法,利用EMA-PCR能够有效地检测并快速定量副溶血弧菌死活细胞混合液中的活细胞,从而对人类食品安全检测发挥重要的作用。
[1] 董雪,王秋雨,金莉莉,等.副溶血弧菌分子分型和检测研究进展[J].中国卫生检验杂志,2008,18(2):379-381.
[2] 王建红,王东方,陈洪永,等.水产品种副溶血弧菌的检验及方法探讨[J].中国卫生检验杂志,2008,18(2):360-361.
[3] 宋立超,樊景凤,刘述锡,等.病原性海洋弧菌快速检测方法的研究进展[J].海洋环境科学,2005,24(1):65-69.
[4] 钟凯.副溶血弧菌快速检测的进展及其应用[J].中国预防医学杂志,2005,6(1):75 -78.
[5] 李志峰,聂军,陈义忠,等.一种快速检测副溶血弧菌的PCR方法[J].解放军预防医学杂志,2004,22(6):443-444.
[6] 杨文鸽,孙翠玲,潘云娣,等.水产品中致病微生物的快速检测方法[J].中国食品学报,2006,6(1):402-406.
[7] Gu W M,Levin R E.Quantification of viable Plesiomonas shigelloides in a Mixture of viable and Dead Cells Using Ethidium Bromide Monoazide and Conventional PCR[J].Food Biotechnology,2007,21(2):145 -149.
[8] Lee J M,Levin R E.Use of ethidium bromide monoazide for quantification of viable and dead mixed bacterial flora from fish fillets by polymerase chain reaction[J].Journal of Microbiological Methods,2006,67:456-462.
[9] Nogva H,Dromtorp S,Nissen H,et al.Ethidium monoazide for DNA-based differentiation of viable and bacteria by 5'-nuclease PCR [J].Biotechniques,2003,34:804 -813.
[10] Rudi K,Moen B,Dromtorp S ,et al.Use of Ethidium Monoazide and PCR in combination for quantification of viable and dead cells in complex samples [J].Appl.Environ.Microbiol,2005,71:1 018-1 024.
[11] Wang S,Levin R.Discrimination of viable Vibrio vulnificus cells from dead cells in real-time PCR[J].J Microbiol Meth,2006,64:1-8.
[12] 冯建军,金志娟,刘西莉,等.一种DNA染料结合聚合酶链反应检测鉴别植物病原细菌死活细胞[J].高等学校化学学报,2008,29(5):944 -948.
[13] 刘晓侠,林建平,岑沛霖.微生物基因组DNA提取方法的比较与改进[J].嘉兴学院学报,2007,19(3):48-50.
[14] Abolmaaty A,Vu C,Oliver J,et al.Development of a new lysis solution for releasing genomic DNA from bacterial cells for DNA amplification by polymerase chain reaction[J].Microbios,2000,101:181-189.
[15] Yukiko H K,Tokuhiro N,HIROSHI N,et al.Improved method for detection of Vibrio parahaemolyticus in seafood[J].Applied and Environmental Microbiology,2001,67(12):5819-5823.
[16] 葛菲菲,徐锋,沈莉萍,等.副溶血弧菌PCR检测方法的建立[J].中国预防兽医学报,2008,30(3):229-232.
[17] 陈瑞英,鲁建章,苏意诚,等.食品中副溶血弧菌的危害分析、检测与预防控制[J].食品科学,2007,28(1):341-347.
[18] Nordstrom J L,Vickery M C L,Blackstone G M,et al.Development of a multiplex real-time PCR assay with an internal amplification control for the detection of total and pathogenic Vibrio parahaemolyticus bacteria in oysters[J].Applied and Environmental Microbiology,2007,73(18):5 840-5 847.
