外来入侵杂草田菁的构件生物量分配与异速生长分析
2023-07-10邓美菁张忠华莫秋丽胡聪徐超昊胡刚
邓美菁 张忠华 莫秋丽 胡聪 徐超昊 胡刚

摘要:田菁(Sesbania cannabina)是我国华南、华中和华东等地区危害严重的外来入侵杂草,通过测定其构件(根、茎、叶、花、果)生物量和植株高度等数据,分析其构件生物量分配特征及其与异速生长的关系,以期掌握该入侵杂草的生长发育规律。结果表明:田菁的构件生物量分配关系表现为茎>叶>果>根>花,而其变异系数从大到小为果>茎>叶>根>花,其地上与地下以及生殖与营养构件间的生物量均表现出极显著的差异性。此外,田菁各构件之间以及株高与构件间均为异速生长关系,株高与各构件生物量间呈极显著正相关性。田菁种群各构件间的异速生长关系以及较高的地上构件生物量与其较强的入侵性有密切联系,该结果对评估田菁的入侵机制具有重要价值。
关键词:田菁;外来入侵杂草;构件生物量分配;异速生长
中图分类号:Q944;S451文献标志码:A文章编号:1003-935X(2023)01-0035-08
Biomass Partitioning in Modules and Allometric Growth of Alien Invasive Weed Sesbania cannabina
DENG Mei-jing, ZHANG Zhong-hua, MO Qiu-li, HU Cong, XU Chao-hao, HU Gang
(School of Environmental and Life Sciences,Nanning Normal University,Nanning 530100,China)
Abstract:Sesbania cannabina is a serious alien invasive weed in South China,Central China and East China. This study measured the data such as biomass partitioning in modules (root,stem,leaf,flower and fruit) and plant height,analyzed the characteristics of biomass partitioning in modules and its relationship with allometric growth so as to understand the regulation of growth and development of this alien invasion weed. Results showed that biomass partitioning in modules presented with stem>leaf>fruit>root>flower,and their coefficients of variation were fruit>stem>leaf>root>flower. Meanwhile,there were very significant difference between biomass partitioning in reproductive and nutritional modules or aboveground and underground modules (P<0.01). In addition,all the biomass partitioning in modules were the allometric growth relationships,so were they and plant heightwith highly significant positive correlation (P<0.01). Allometric growth and higher biomass in underground modules of S. cannabina were closely related to its strong invasiveness. The results was of great value to evaluate the invasion mechanism of S. cannabina.
Key words:Sesbania cannabina;alien invasive weed;biomass partitioning in modules;allometric growth
收稿日期:2022-07-31
基金項目:广西自然科学基金(编号:2021GXNSFFA196005、2021GXNSFAA196024);国家自然科学基金(编号:31960275、31760128)。
作者简介:邓美菁(2002—),女,湖南衡阳人,主要从事外来入侵植物方面的研究。E-mail:3320427468@qq.com。
通信作者:胡 刚,博士,副教授,主要从事植被生态学研究。E-mail:ahhugang@126.com。
外来入侵物种会对入侵地的生态系统构成一定的威胁,可通过物种替代的方式改变入侵地生态关键过程,这已成为全球生物丧失多样性的重要因素之一[1]。外来物种入侵已对全球生态、经济和社会造成诸多负面影响,已成为政府、民众和学者高度关注的生态环境问题,因而监测和研究入侵物种、防控其传播扩散显得尤为重要[2-3]。就外来入侵植物而言,其多为繁殖速度快、适应能力强的物种,侵占入侵地土著植物的生存空间并排挤其土著植物,对入侵地土著植物多样性造成一定的危害[4]。伴随我国经济的快速发展,外来植物入侵问题越来越严重,已造成了不可估量的经济损失[5]。最新研究表明,我国共有归化植物1 099种,隶属于112科578属[6],其中外来入侵植物达72科285属515种,这些外来入侵植物尤以菊科、豆科和禾本科为主,生活型以草本为主[7]。一些适应能力强的外来入侵植物如紫茎泽兰(Ageratina adenophora)、加拿大一枝黄花(Solidago canadensis)、喜旱莲子草(Alternanthera philoxeroides)、互花米草(Spartina alterniflora)等已对我国陆地或水域生态系统造成了严重的影响。
田菁(Sesbania cannabina)是豆科(Fabaceae)田菁属(Sesbania)的一年生亚灌木状草本植物,株高可达3 m,原产于澳大利亚至西南太平洋岛屿,归化于亚洲、非洲部分地区以及印度洋岛屿,早期作为绿肥植物被有意引入我国,现已入侵至我国的广东、广西、湖南、湖北、上海、浙江等18个省(市),属于严重入侵类的外来植物[7]。田菁适应性强,耐盐性、耐涝性、耐旱性、抗逆性和抗病虫害能力也较强,可快速逸生为习见的高大杂草,常在荒地、弃耕地以及人为扰动较大的区域形成优势种群,排挤土著植物[8]。迄今,针对田菁的叶绿体全基因组测序[9]、种子萌发的生物学特征[8]、营养器官的解剖结构特征[10]、抗逆性[11]等方面都已有了一定的研究。然而,与紫茎泽兰、加拿大一枝黄花、喜旱莲子草等外来入侵植物相比,同样作为一种对我国陆域生态系统造成严重入侵的外来物种,除有学者初步总结了其在我国的分布区范围外[6],几乎无其入侵机制方面的研究报道,不利于其防控措施的制定。
植物生物量的分配策略是植物对环境适应的结果,与植物的生长发育息息相关,反映出植物在各构件生物量分配中的权衡关系[12-13]。有学者对外来入侵植物进行了构件生物量分配格局与异速生长分析,例如,张桥英等探讨了增温对入侵植物马缨丹(Lantana camara)生物量分配和异速生长的影响,发现温度升高有利于其同化作用的增强,从而提高其竞争力[14];闫小红等分析了不同生育期入侵植物大狼耙草(Bidens frondosa)的生物量分配格局及异速生长,发现其生长特性、生物量分配格局及异速生长关系在不同生育期间存在较大差异,体现了其灵活的生长和资源分配策略[15];Rayamajhi等构建了佛罗里达州外来入侵藤本植物黄独(Dioscorea bulbifera)的异速生长模型,该模型为估计该入侵植物生物量组成和营养繁殖体密度提供了非破坏性的方法[16]。另有研究表明,异速生长和干扰之间的相互作用可能是喜旱莲子草成功入侵的部分原因[17]。此外,对银胶菊(Parthenium hysterophorus)[18-19]、飞机草(Chromolaena odorata)[20]、牛膝菊(Galinsoga parviflora)[21]等入侵植物均进行了构件生物量分配格局或构件异速生长方面的研究。上述学者从构件生物量分配和异速生长视角分析不同外来入侵植物的生长发育规律,有助于深入探讨这些物种的入侵机制。然而,对于已在我国中东和华南地区造成严重入侵后果的外来杂草田菁仍未开展过相关研究,尚不清楚其构件生物量的分配规律及其与入侵能力间的关系。
广西南宁的荒地、弃耕地和公共绿地等开阔生境往往分布有较高优势度的田菁种群,在较大程度上抑制了土著植物的生长,并造成较为明显的土著生物多样性丧失。为分析田菁的生长发育规律,本研究通过分析田菁根、茎、叶、花、果的生物量分配情况并比较各构件(包括地上与地下构件、营养与生殖构件)生物量间及其与株高间的异速生长特征,解析田菁种群各构件生物量之间的分配特征和异速生长关系,以期为探究其入侵机制和防控种群扩散提供科学依据。
1 材料与方法
1.1 研究区概况
南宁市(22°13′~23°32′N,107°45′~108°51′E)位于广西壮族自治区的南部,北回归线南侧,属湿润的亚热带季风气候,阳光充足,雨量充沛,霜少无雪,气候温和,夏长冬短。年均气温约21.6 ℃,年均降水量达1 304.2 mm,平均相对湿度为79%。南宁地区的地貌主要有平地、低山、石山、丘陵和台地5种类型,地带性土壤类型为红壤,地带性植被为亚热带常绿阔叶林。良好的地理和气候条件孕育了丰富的植物资源,已知有维管束植物3 000余种,隶属于209科764属[22]。
1.2 测定方法
2020年8月,选取南宁城市荒地中正处于花果期的田菁种群并对其进行取样。随机选取40株大小不等的田菁成熟植株,用钢卷尺测量其株高,然后对每株田菁地上部分齐地面进行剪割,剪取后用枝剪分离花、果实、叶以及茎秆,将其分别装入信封,称取鲜重,再带回实验室备测干重;采用挖掘法连同土壤一起挖出所有根系,编号后带回实验室,洗净根上的泥土并称取鲜重。将田菁各部分构件(根、茎、叶、花、果)放入105 ℃烘箱中杀青15 min,然后75 ℃烘至恒重,称取干重并记录各构件的生物量数据。
1.3 数据分析
首先对田菁的各构件生物量进行描述性统计分析,然后对各构件生物量数据进行显著性检验(t检验和方差分析)和相关性分析,再对各构件的生物量与株高数据进行拟合分析,最后对不同拟合模型的相关系数作显著性检验。图表中数据为“平均值±标准差”,采用SPSS 22.0统计分析软件进行数据分析。
为分析不同构件间的异速生长关系,不同构件生物量用方程y=bxa来描述,线性转换成ln y=ln b+a(ln x),式中:x和y分別为不同生物量数据;b为构件生物量关系的截距;a为两者关系的斜率,也就是相对生长的指数,a=1时为等速生长,a≠1时为异速生长。分析方法采用标准化主轴估计(standardized major axis estimation,SMA)法,各参数的确定均在软件SMATR[23]中完成。
2 结果与分析
2.1 田菁构件间的生物量分配
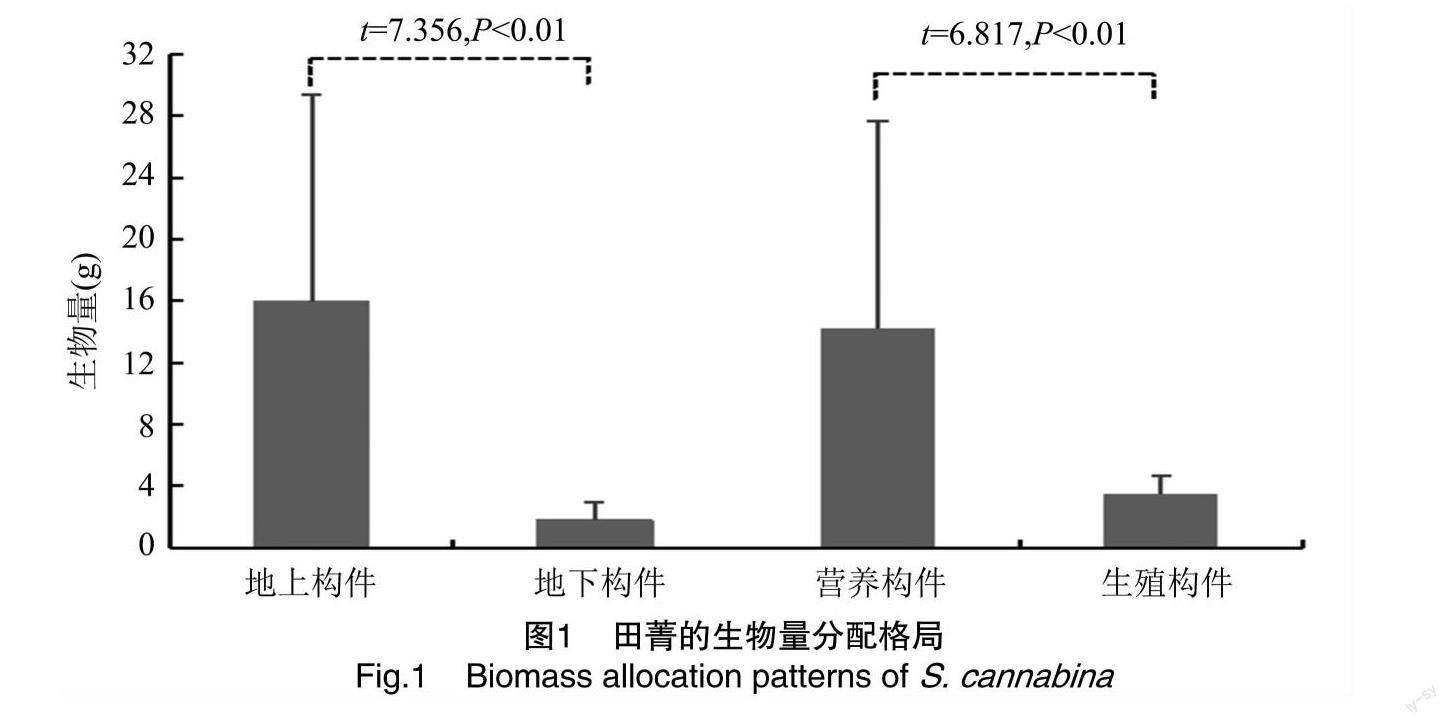
由描述性统计分析结果(表1)可知,田菁种群各构件生物量大小关系为茎>叶>果>根>花,变异系数(coefficient of variation,CV)为51.28%~114.86%,其中根与茎、叶、花之间生物量差异显著(P<0.05),花与茎、叶、果和根之间差异显著(P<0.05)。田菁株高为43.30~180.40 cm,CV为39.75%。田箐总生物量为3.54~61.85 g,平均值为17.71 g,CV为81.82%。由图1可知,田菁种群的生物量分配关系表现为地上构件>地下构件、营养构件>生殖构件,且地上与地下构件、营养与生殖构件间均存在极显著差异(P<0.01)。
2.2 田菁构件生物量间的异速生长关系
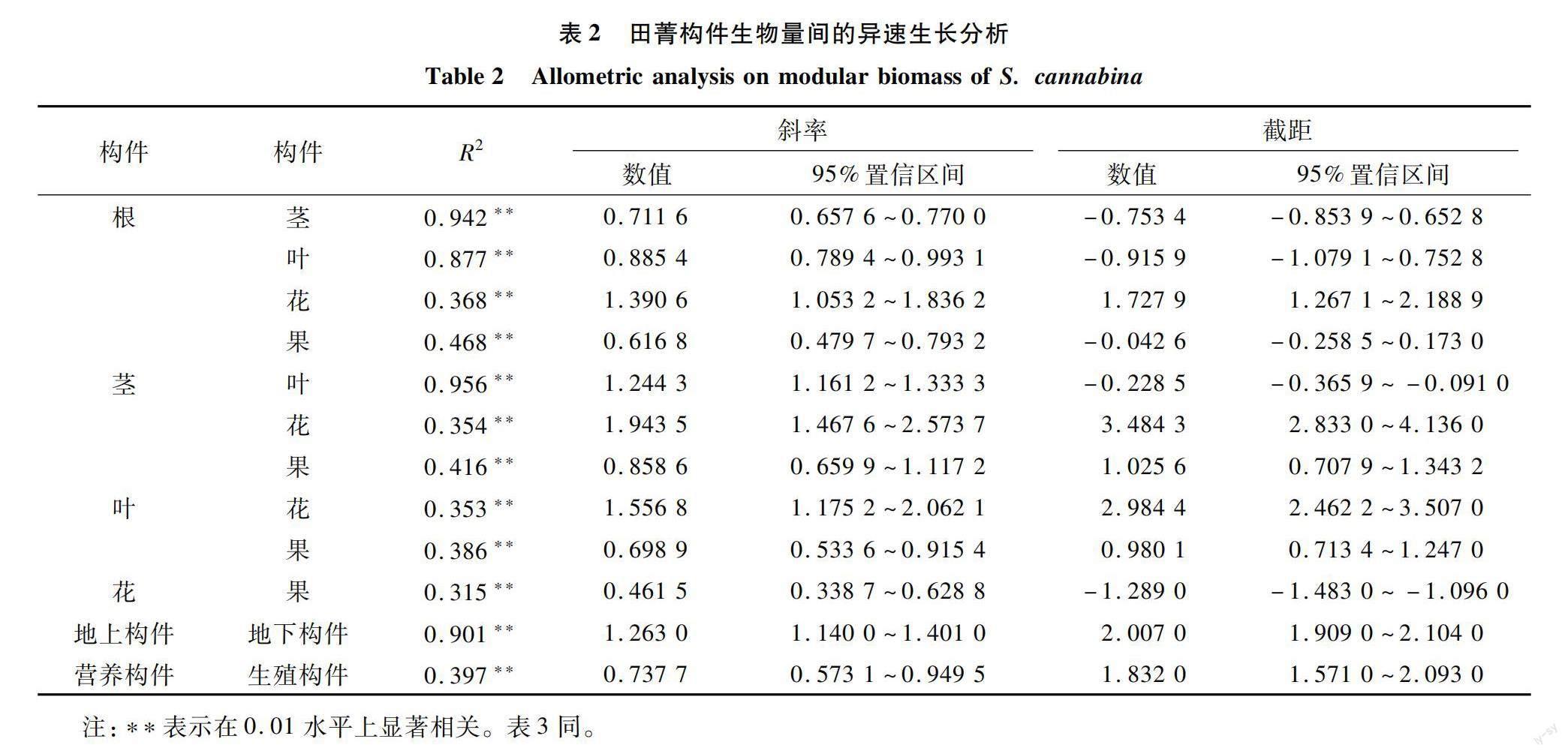
如表2所示,田菁各构件生物量间均为异速生长关系(a≠1)。地上构件与地下构件生物量差异极显著(P<0. 01),斜率为1.263[95%置信区间(CI)=1.140~1.401],表明地上构件与地下构件之间不成比例地生长。随着田菁地下构件生物量的增加,地上构件生物量的增加速度变缓。同时,营养构件与生殖构件生物量也呈极显著关系(P<0.01),斜率为0.737 7(CI=0.573 1~0.949 5);随着生殖构件生物量的增加,营养构件根、茎、叶的增加速度变缓。地上构件与地下构件、营养构件与生殖构件、根、茎、叶、花、果之间均表现出异速生长关系(a≠1)。
2.3 田菁株高与构件生物量间的拟合与异速生长关系
回归分析和相关性检验(图2)表明,在田菁繁殖生长期间,营养器官根、茎、叶的生物量和繁殖器官花、果实的生物量都与株高存在极显著的正相关关系(P<0.01),根、茎、叶、花和果生物量都随着株高的增长而提高。株高与总生物量、叶生物量和果生物量间可用幂函数较好拟合(图2-a、图2-d、图2-f),株高与根和茎的生物量间可用对数函数拟合(图2-b、图2-c),而株高与花生物量间利用线性函数较好拟合(图2-e)。此外,田菁株高与总生物量、各构件生物量间均表现为异速生长关系(a≠1)(表3)。
3 讨论与结论
3.1 田菁的构件生物量结构及其分配特征
生物量是植物的基本生物学特征和功能性状之一,其在各构件上的分配受遗传特性和环境因素共同作用,反映了植物的物质积累状况和对环境资源利用的能力[24]。茎和叶是植物合成、储存和运输营养物质的重要构件,所以其生物量投入往往最大[25]。本研究结果表明,田菁种群的茎和叶生物量占比分别为36.82%和32.85%,说明田菁将近70%的生物量分配给能获取资源的营养构件。Tilman认为,一年生植物通常将更多的资源分配给地上部分,而多年生植物则将更多的资源分配给地下部分[26]。本研究表明,一年生植物田菁具有较高的地上构件生物量和较低的地下构件生物量,进一步印证了上述观点。田菁较低的根冠比(0.11),表明其较小的地下构件投入便能支持较大的地上构件生长发育。繁殖是生物繁衍后代以及延续种族最基本的行为和过程,也是生物生活史过程中最为重要的一环,植物种群的更新和种群中植物个体生活史的完成均依赖于其繁殖过程[27]。植物在生殖期,其植株需要将部分资源分配给生殖构件,这势必减少对包括根等其他构件的分配。田菁各构件生物量的变异系数为51.28%~114.86%,与入侵能力强的小飞蓬的生物量变异系數(99.69%~107.48%)[28]相比,其各构件的表型可塑性可能更强。田菁各构件生物量分配比的变异系数大小排列为果>茎>叶>根>花,表明果、茎等构件生物量可塑性可能更强。
最优分配理论(optimal partitioning theory)认为,植物倾向于将生物量分配给能够获取稀缺资源的器官以最大化适应外界环境的变化[29],也有研究认为生物量分配是由植物本身的生长模式可塑性主导的[30]。田菁在其入侵的群落中会形成高密度的种群,密度的增大可引起多种资源的变异,涉及复杂的种内相互作用(包括促进作用和竞争作用)。有研究表明,个体大小的增加使植物间相互作用(同时有地上构件和地下构件相互作用)增强,并逐渐开始发生竞争作用,贫瘠的土壤中资源匮乏加剧了植物地下构件间的竞争强度,导致根生物量显著降低[31]。随着植株的生长,贫瘠土壤中高密度田菁种群倾向于降低根生物量和根冠比,这可能是其地下构件生物量较低的原因之一。然而,田菁种群密度增加与其生境适应策略关系仍需进一步研究。
3.2 田菁的构件生物量分配与其入侵能力的关系
外来植物的入侵性与植物本身的遗传机制和对环境的适应能力有关[32-34]。在入侵地天敌的缺乏、资源利用率高、生长竞争力强、繁殖能力强和生态幅广等因素均决定了外来植物的入侵性。田菁是一年生草本植物,其植株高大(平均株高为93.46 cm),在群落中与低矮的土著植物相比更容易获取光照,从而影响土著植物的生长发育。田菁叶片生物量占比达32.85%,有利于田菁把更多的光能转变为化学能,促进田菁的快速生长,而果在总生物量中的占比为17.74%,表明田菁具有较高的种子产量。田菁在长达半年的花果期内可边开花,边结果,边传播种子,边萌发形成新个体,这一特性有利于田菁群落抢占更多的生存资源和空间,从而具有更强的繁殖竞争能力。Wang等比较分析了入侵植物与土著植物的性状差异,表明入侵植物的株高、根冠比和繁殖分配系数等显著大于土著植物,这些在其成功入侵进程中可能起重要作用[33]。Cheng等研究发现,入侵植物加拿大一枝黄花生长过程中会将更多的生物量分配给繁殖器官,从而有助于其成功入侵[27]。本研究同样表明,随着田菁株高的增加,繁殖构件(花和果)生物量出现显著增加的趋势,更高的繁殖生物量分配表明其种群更新能力可能更强。
植物各构件生物量与总生物量以及不同构件生物量间的关系通常表现为异速生长关系,这种关系广泛存在于植物中,能够揭示植物构件生物量分配生物学特征间的内在规律[29]。异速生长被认为是表型可塑性的特征之一,是影响植物生物量分配的关键因素[35-36]。本研究结果表明,在田菁繁殖期间,其各构件生物量、地上构件与地下构件以及营养构件与生殖构件间均呈异速生长关系。因此,田菁在繁殖期其构件生物量的分配较为灵活,能够更好地适应环境变化,同时又可以为其更高效的繁殖提供保障,增强了其入侵性。对植物构件异速生长关系及其变化进行研究,有助于了解植物构件对环境的适应性响应,植物构件之间多具有显著的相关性和异速生长关系,体现了构件之间的协同生长策略[13]。不同构件间异速生长关系存在差异,这在马缨丹[14]、大狼耙草[15]、喜旱莲子草[17]等外来入侵植物中也得到了普遍验证。田菁各构件生物量间及其与株高间均表现为显著的异速生长关系,同样表明其能较好地调节植物的生长速率,使其具有更强的适应性和入侵能力。
多样性阻抗假说 (diversity resistance hypothesis) 认为,群落的生物多样性对抵抗外来种的入侵起着关键性作用,越是物种多样且结构复杂的生态系统,其空余生态位越少而对入侵种的抵抗能力越强[37]。外来植物的入侵能力在很大程度上取决于生境中可利用的非生物资源如光照、水分、养分等[38]。 田菁往往入侵荒地、弃耕地和公共绿地等人为干扰严重的开阔生境,笔者建议在这些生境中以乡土树种为主进行植物功能群配置,从而实施生态替代措施,利用乡土植物对光照、土壤养分等资源的竞争优势来减少空余生态位,从而阻碍阳性植物田菁的生长和种群扩散,这样既降低了入侵植物的競争能力,又提高了生境对入侵植物的抵抗力。
本研究对外来入侵植物田菁种群进行生物量分配和异速生长关系分析,结果表明,田菁不同构件间生物量及其分配具有显著的差异,这些差异来源于构件间或构件与个体大小之间不同的相对生长速率。田菁各构件生物量分配表现为茎>叶>果>根>花,且地上构件生物量显著大于地下构件,营养构件生物量显著大于生殖构件。田菁的株高与各构件生物量呈极显著的正相关关系,各构件之间以及株高与构件间均呈现异速生长关系,表明田菁在生物量分配时具有灵活性,可增强其对环境的适应性,从而提高其入侵能力。本研究从其生物量分配和异速生长关系角度揭示了外来杂草田菁在花果期的生长特征,有助于进一步理解其环境适应策略和入侵机制,并为其防控提供科学指导。然而,目前对田菁入侵机制的理解还较为有限,进一步的研究需要关注田菁原产地和入侵地的表型可塑性差异及其固氮能力与其入侵性的关系。
参考文献:
[1]Doherty T S,Glen A S,Nimmo D G,et al. Invasive predators and global biodiversity loss[J]. Proceedings of the National Academy of Sciences of the United States of America,2016,113(40):11261-11265.
[2]Kopf R K,Nimmo D G,Humphries P,et al. Confronting the risks of large-scale invasive species control[J]. Nature Ecology & Evolution,2017,1(6):172.
[3]Luque G M,Bellard C,Bertelsmeier C,et al. The 100th of the world's worst invasive alien species[J]. Biological Invasions,2014,16:981-985.
[4]Paini D R,Sheppard A W,Cook D C,et al. Global threat to agriculture from invasive species[J]. Proceedings of the National Academy of Sciences of the United States of America,2016,113(27):7575-7579.
[5]Horvitz N,Wang R,Wan F H,et al. Pervasive human-mediated large-scale invasion:analysis of spread patterns and their underlying mechanisms in 17 of China's worst invasive plants[J]. Journal of Ecology,2017,105(1):85-94.
[6]许光耀,李洪远,莫训强,等. 中国归化植物组成特征及其时空分布格局分析[J]. 植物生态学报,2019,43(7):601-610.
[7]闫小玲,刘全儒,寿海洋,等. 中国外来入侵植物的等级划分与地理分布格局分析[J]. 生物多样性,2014,22(5):667-676.
[8]Iqbal N,Manalil S,Chauhan B S,et al. Germination biology of sesbania (Sesbania cannabina):an emerging weed in the Australian cotton agro-environment[J]. Weed Science,2019,67(1):68-76.
[9]Su C,Chen H F,Chang Z Y,et al. The complete chloroplast genome of Sesbania cannabina (Fabaceae) from China[J]. Mitochondrial DNA (Part B:Resources),2020,5(2):1890-1891.
[10]吴 晓,周金鑫,翁 乐,等. 田菁营养器官的解剖结构特征[J]. 亚热带植物科学,2018,47(2):128-133.
[11]Srivastava N,Kumar G.Influence of drought stress on cytological behavior of green manure crop Sesbania cannabina Poir.[J]. Cytologia,2014,79(3):325-329.
[12]Poorter H,Nagel O.The role of biomass allocation in the growth response of plants to different levels of light,CO2,nutrients and water:a quantitative review[J]. Functional Plant Biology,2000,27(6):595.
[13]Liu R,Yang X J,Gao R R,et al. Allometry rather than abiotic drivers explains biomass allocation among leaves,stems and roots of Artemisia across a large environmental gradient in China[J]. Journal of Ecology,2021,109(2):1026-1040.
[14]張桥英,彭少麟. 增温对入侵植物马缨丹生物量分配和异速生长的影响[J]. 生态学报,2018,38(18):6670-6676.
[15]闫小红,何春兰,周 兵,等. 不同生育期入侵植物大狼杷草的生物量分配格局及异速生长分析[J]. 生态与农村环境学报,2017,33(2):150-158.
[16]Rayamajhi M B,Rohrig E,Tipping P W,et al. Allometric equations for the invasive vine air potato (Dioscorea bulbifera) in its exotic range in Florida[J]. Invasive Plant Science and Management,2020,13(2):76-83.
[17]Jia X,Pan X Y,Li B,et al. Allometric growth,disturbance regime,and dilemmas of controlling invasive plants:a model analysis[J]. Biological Invasions,2009,11:743-752.
[18]蒲高忠,唐赛春,潘玉梅,等. 入侵植物银胶菊在不同生境下表型可塑性和构件生物量[J]. 广西植物,2010,30(5):641-646,625.
[19]Rathee S,Ahmad M,Sharma P,et al. Biomass allocation and phenotypic plasticity are key elements of successful invasion of Parthenium hysterophorus at high elevation[J]. Environmental and Experimental Botany,2021,184(1):104392.
[20]Beest M T,Stevens N,Olff H,et al. Plant-soil feedback induces shifts in biomass allocation in the invasive plant Chromolaena odorata[J]. Journal of Ecology,2009,97(6):1281-1290.
[21]齐淑艳,徐文铎,文 言. 外来入侵植物牛膝菊种群构件生物量结构[J]. 应用生态学报,2006,17(12):2283-2286.
[22]黄良美,黄玉源,黎 桦,等. 南宁市植物群落结构特征与局地小气候效应关系分析[J]. 广西植物,2008,28(2):211-217.
[23]Falster D S,Warton D I,Wright I J. SMATR:standardised major axis tests and routines,Version 2.0[DB/OL]. [2022-05-15]. http://www.bio.mq.edu.au/ecology/SMATR/.
[24]陶 冶,张元明. 准噶尔荒漠6种类短命植物生物量分配与异速生长关系[J]. 草业学报,2014,23(2):38-48.
[25]Poorter H,Niklas K J,Reich P B,et al. Biomass allocation to leaves,stems and roots:meta-analyses of interspecific variation and environmental control[J]. New Phytologist,2012,193(1):30-50.
[26]Tilman D. Plant strategies and the dynamics and structure of plant communities[M]. Princeton:Princeton University Press,1988.
[27]Cheng H Y,Wang S,Wei M,et al. Reproductive allocation of Solidago canadensis L. plays a key role in its invasiveness across a gradient of invasion degrees[J]. Population Ecology,2021,63(4):290-301.
[28]潘玉梅,唐赛春,蒲高忠,等. 外来入侵植物小飞蓬种群构件生物量结构特征[J]. 广西植物,2009,29(4):476-480.
[29]Reich P B,Luo Y J,Bradford J B,et al. Temperature drives global patterns in forest biomass distribution in leaves,stems,and roots[J]. Proceedings of the National Academy of Sciences of the United States of America,2014,111(38):13721-13726.
[30]McCarthy M C,Enquist B J. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation[J]. Functional Ecology,2007,21(4):713-720.
[31]Wang S,Li L,Zhou D W. Root morphological responses to population density vary with soil conditions and growth stages:the complexity of density effects[J]. Ecology and Evolution,2021,11(15):10590-10599.
[32]陳 超,滕文军,武菊英. 细茎针茅的园林应用、生物学和生态学特性及风险评估[J]. 杂草学报,2022,40(1):1-7.
[33]Wang C Y,Zhou J W,Liu J,et al. Differences in functional traits and reproductive allocations between native and invasive plants[J]. Journal of Central South University,2018,25(3):516-525.
[34]胡沁沁,周 颖,华春霞,等. 浙江农林大学校园外来入侵植物及其入侵风险等级评估[J]. 杂草学报,2021,39(2):28-35.
[35]熊韫琦,赵彩云. 表型可塑性与外来植物的成功入侵[J]. 生态学杂志,2020,39(11):3853-3864.
[36]Weiner J. Allocation,plasticity and allometry in plants[J]. Perspectives in Plant Ecology,Evolution and Systematics,2004,6(4):207-215.
[37]Elton C S. The ecology of invasions by animals and plants[M]. Berlin:Springer Science+Business Media,1958.
[38]廖慧璇,周 婷,陈宝明,等. 外来入侵植物的生态控制[J]. 中山大学学报(自然科学版),2021,60(4):1-11.
