植物激素对水稻籽粒灌浆、粒质量与品质的调控作用研究进展
2023-05-29刘洋肖文惠蔡文璐张伟杨王志琴徐云姬
刘洋 肖文惠 蔡文璐 张伟杨 王志琴 徐云姬
(扬州大学教育部农业与农产品安全国际合作联合实验室/江苏省粮食作物现代产业技术协同创新中心,江苏扬州 225009;第一作者:19736 39167@qq.com;*通讯作者:yunjixu@yzu.edu.cn)
水稻是重要的粮食作物之一,我国有60%以上的人口以稻米为主食[1]。随着我国经济社会的发展与人们消费能力的提高,稻米品质愈加受到重视[2-3]。水稻遗传育种和栽培调控的目标已由单纯追求高产逐渐转向优质高产[4-5]。籽粒灌浆期是水稻生长发育的关键阶段,对粒质量和稻米品质形成具有决定作用。因此,阐明水稻籽粒灌浆特性及其机理,对促进籽粒灌浆、提高粒质量和品质具有重要意义。
植物激素或称植物生长物质,是一类由植物自身产生且在极低浓度下有明显生理效应的微量有机物[6]。目前,植物激素有生长素类(IAA)、赤霉素类(GAs)、细胞分裂素类(CTK)、脱落酸(ABA)和乙烯(ETH)5 类经典激素,及油菜素甾醇类(BRs)、多胺(PAs)、茉莉酸类(JA)、水杨酸(SA)和独脚金内酯(SL)等新型植物激素[7]。大量研究表明,植物激素对水稻籽粒灌浆、粒质量及品质具有重要调控作用,而且不同植物激素发挥的作用及其机制不同。本文在介绍水稻籽粒灌浆增重特性及其与稻米品质关系的基础上,重点综述了水稻灌浆籽粒中不同植物激素含量的变化特点,及其对籽粒灌浆、粒质量和品质的调控作用及其机理研究进展,旨在为水稻优质高产的遗传改良和栽培调控提供理论依据。
1 水稻籽粒灌浆增重特性与稻米品质
1.1 籽粒灌浆增重特性
水稻籽粒灌浆是水稻生长发育的关键阶段,其灌浆物质主要来源于花前茎秆(含鞘)中的非结构性碳水化合物和花后叶片的光合同化物[8]。通常用Richards 方程W=A/(1+Be-kt)1/N 拟合籽粒灌浆增重动态,通过该方程及其变式可推导出平均灌浆速率(Gmean)、最大灌浆速率(Gmax)、起始生长势(R0)、活跃灌浆期(D)和灌浆持续时间(T)等灌浆相关参数[9]。
水稻不同位置籽粒的灌浆增重具有明显的时空差异性,且在大穗型水稻品种中尤为突出。位于稻穗中上部一次枝梗上的颖果一般开花较早,灌浆速率快,结实率高,粒质量高,称为强势粒;而稻穗基部二次枝梗上的颖果开花较迟,灌浆速率慢,结实率低,粒质量低,称为弱势粒[10]。关于水稻强、弱势粒灌浆差异,目前主要归因于同化物供应限制[11]、蔗糖-淀粉代谢途径关键酶活性低[10]、同化物转运效率低[12-13]、库容及库活性限制[14-16]等。根据强、弱势粒开花时间的差异,水稻籽粒灌浆类型可分为同步灌浆型和异步灌浆型[17]。同步灌浆型中,强、弱势粒灌浆启动时间相近,各灌浆参数差异较小;而异步灌浆型中,强、弱势粒灌浆启动时间以及各灌浆参数差异显著[17]。研究表明,水稻弱势粒较低的灌浆速率和结实率是制约其粒质量及产量提高的重要原因[18]。
1.2 稻米品质
稻米品质主要包括加工品质、外观品质、蒸煮与食味品质、营养品质和卫生品质,且不同品质类型的指标各不相同(图1)。其中,蒸煮与食味品质最受人们关注,衡量其优劣的指标多与淀粉理化性质有关[19-20]。据报道,直链淀粉含量较高的稻米质地干松、色泽较淡、胀性大、黏性小、冷却后易变硬,而含量低的米粒则富有光泽、胀性小、黏性高[21-22]。对于优质大米的评判,籼稻和粳稻品种的标准不同,目前我国已出台了具体的数值指标[23]。
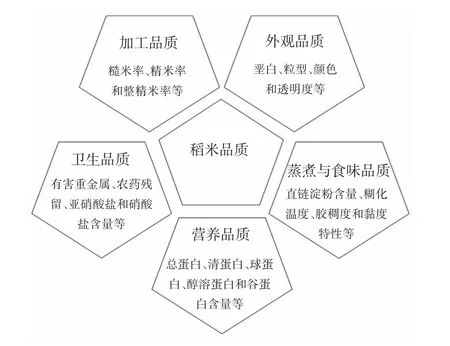
图1 稻米品质的类型及其衡量指标
1.3 籽粒灌浆增重特性与稻米品质的关系
水稻籽粒灌浆增重过程对稻米品质形成至关重要。有学者指出,播期延迟会导致向籽粒运输的灌浆物质减少,整精米率下降,加工品质变劣[24]。籽粒灌浆充实不良也会引起稻米垩白形成,如高温下籽粒灌浆速率过快,淀粉结构松散[25]、气隙较大[26],同时有效灌浆时间缩短,有机物积累不足[27],会导致垩白增大,透明度下降,严重损害稻米外观品质。随着灌浆期温度的升高,直链淀粉含量、较短支链淀粉比例也会降低[28],相对结晶度、糊化温度和糊化焓值升高[28-30]。此外,氮肥后移[31]、优化氮素管理[31-32]和轻-干湿交替灌溉[33]等栽培措施对稻米淀粉理化特性具明显改善作用。
2 水稻灌浆籽粒中植物激素含量的变化特点
2.1 经典植物激素含量的动态变化
在水稻灌浆进程中,籽粒吲哚-3-乙酸(IAA)、玉米素(Z,一种天然细胞分裂素)、玉米素核苷(ZR,也是一种天然细胞分裂素)和脱落酸(ABA)的含量呈明显的倒“V”形单峰曲线变化趋势[34-36]。研究显示,IAA、Z、ZR 和ABA 含量在水稻强势粒中到达峰值的时间比弱势粒要早,灌浆前期强势粒中上述植物激素含量高于弱势粒,灌浆后期则相反[34-35,37]。但是,上述激素含量在水稻一些突变体材料的强、弱势粒间差异并不明显[38]。
灌浆籽粒中赤霉素类(GAs)因其种类较多,含量变化相对复杂。GA3含量一般呈倒“V”形单峰曲线或逐渐降低的变化趋势[17,36],GA4和GA7均呈先降低后升高、再降低的变化趋势[40]。而且,水稻强势粒的GAs 含量高于弱势粒[39-40]。
乙烯(ETH)是一种重要的气态植物内源激素,其含量无法直接测定,常用1-氨基环丙烷-1-羧酸(ACC,一种乙烯合成前体)含量和乙烯释放速率(EER,ethylene evolution rate)表示[41]。水稻籽粒的EER在灌浆初期很高,随着灌浆进程推进总体呈下降趋势[42],ACC 含量变化与EER 类似[43]。与其他4 种经典激素不同的是,水稻弱势粒的EER 和ACC 含量显著高于强势粒[42-43]。
2.2 新型植物激素含量的动态变化
有关水稻灌浆籽粒新型植物激素含量变化特性的研究相对较少,目前主要集中在多胺(PAs)和水杨酸(SA)上。植物体中PAs 主要包括腐胺(Put)、亚精胺(Spd)和精胺(Spm),在水稻籽粒中多以游离态存在[44]。游离Put、Spd 和Spm 含量在籽粒灌浆过程中均呈倒“V”形单峰曲线变化,但其含量大小因多胺种类、品种不同而差异较大[43,45]。SA 含量在灌浆过程中也表现为倒“V”形单峰曲线变化,灌浆前、后期的SA 含量差异明显,其中前期强势粒中SA 含量高于弱势粒,而后期则相反[46]。
3 植物激素对籽粒灌浆和粒质量的调控作用及机理
3.1 生长素(IAA)对籽粒灌浆和粒质量的调控作用及机理
研究表明,内源IAA 含量与籽粒灌浆速率呈显著或极显著正相关[34,36,47]。IAA 通过增强蔗糖酶(Sucrase)、蔗糖合酶(SuSase)、腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)、淀粉合酶(StSase)和淀粉分支酶(SBE)等蔗糖-淀粉代谢途径关键酶活性[34-36,48],促进了籽粒灌浆及粒质量提高。王志琴等[49]认为,籽粒中IAA 含量和H+-ATP 酶活性是反映库容的2 个重要指标。籽粒灌浆过程中IAA 含量动态变化与籽粒中H+-ATP 酶(H+-ATPases)活性变化趋势基本一致,而其与IAA 氧化酶(IAA oxidase)、IAA 过氧化物酶(IAA peroxidase)的活性则呈负相关[48],表明内源IAA 可有效增加籽粒库容。
据报道,喷施外源IAA 不仅可显著提高籽粒的物质转运率、结实率和粒质量[50-51],还可缩小强、弱势粒内源IAA 含量的差异,削弱杂交稻的粒间顶端优势[47,52-53]。此外,喷施IAA 增加了籽粒胚乳细胞数目及体积[38-39,54]。而且,IAA 可参与调节与胚乳细胞壁伸展有关的β-1,3-葡聚糖酶和β-1,4-葡聚糖酶活性,进而影响胚乳体积[48]。
3.2 赤霉素类(GAs)和细胞分裂素类(CTK)对籽粒灌浆和粒质量的调控作用及机理
GAs 对籽粒灌浆和粒质量的调控作用体现在与其他植物激素的协同效应,如灌浆前、中期较低的GA1+3/ABA 值、较高的GA1+3/(IAA+ABA+GA1+3+GA4+7)值有利于籽粒结实率的提高[39,55],而单一喷施外源GA1+4对籽粒胚乳增殖速率、灌浆速率、茎鞘物质输出率、结实率和充实度等影响不大[38,55-56]。还有研究表明,GAs 一方面可促进色氨酸合成IAA,抑制IAA 氧化酶和IAA 过氧化物酶的活性[55],另一方面可增强α-淀粉酶的活性,降低AGPase 和StSase 的活性,抑制淀粉的合成与积累[56-58]。此外,王熹等[59]进一步提出,GAs 对IAA 的作用具有粒位差异性,外源喷施GA3促进强势粒中IAA 含量的提高,而对弱势粒无显著影响,导致强、弱势粒灌浆差异更大。
研究表明,籽粒灌浆前期较高的Z 和ZR 含量有利于胚乳细胞体积扩大、细胞数量增多,且Z 和ZR 含量与籽粒灌浆速率、充实率和千粒重均呈正相关[34,36]。此外,喷施外源6-BA(一种人工合成细胞分裂素)有效减缓了水稻生育后期叶片叶绿素含量、Rubisco 活性及其他蛋白质功能的丧失,延缓了叶片早衰,延长了叶片光合作用功能期,促进籽粒灌浆[60-61]。CTK 还可调节叶片氮代谢相关酶活性,提高氮利用效率,进而促进更多的氮和干物质向穗部分配[62]。
3.3 脱落酸(ABA)和乙烯(ETH)对籽粒灌浆和粒质量的调控作用及机理
据报道,ABA 对水稻籽粒灌浆的调控作用具有“双重性”。灌浆初期籽粒中低浓度的ABA 通过增强籽粒中SuSase 和AGPase 活性,进而提高水稻的结实率和产量[56,63];而灌浆后期高浓度的ABA 却抑制了同化物质的转运[64]。灌浆后期ABA 对籽粒灌浆的抑制作用机制目前尚不清楚。
ETH 对籽粒灌浆具有抑制作用。刘凯等[65]报道,喷施硝酸钴(一种ETH 合成抑制剂)降低了籽粒ACC 含量和EER,提高了灌浆速率和粒质量,而喷施乙烯利(ETH 释放的促进剂)的效果则相反。这与籽粒中SuSase、AGPase 以及可溶性淀粉合酶(SSS)的活性增强或降低密切相关[42,57,65]。据报道,ETH 也可作为信号分子诱导α-淀粉酶的表达,降低籽粒中淀粉的积累[66]。此外,ABA 和ACC 之间的相互作用也介导了水稻的花后发育,ABA/ACC 值与灌浆速率和籽粒充实率呈显著正相关,抑制ETH 水平或提高ABA/ACC 比值均有利于淀粉合成[67-68]。
3.4 多胺(PAs)和油菜素甾醇类(BRs)对籽粒灌浆和粒质量的调控作用及机理
据报道,籽粒游离Spd 和Spm 含量与灌浆速率呈显著或极显著正相关,而游离Put 含量与灌浆速率无显著相关性甚至呈负相关[45,69]。研究显示,水稻弱势粒中较低的游离Spd 和Spm 含量、较高的游离Put 含量是其灌浆速率慢、粒质量低的重要原因[69],较高的游离Spd 和Spm 含量、Spd/Put 或Spm/Put 比值有利于籽粒灌浆和粒质量的提高[70]。PAs 可通过调节SuSase、AGPase 和SSS 活性来影响籽粒灌浆[43,45,69]。
BRs 对水稻籽粒灌浆和粒质量具有正向调节作用。WU 等[71]通过构建转基因植株,发现BRs 加强了同化物从“源”向“库”的转运,促进了籽粒灌浆。王吉生等[72]在孕穗期对稻穗喷施2, 4-表油菜素内酯(BRs 家族的一种),结果显示,籽粒中SuSase 活性明显提高,蔗糖分解加速,进而促进了淀粉的合成与积累。
4 植物激素对稻米品质的调控作用及机理
4.1 植物激素对稻米加工与外观品质的调控作用及机理
研究表明,籽粒ETH 与游离Put 对稻米加工与外观品质的负面作用明显。EER 和游离Put 含量与稻米糙米率和精米率呈显著或极显著负相关。喷施外源ACC 使籽粒ETH 水平提高,对稻米加工与外观品质产生不良影响[73]。灌浆期籽粒中IAA、Z、ZR 和ABA 对稻米加工与外观品质也存在显著的调控作用,但其作用效应比较复杂,受品种、种植方式、籽粒粒位、外源化学物质施用浓度及施用时期等影响较大,且内在机理尚不清楚,有待深入研究[74-75]。
4.2 植物激素对稻米蒸煮与食味品质的调控作用及机理
据报道,灌浆中后期根系中Z 和ZR 含量与稻米胶稠度(GC)和碱消值呈显著或极显著正相关,与直链淀粉含量(AC)呈显著或极显著负相关[76]。灌浆中期根系ABA 浓度与GC 和碱消值呈极显著负相关,与AC 呈极显著正相关。外源喷施上述激素也有类似效应[73,76]。研究显示,灌浆中后期淀粉分支酶和异淀粉酶在一些米胶长、AC 低的品种中或外源喷施Z 和ZR 后活性较高,而在米胶短、AC 高的品种或喷施ABA 后活性较低[76]。除ABA 外,外源GAs 也有使GC 下降、AC 升高的趋势,不利于支链淀粉的积累,这可能与淀粉分支酶活性下降有关[74]。
此外,不同旱种方式下不同灌浆时期IAA、GA1、GA4、Z、ZR 和ABA 等含量与GC、AC 相关性不显著[78],这与常二华等[76]研究结果不同,可能是由于种植方式或品种的差异所致。不同旱种方式下灌浆各时期EER 与碱消值和崩解值呈显著或极显著负相关[77],说明ETH 对稻米蒸煮与食味品质存在一定负面影响[75]。
4.3 植物激素对稻米营养与卫生品质的调控作用及机理
稻米营养品质主要与稻米蛋白质或氨基酸含量有关[78]。研究表明,外源ABA 和GAs 有使籽粒粗蛋白含量下降的作用,其中外源ABA 的作用不显著[74]。XU 等[79]报道,EER 和ACC 含量与精米中必需氨基酸(EAAs)、非必需氨基酸(NEAAs)和总氨基酸(TAAs)含量呈极显著负相关,籽粒中较高的ETH 水平不利于氨基酸的积累。籽粒游离Spd 和Spm 等多胺类对稻米营养品质也具有重要影响。据报道,游离Spd 和Spm 含量与精米EAAs、NEAAs 和TAAs 含量呈极显著正相关,这得益于Spd 和Spm 通过增强籽粒中谷氨酸合成酶(GOGAT)、天冬氨酸转氨酶(AST)和丙氨酸转氨酶(ALT)等氨基酸合成代谢关键酶活性,进而促进籽粒氨基酸合成与积累[79]。
在稻米卫生品质上,潘九月[80]通过水培实验发现,施用外源SA 可使籽粒中镉(Cd)含量显著下降,这是因为SA 促进了细胞壁果胶和木质素的合成以及果胶的去甲酯化,增大了游离羧基的含量以及细胞壁的厚度,从而增强了对Cd 的吸附和阻挡作用,有效阻止Cd离子进入细胞壁。张盛楠等[81]报道,外源喷施JA 后,水稻品种中嘉早17 地上部和根部Cd 含量均下降,这与JA 降低Cd 离子的含量有关。
5 研究展望
近140 余年来,人们对植物激素的研究不断深入,其在水稻籽粒灌浆、粒质量和品质形成等方面取得的重要进展为稻米优质高产的遗传改良和栽培调控奠定了坚实基础。但是,目前关于植物激素对水稻籽粒灌浆、粒质量与品质的研究仍存在着一些问题。作者提出以下几点存在问题及今后研究的建议:
1)不同植物激素的相互作用对水稻品质形成的作用及机制缺乏研究。以往研究中,关于不同植物激素相互作用调控水稻灌浆和粒质量的研究较多,而其对稻米品质形成的作用及机制缺乏报道。建议今后进一步研究不同植物激素(包括经典植物激素和新型植物激素)对稻米品质形成的互作机制。
2)新型植物激素对水稻籽粒灌浆、粒质量和品质的影响及其机理研究较少。已有的植物激素对水稻籽粒灌浆、粒质量和品质形成的研究大多以5 类经典植物激素为中心,对PAs、BRs 和SA 等新型植物激素虽然有些研究,但不够深入。建议今后研究水稻籽粒中BRs、JA、SA 和SL 的含量变化特点及其对籽粒灌浆、粒质量和品质的重要调控作用。
3)植物激素对水稻籽粒灌浆、粒质量和品质的调控机理研究不深入。关于植物激素对籽粒灌浆、粒质量和品质的调控机理大多局限在蔗糖-淀粉代谢途径关键酶活性的调控上,缺乏深层次的研究。建议今后利用基因组学、转录组学、蛋白质组学、脂质组学以及代谢组学等技术,从籽粒碳、氮代谢途径和信号转导通路等多方位、多角度深入解析植物激素对水稻籽粒灌浆、粒质量和品质形成的生理与分子机制。
